
Effects of Short-Term Rice Straw Return on the Soil Microbial Community

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. DNA Extraction and High-Throughput Sequencing
2.4. Data Analysis
3. Results
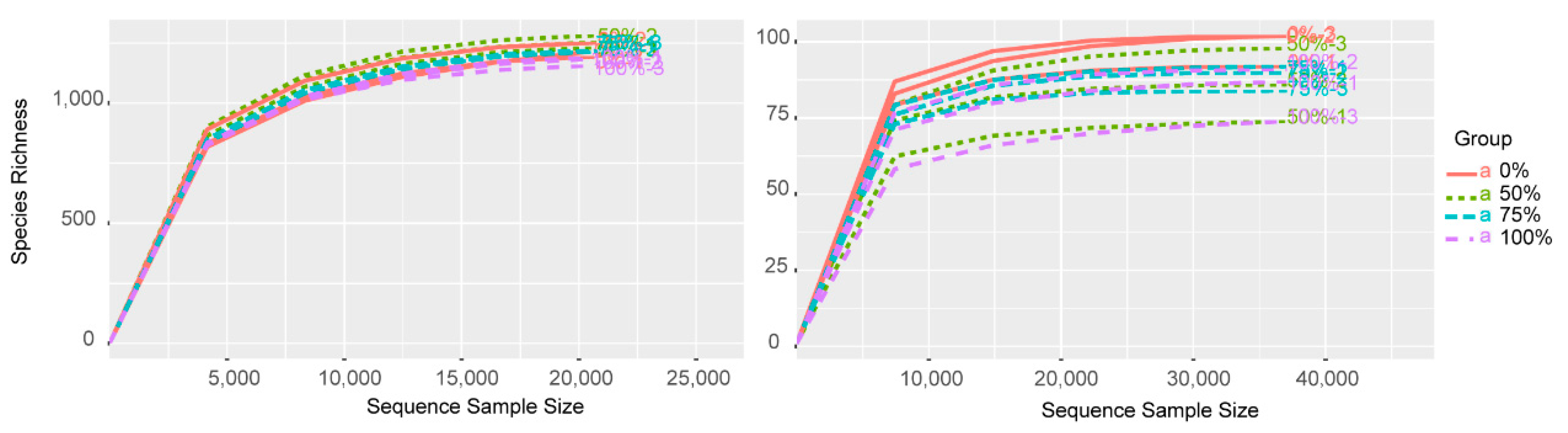
3.1. Alpha Diversity of the Microbial Communities
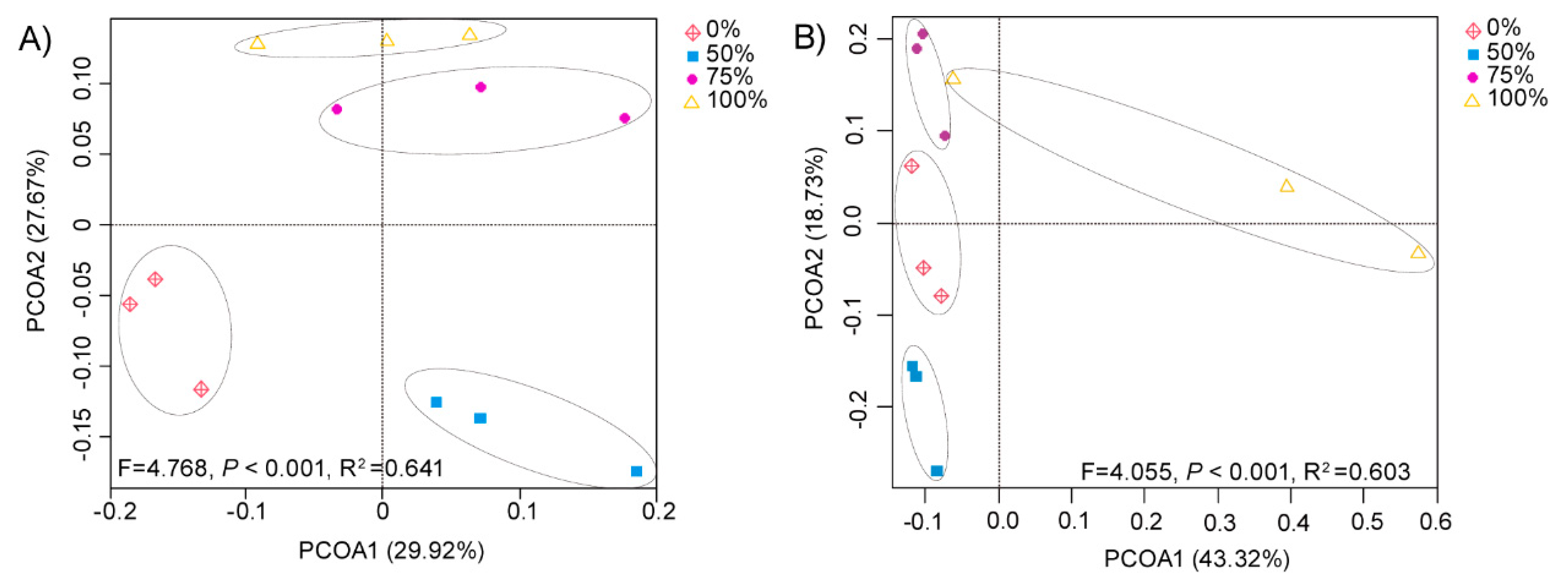
3.2. Effect of Rice Straw Returning on Bacterial and Fungal Beta Diversity
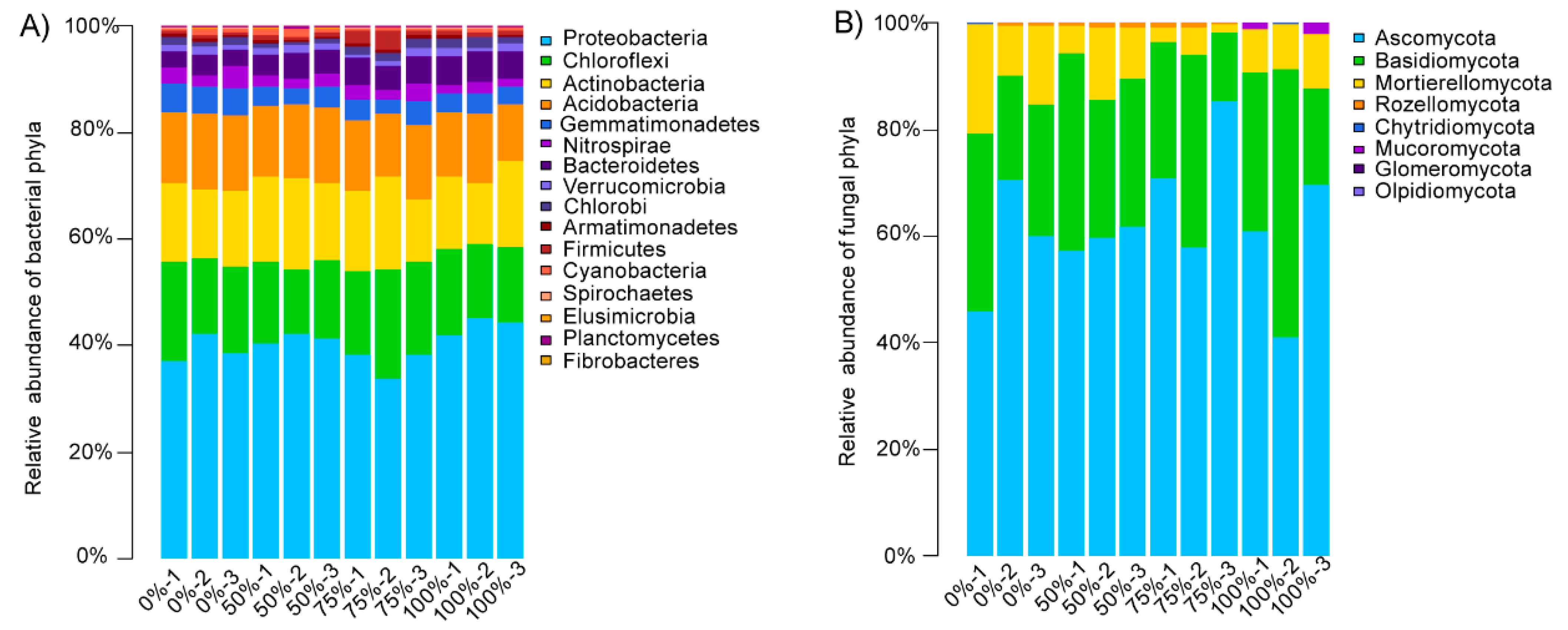
3.3. Compositional Characteristics of Soil Microbial Communities under Different Straw Addition Levels
3.3.1. Compositional Characteristics of Soil Bacterial Communities under Different Straw Addition Levels
3.3.2. Compositional Characteristics of Soil Fungal Communities under Different Straw Addition Levels
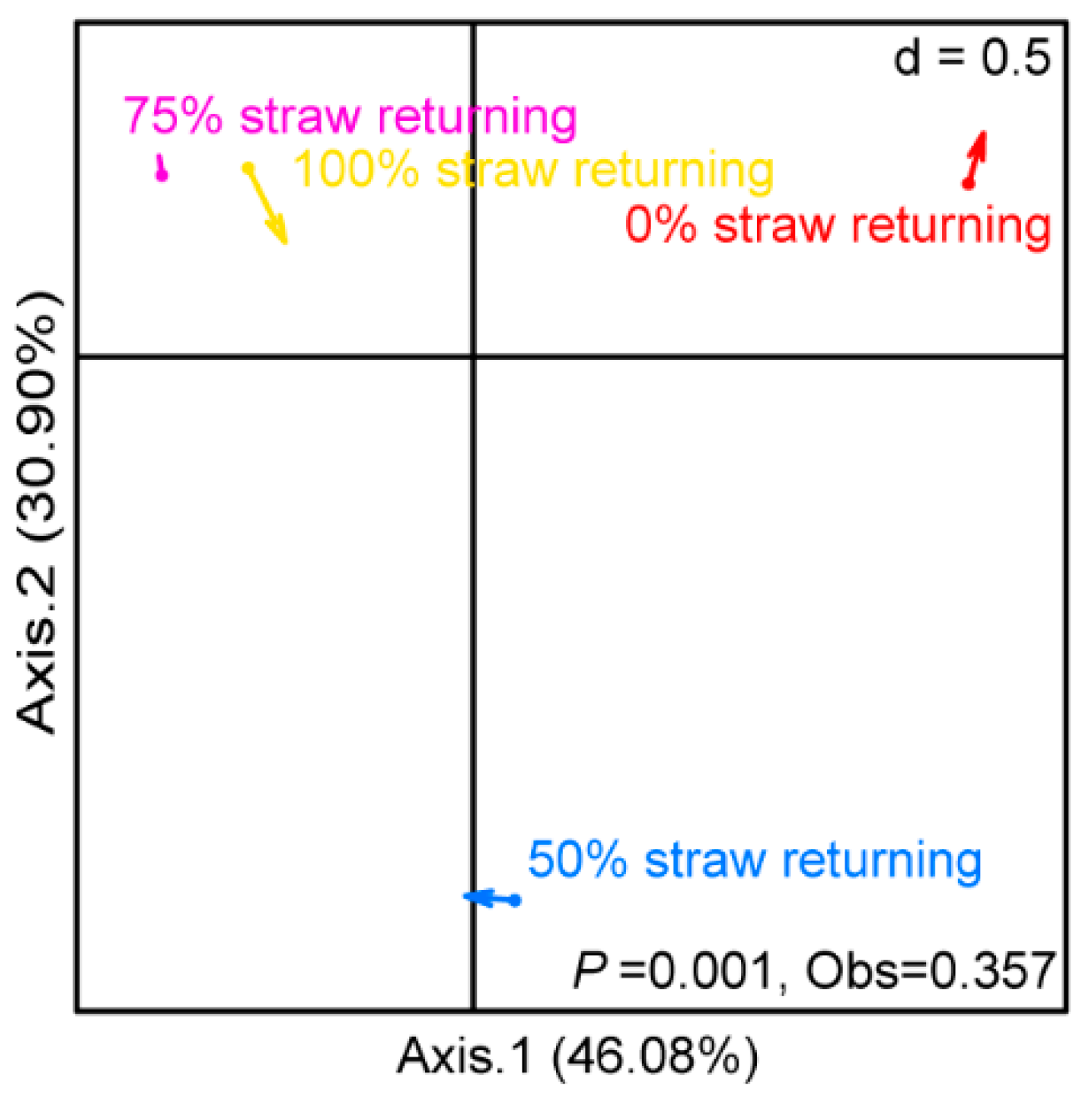
