
PPO2 Mutations in Amaranthus palmeri: Implications on Cross-Resistance

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fomesafen Dose–Response Assay
2.2. Response to Other Foliar-Applied PPO Herbicides
2.3. Detection of Mutations by TaqMan Genotyping Assay
3. Results
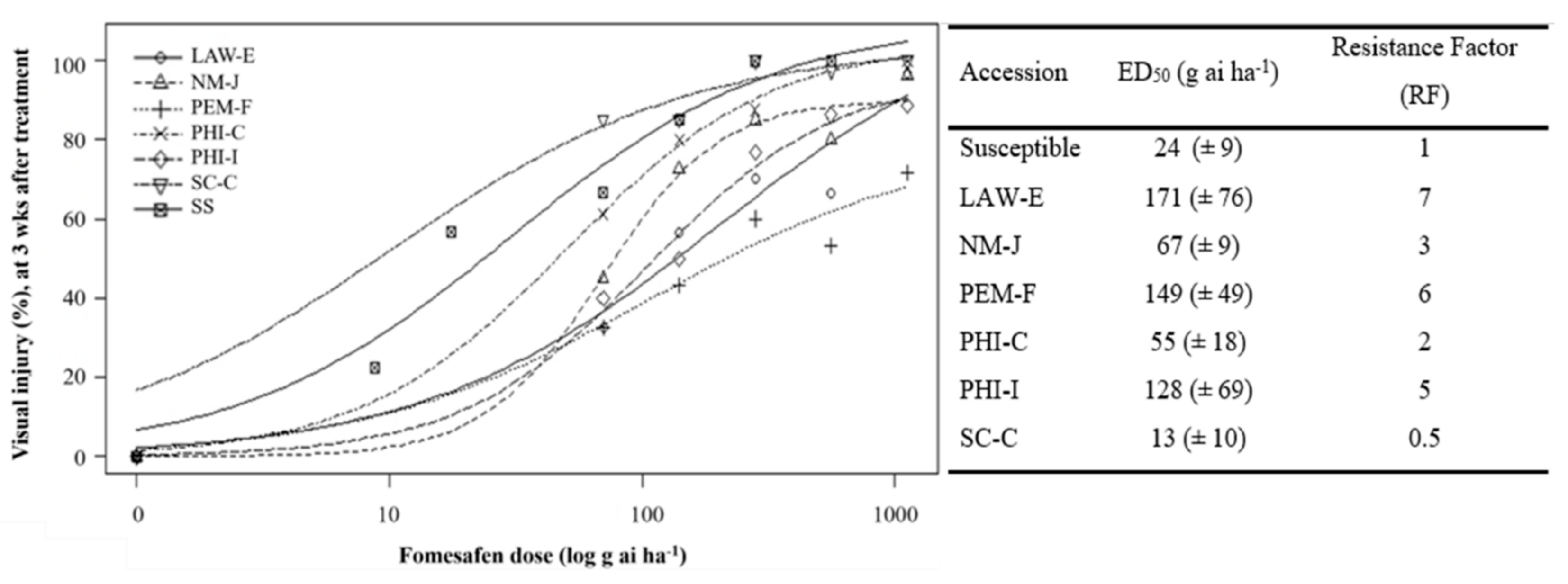
3.1. Fomesafen Dose–Response Assay
3.2. Response to Other Foliar-Applied Herbicides
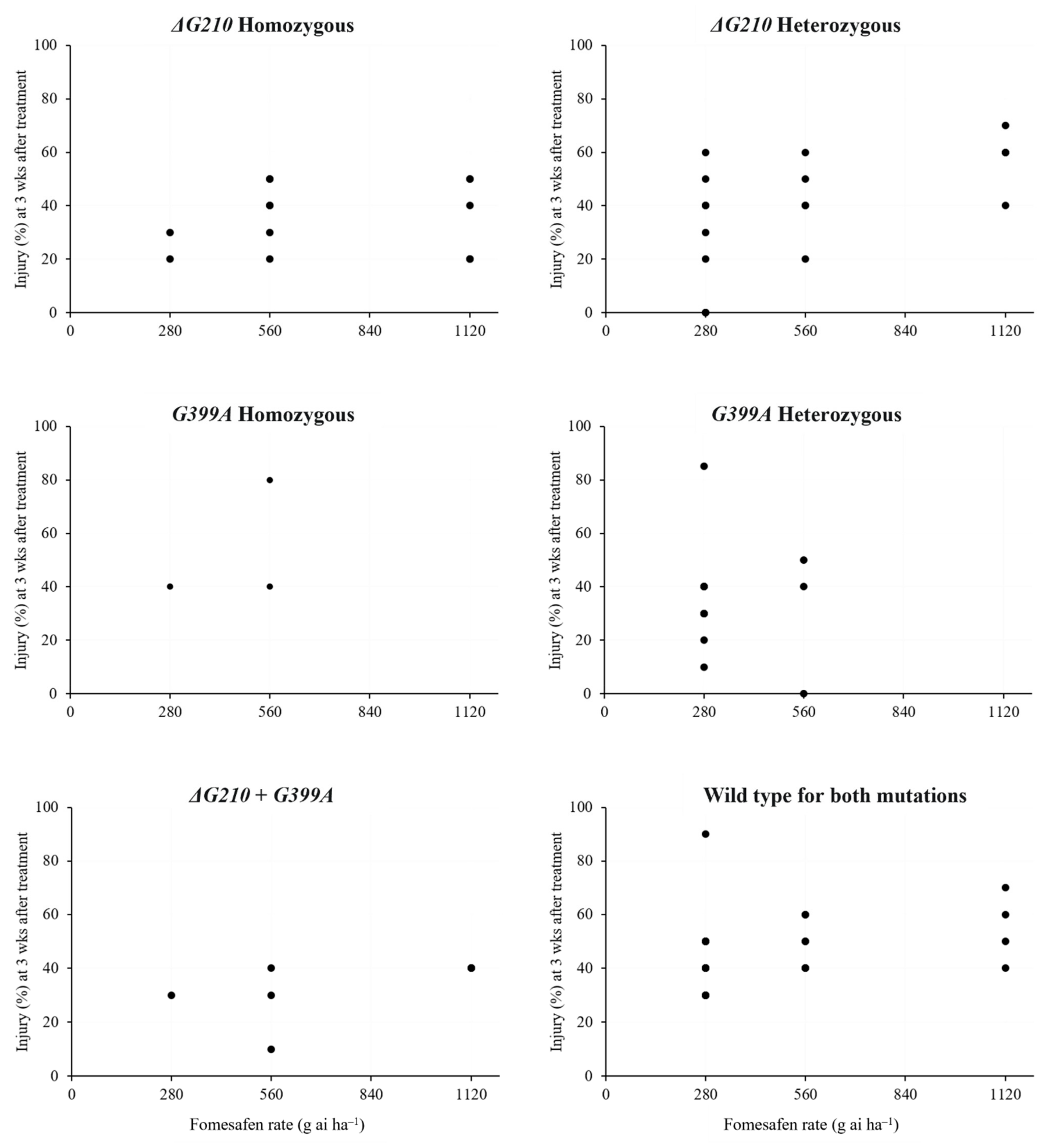
3.3. Detection of Mutations by TaqMan Genotyping Assay
4. Discussion
4.1. Resistance Level to Fomesafen and Overall Response to Other Foliar-Applied Herbicides
4.2. ppo2 Mutations Patterns among Survivors: Implications on Cross-Resistance to PPO Herbicides
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonny, S. Genetically modified herbicide-tolerant crops, weeds, and herbicides: Overview and impact. Environ. Manag. 2016, 57, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O. Taking stock of herbicide-resistant crops ten years after introduction. Pestic. Sci. 2005, 61, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Vencill, W.K.; Nichols, R.L.; Webster, T.M.; Soteres, J.K.; Mallory-Smith, C.; Burgos, N.R.; Johnson, W.G.; McClelland, M.R. Herbicide resistance: Toward an understanding of resistance development and the impact of herbicide-resistant crops. Weed Sci. 2012, 60, 2–30. [Google Scholar] [CrossRef] [Green Version]
- Christoffers, M.J. Genetic aspects of herbicide-resistant weed management. Weed Technol. 1999, 13, 647–652. [Google Scholar] [CrossRef]
- Gaines, T.A.; Duke, S.O.; Morran, S.; Rigon, C.A.; Tranel, P.J.; Küpper, A.; Dayan, F.E. Mechanisms of evolved herbicide resistance. J. Biol. Chem. 2020, 295, 10307–10330. [Google Scholar] [CrossRef] [PubMed]
- Heap, I. The International Herbicide-Resistant Weed Database. Available online: http://www.weedscience.org/ (accessed on 12 February 2021).