3.4. Correlation between Bacteria and Fungi under Different Straw Addition Levels
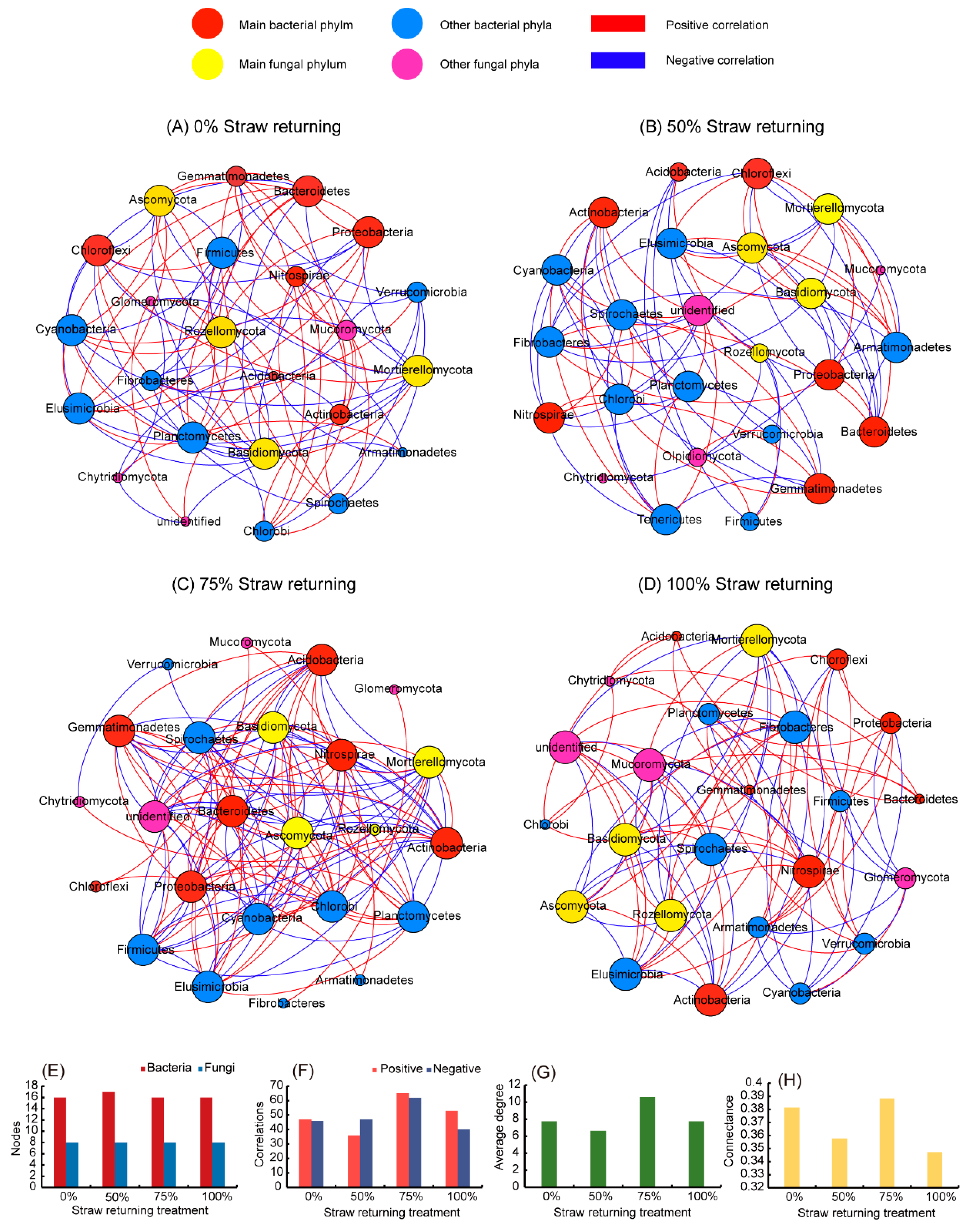
3.5. The Network Correlation between Bacteria and Fungi under Different Amounts of Straw Returning
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Network Parameter Characteristics | 0% Straw Returning | 50% Straw Returning | 75% Straw Returning | 100% Straw Returning |
|---|---|---|---|---|
| Bacteria notes | 16 | 17 | 16 | 16 |
| Fungi notes | 8 | 8 | 8 | 8 |
| Positive correlations | 47 | 36 | 65 | 53 |
| Negative correlations | 46 | 47 | 62 | 40 |
| Average degree | 7.75 | 6.64 | 10.583 | 7.75 |
| Density | 0.337 | 0.277 | 0.46 | 0.337 |
| Modularity | 0.548 | 0.609 | 0.106 | 0.548 |
| Connectance | 0.3815 | 0.3578 | 0.3885 | 0.3473 |
References