- Franssen, A.S.; Skinner, D.Z.; Al-Khatib, K.; Horak, M.J.; Kulakow, P.A. Interspecific hybridization and gene flow of ALS resistance in Amaranthus species. Weed Sci. 2001, 49, 598–606. [Google Scholar] [CrossRef]
- Molin, W.T.; Nandula, V.K.; Wright, A.A.; Bond, J.A. Transfer and expression of ALS inhibitor resistance from Palmer amaranth (Amaranthus palmeri) to an A. spinosus × A. palmeri hybrid. Weed Sci. 2016, 64, 240–247. [Google Scholar] [CrossRef]
- Steckel, L.E. The dioecious Amaranthus spp.: Here to stay. Weed Technol. 2007, 21, 567–570. [Google Scholar] [CrossRef]
- Chaudhari, S.; Varanasi, V.K.; Nakka, S.; Bhowmik, P.C.; Thompson, C.R.; Peterson, D.E.; Currie, R.S.; Jugulam, M. Evolution of target and non-target based multiple herbicide resistance in a single Palmer amaranth (Amaranthus palmeri) population from Kansas. Weed Technol. 2020, 34, 447–453. [Google Scholar] [CrossRef]
- Salas-Perez, R.A.; Burgos, N.R.; Rangani, G.; Singh, S.; Refatti, J.P.; Piveta, L.; Tranel, P.J.; Mauromoustakos, A.; Scott, R.C. Frequency of Gly-210 deletion mutation among protoporphyrinogen oxidase inhibitor–resistant Palmer amaranth (Amaranthus palmeri) populations. Weed Sci. 2017, 65, 718–731. [Google Scholar] [CrossRef]
- Spaunhorst, D.J.; Nie, H.; Todd, J.R.; Young, J.M.; Young, B.G.; Johnson, W.G. Confirmation of herbicide resistance mutations Trp574Leu, ΔG210, and EPSPS gene amplification and control of multiple herbicide-resistant Palmer amaranth (Amaranthus palmeri) with chlorimuron-ethyl, fomesafen, and glyphosate. PLoS ONE 2019, 14, e0214458. [Google Scholar] [CrossRef]
- Jacobs, J.M.; Jacobs, N.J.; Sherman, T.D.; Duke, S.O. Effect of diphenyl ether herbicides on oxidation of protoporphyrinogen to protoporphyrin in organellar and plasma membrane enriched fractions of barley. Plant Physiol. 1991, 97, 197–203. [Google Scholar] [CrossRef]
- Lee, H.J.; Duke, M.V.; Duke, S.O. Cellular localization of protoporphyrinogen-oxidizing activities of etiolated barley (Hordeum vulgare L.) leaves (relationship to mechanism of action of protoporphyrinogen oxidase-inhibiting herbicides). Plant Physiol. 1993, 102, 881–889. [Google Scholar] [CrossRef] [Green Version]
- Matringe, M.; Camadro, J.M.; Labbe, P.; Scalla, R. Protoporphyrinogen oxidase as a molecular target for diphenyl ether herbicides. Biochem. J. 1989, 260, 231–235. [Google Scholar] [CrossRef]
- Orr, G.L.; Hess, F.D. Mechanism of action of the diphenyl ether herbicide acifluorfen-methyl in excised cucumber (Cucumis sativus L.) cotyledons: Light activation and the subsequent formation of lipophilic free radicals. Plant Physiol. 1982, 69, 502–507. [Google Scholar] [CrossRef] [Green Version]
- Poulson, R.; Polglase, W.J. The enzymic conversion of protoporphyrinogen IX to protoporphyrin IX. Protoporphyrinogen oxidase activity in mitochondrial extracts of Saccharomyces cerevisiae. J. Biol. Chem. 1975, 250, 1269–1274. [Google Scholar] [CrossRef]
- Lermontova, I.; Kruse, E.; Mock, H.P.; Grimm, B. Cloning and characterization of a plastidal and a mitochondrial isoform of tobacco protoporphyrinogen IX oxidase. Proc. Natl. Acad. Sci. USA 1997, 94, 8895–8900. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, N.; Che, F.S.; Iwano, M.; Takayama, S.; Yoshida, S.; Isogai, A. Dual targeting of spinach protoporphyrinogen oxidase II to mitochondria and chloroplasts by alternative use of two in-frame initiation codons. J. Biol. Chem. 2001, 276, 20474–20481. [Google Scholar] [CrossRef] [Green Version]
- Copeland, J.D.; Giacomini, D.A.; Tranel, P.J.; Montgomery, G.B.; Steckel, L.E. Distribution of PPX2 mutations conferring PPO-inhibitor resistance in Palmer amaranth populations of Tennessee. Weed Technol. 2018, 32, 592–596. [Google Scholar] [CrossRef]
- Patzoldt, W.L.; Hager, A.G.; McCormick, J.S.; Tranel, P.J. A codon deletion confers resistance to herbicides inhibiting protoporphyrinogen oxidase. Proc. Natl. Acad. Sci. USA 2006, 103, 12329–12334. [Google Scholar] [CrossRef] [Green Version]
- Salas, R.A.; Burgos, N.R.; Tranel, P.J.; Singh, S.; Glasgow, L.; Scott, R.C.; Nichols, R.L. Resistance to PPO-inhibiting herbicide in Palmer amaranth from Arkansas. Pest Manag. Sci. 2016, 72, 864–869. [Google Scholar] [CrossRef] [Green Version]
- Giacomini, D.A.; Umphres, A.M.; Nie, H.; Mueller, T.C.; Steckel, L.E.; Young, B.G.; Scott, R.C.; Tranel, P.J. Two new PPX2 mutations associated with resistance to PPO-inhibiting herbicides in Amaranthus palmeri. Pest Manag. Sci. 2017, 73, 1559–1563. [Google Scholar] [CrossRef]
- Varanasi, V.K.; Brabham, C.; Norsworthy, J.K.; Nie, H.; Young, B.G.; Houston, M.; Barber, T.; Scott, R.C. A statewide survey of PPO-inhibitor resistance and the prevalent target-site mechanisms in Palmer amaranth (Amaranthus palmeri) accessions from Arkansas. Weed Sci. 2018, 66, 149–158. [Google Scholar] [CrossRef]
- Rousonelos, S.L.; Lee, R.M.; Moreira, M.S.; VanGessel, M.J.; Tranel, P.J. Characterization of a common ragweed (Ambrosia artemisiifolia) population resistant to ALS- and PPO-inhibiting herbicides. Weed Sci. 2012, 60, 335–344. [Google Scholar] [CrossRef]
- Rangani, G.; Salas-Perez, R.A.; Aponte, R.A.; Knapp, M.; Craig, I.R.; Mietzner, T.; Langaro, A.C.; Noguera, M.M.; Porri, A.; Roma-Burgos, N. A novel single-site mutation in the catalytic domain of protoporphyrinogen oxidase IX (PPO) confers resistance to PPO-inhibiting herbicides. Front. Plant Sci. 2019, 10, 568. [Google Scholar] [CrossRef]
- Wu, C.; Goldsmith, M.R.; Pawlak, J.; Feng, P.; Smith, S.; Navarro, S.; Perez-Jones, A. Differences in efficacy, resistance mechanism and target protein interaction between two PPO inhibitors in Palmer amaranth (Amaranthus palmeri). Weed Sci. 2020, 68, 105–115. [Google Scholar] [CrossRef]
- Noguera, M.M.; Rangani, G.; Heiser, J.; Bararpour, T.; Steckel, L.E.; Betz, M.; Porri, A.; Lerchl, J.; Zimmermann, S.; Nichols, R.L.; et al. Functional PPO2 mutations: Co- occurrence in one plant or the same ppo2 allele of herbicide-resistant Amaranthus palmeri in the US Mid-south. Pest Manag. Sci. 2020, 77, 1001–1012. [Google Scholar] [CrossRef]
- Burgos, N.R.; Tranel, P.J.; Streibig, J.C.; Davis, V.M.; Shaner, D.; Norsworthy, J.K.; Ritz, C. Confirmation of resistance to herbicides and evaluation of resistance levels. Weed Sci. 2013, 61, 4–20. [Google Scholar] [CrossRef]
- Frans, R.; Talbert, R.; Marx, D.; Crowley, H. Experimental design and techniques for measuring and analyzing plant responses to weed control practices. In Southern Weed Science Society, Research Methods in Weed Science; Camper, N.D., Ed.; Weed Science Society of America: Champaign, IL, USA, 1986; pp. 29–46. [Google Scholar]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef] [Green Version]
- Ritz, C. Toward a unified approach to dose–response modeling in ecotoxicology. Environ. Toxicol. Chem. 2010, 29, 220–229. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Gbur, E.E.; Stroup, W.W.; McCarter, K.S.; Durham, S.; Young, L.J.; Christman, M.; West, M.; Kramer, M. Analysis of Generalized Linear Mixed Models in the Agricultural and Natural Resources Sciences; American Society of Agronomy, Soil Science Society of America, Crop Science Society of America: Madison, WI, USA, 2012; pp. 7–58. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Wuerffel, R.J.; Young, J.M.; Lee, R.M.; Tranel, P.J.; Lightfoot, D.A.; Young, B.G. Distribution of the ΔG210 protoporphyrinogen oxidase mutation in Illinois waterhemp (Amaranthus tuberculatus) and an improved molecular method for detection. Weed Sci. 2015, 63, 839–845. [Google Scholar] [CrossRef]
- Varanasi, V.K.; Brabham, C.; Korres, N.E.; Norsworthy, J.K. Nontarget site resistance in Palmer amaranth [Amaranthus palmeri (S.) Wats.] confers cross-resistance to protoporphyrinogen oxidase-inhibiting herbicides. Weed Technol. 2019, 33, 349–354. [Google Scholar] [CrossRef]
- Lillie, K.J.; Giacomini, D.A.; Tranel, P.J. Comparing responses of sensitive and resistant populations of Palmer amaranth (Amaranthus palmeri) and waterhemp (Amaranthus tuberculatus var. rudis) to PPO inhibitors. Weed Technol. 2019, 34, 140–146. [Google Scholar] [CrossRef]
- Shoup, D.E.; Al-Khatib, K.; Peterson, D.E. Common waterhemp (Amaranthus rudis) resistance to protoporphyrinogen oxidase-inhibiting herbicides. Weed Sci. 2003, 51, 145–150. [Google Scholar] [CrossRef]
- Huang, Z.; Cui, H.; Wang, C.; Wu, T.; Zhang, C.; Huang, H.; Wei, S. Investigation of resistance mechanism to fomesafen in Amaranthus retroflexus L. Pestic. Biochem. Physiol. 2020, 165, 104560. [Google Scholar] [CrossRef]
- Sharpen Herbicide Product Label. Research Drive; BASF Corporation: Florham Park, NJ, USA, 2020; Available online: http://www.cdms.net/ldat/ld99E001.pdf (accessed on 12 March 2021).
- Montgomery, G.B.; Bond, J.A.; Golden, B.R.; Gore, J.; Edwards, H.M.; Eubank, T.W.; Walker, T.W. Evaluation of saflufenacil in drill-seeded rice (Oryza sativa). Weed Technol. 2014, 28, 660–670. [Google Scholar] [CrossRef]
- Morichetti, S.; Ferrell, J.; MacDonald, G.; Sellers, B.; Rowland, D. Weed management and peanut response from applications of saflufenacil. Weed Technol. 2012, 26, 261–266. [Google Scholar] [CrossRef]
- Houston, M.M.; Norsworthy, J.K.; Barber, T.; Brabham, C. Field evaluation of preemergence and postemergence herbicides for control of protoporphyrinogen oxidase-resistant Palmer amaranth (Amaranthus palmeri S. Watson). Weed Technol. 2019, 33, 610–615. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.M.; Strom, S.A.; Riechers, D.E.; Davis, A.S.; Tranel, P.J.; Hager, A.G. Characterization of a waterhemp (Amaranthus tuberculatus) population from Illinois resistant to herbicides from five site-of-action groups. Weed Technol. 2019, 33, 400–410. [Google Scholar] [CrossRef]
- BASF Corporation. Voraxor Herbicide Product Label. Available online: https://agro.basf.ca/basf/agprocan/agsolutions/solutions.nsf/Images/PDC-CREO-BZ8N4V/$File/Voraxor_Product_Label.pdf (accessed on 12 March 2021).
- Bi, B.; Wang, Q.; Coleman, J.J.; Porri, A.; Peppers, J.M.; Patel, J.D.; Betz, M.; Lerchl, J.; McElroy, J.S. A novel mutation A212T in chloroplast protoporphyrinogen oxidase (PPO1) confers resistance to PPO inhibitor oxadiazon in Eleusine indica. Pest Manag. Sci. 2020, 76, 1786–1794. [Google Scholar] [CrossRef]
- Wang, D.W.; Zhang, R.B.; Yu, S.Y.; Liang, L.; Ismail, I.; Li, Y.H.; Xu, H.; Wen, X.; Xi, Z. Discovery of novel N-isoxazolinylphenyltriazinones as promising protoporphyrinogen IX oxidase inhibitors. J. Agric. Food Chem. 2019, 67, 12382–12392. [Google Scholar] [CrossRef]
- Armel, G.R.; Hanzlik, K.; Witschel, M.; Hennigh, D.S.; Bowe, S.; Simon, A.; Liebl, R.; Mankin, L. Trifludimoxazin: A new PPO inhibitor that controls PPO resistant weed biotypes. In Proceedings of the Weed Science Society of America, Tucson, AZ, USA, 6–9 February 2017; p. 218. [Google Scholar]
- Montgomery, J.S.; Giacomini, D.A.; Tranel, P.J. Molecular confirmation of resistance to PPO inhibitors in Amaranthus tuberculatus and Amaranthus palmeri, and isolation of the G399A PPO2 substitution in A. palmeri. Weed Technol. 2020, 35, 1–7. [Google Scholar] [CrossRef]
- Yu, Q.; Jalaludin, A.; Han, H.; Chen, M.; Sammons, R.D.; Powles, S.B. Evolution of a double amino acid substitution in the 5-enolpyruvylshikimate-3-phosphate synthase in Eleusine indica conferring high-level glyphosate resistance. Plant Physiol. 2015, 167, 1440–1447. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Vila-Aiub, M.M.; Jalaludin, A.; Yu, Q.; Powles, S.B. A double EPSPS gene mutation endowing glyphosate resistance shows a remarkably high resistance cost. Plant Cell Environ. 2017, 40, 3031–3042. [Google Scholar] [CrossRef]
- Singh, S.; Singh, V.; Salas-Perez, R.A.; Bagavathiannan, M.V.; Lawton-Rauh, A.; Roma-Burgos, N. Target-site mutation accumulation among ALS inhibitor-resistant Palmer amaranth. Pest Manag. Sci. 2019, 75, 1131–1139. [Google Scholar] [CrossRef]
- Brabham, C.; Varanasi, V.; Norsworthy, J.K. The level of PPO-inhibitor resistance conferred by different mutations in Palmer amaranth. In Proceedings of the 71st Annual Meeting of the Southern Weed Science Society, Atlanta, GE, USA, 22–24 January 2018; p. 18. [Google Scholar]
- Obenland, O.A.; Ma, R.; O’Brien, S.R.; Lygin, A.V.; Riechers, D.E. Carfentrazone-ethyl resistance in an Amaranthus tuberculatus population is not mediated by amino acid alterations in the PPO2 protein. PLoS ONE 2019, 14, e0215431. [Google Scholar] [CrossRef] [Green Version]
- Varanasi, V.K.; Brabham, C.; Norsworthy, J.K. Confirmation and characterization of non—Target site resistance to fomesafen in Palmer amaranth (Amaranthus palmeri). Weed Sci. 2018, 66, 702–709. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Accession | Origin State | ΔG210 | G399A |
|---|---|---|---|
| LAW-E a | Arkansas | Present | Absent |
| NM-J | Missouri | Present | Present |
| PEM-F | Missouri | Present | Present |
| PHI-C | Arkansas | Present | Absent |
| PHI-I | Arkansas | Absent | Present |
| SC-C | Missouri | Absent | Present |
| SS b | Arkansas | Absent | Absent |
| Common Name a | Trade Name | Chemical Family | Field Rate |
|---|---|---|---|
| g ai ha−1 | |||
| Carfentrazone | Aim® 2EC | Aryl triazinone | 280 |
| Flumioxazin | Valor® SX 51WDG | N-phenylphthalimide | 71.5 |
| Fomesafen | Flexstar® 1.88SL | Diphenyl ether | 280 |
| Saflufenacil | Sharpen® 4F | Pyrimidinedione | 25 |
| Trifludimoxazin | Tirexor™ b | Triazinone | 30 |
| Factors Evaluated | ANOVA p Values | |||
|---|---|---|---|---|
| Carfentrazone | Flumioxazin | Saflufenacil | Trifludimoxazin | |
| Accession | <0.0001 | <0.0001 | <0.0001 | 0.0277 |
| Dose | 0.0002 | 0.1332 | 0.0201 | 0.0556 |
| Acc × Dose | 0.0389 | 0.5124 | 0.4518 | 0.8619 |
| Accession | Dose a | Mortality | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Carfentrazone | Flumioxazin | Saflufenacil | Trifludimoxazin | ||||||
| % | |||||||||
| LAW-E | 1× | 27 | * | 16 | * | 54 | * | 87 | * |
| 2× | 31 | * | 10 | 78 | 92 | ||||
| NM-J | 1× | 41 | * | 18 | * | 86 | ns | 90 | ns |
| 2× | 93 | ns | 42 | 93 | 94 | ||||
| PEM-F | 1× | 30 | * | 10 | * | 75 | * | 87 | * |
| 2× | 42 | * | 11 | 83 | 94 | ||||
| PHI-C | 1× | 66 | * | 95 | ns | 93 | ns | 100 | ns |
| 2× | 83 | * | 99 | 100 | 100 | ||||
| PHI-I | 1× | 37 | * | 20 | * | 93 | ns | 94 | ns |
| 2× | 58 | * | 47 | 92 | 100 | ||||
| SC-C | 1× | 55 | * | 87 | ns | 89 | ns | 100 | ns |
| 2× | 93 | ns | 96 | 93 | 100 | ||||
| SS b | 1× | 100 | 100 | 100 | 100 | ||||
| 2× | 100 | 100 | 100 | 100 | |||||
| Accession | No. of Plants Genotyped | ΔG210 Only | G399A Only | ΔG210 + G399A | WT c | WT Injury (%) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| RR a | Rr b | RR | Rr | Min | Max | Average | ||||
| LAW-E d | 16 | 11 | 4 | - | - | - | 1 | 50 | 50 | 50 |
| NM-J d | 13 | - | 2 | 2 | 4 | - | 5 | 40 | 70 | 45 |
| PEM-F d | 22 | 2 | 6 | - | 6 | 7 | 1 | 50 | 50 | 50 |
| PHI-C e | 4 | - | 2 | - | - | - | 2 | 30 | 30 | 30 |
| PHI-I d | 16 | - | - | - | 1 | - | 15 | 30 | 60 | 46 |
| SC-C f | 4 | - | 1 | 1 | 1 | - | 1 | 50 | 50 | 50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carvalho-Moore, P.; Rangani, G.; Heiser, J.; Findley, D.; Bowe, S.J.; Roma-Burgos, N. PPO2 Mutations in Amaranthus palmeri: Implications on Cross-Resistance. Agriculture 2021, 11, 760. https://doi.org/10.3390/agriculture11080760
Carvalho-Moore P, Rangani G, Heiser J, Findley D, Bowe SJ, Roma-Burgos N. PPO2 Mutations in Amaranthus palmeri: Implications on Cross-Resistance. Agriculture. 2021; 11(8):760. https://doi.org/10.3390/agriculture11080760
Chicago/Turabian StyleCarvalho-Moore, Pâmela, Gulab Rangani, James Heiser, Douglas Findley, Steven J. Bowe, and Nilda Roma-Burgos. 2021. "PPO2 Mutations in Amaranthus palmeri: Implications on Cross-Resistance" Agriculture 11, no. 8: 760. https://doi.org/10.3390/agriculture11080760