- Liu, X.B.; Zhang, X.Y.; Wang, Y.X.; Sui, Y.Y.; Zhang, S.L.; Herbert, S.J. Soil degradation: A problem threatening the sustainable development of agriculture in Northeast China. Plant Soil Environ. 2010, 56, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.J.; Sui, Y.Y.; Yu, Z.H.; Shi, Y.; Chu, H.Y.; Jin, J. Soil carbon content drives the biogeographical distribution of fungal communities in the black soil zone of northeast China. Soil Biol. Biochem. 2015, 83, 29–39. [Google Scholar] [CrossRef]
- von Luetzow, M.; Koegel-Knabner, I.; Ekschmitt, K.; Matzner, E.; Guggenberger, G. Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions—A review. Eur. J. Soil Sci. 2006, 57, 426–445. [Google Scholar] [CrossRef]
- Zhang, P.; Wei, T.; Jia, Z.K.; Han, Q.F.; Ren, X.L. Soil aggregate and crop yield changes with different rates of straw incorporation in semiarid areas of northwest China. Geoderma 2014, 230, 41–49. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Friedel, J.K.; Stahr, K. Review of mechanisms and quantification of priming effects. Soil Biol. Biochem. 2000, 32, 1485–1498. [Google Scholar] [CrossRef]
- Song, C.; Wang, E.L.; Han, X.Z.; Stirzaker, R. Crop production, soil carbon and nutrient balances as affected by fertilisation in a Mollisol agroecosystem. Nutr. Cycl. Agroecosys 2011, 89, 363–374. [Google Scholar] [CrossRef]
- Shahbaz, M.; Kumar, A.; Kuzyakov, Y.; Borjesson, G.; Blagodatskaya, E. Priming effects induced by glucose and decaying plant residues on SOM decomposition: A three-source C-13/C-14 partitioning study. Soil Biol. Biochem. 2018, 121, 138–146. [Google Scholar] [CrossRef]
- Li, H.; Cao, Y.; Wang, X.M.; Ge, X.; Li, B.Q.; Jin, C.Q. Evaluation on the Production of Food Crop Straw in China from 2006 to 2014. BioEnergy Res. 2017, 10, 949–957. [Google Scholar] [CrossRef]
- Yang, H.S.; Fang, C.; Meng, Y.; Dai, Y.J.; Liu, J. Long-term ditch-buried straw return increases functionality of soil microbial communities. Catena 2021, 202, 105316. [Google Scholar] [CrossRef]
- Shan, J.; Yan, X.Y. Effects of crop residue returning on nitrous oxide emissions in agricultural soils. Atmos. Environ. 2013, 71, 170–175. [Google Scholar] [CrossRef]
- Karami, A.; Homaee, M.; Afzalinia, S.; Ruhipour, H.; Basirat, S. Organic resource management: Impacts on soil aggregate stability and other soil physico-chemical properties. Agric. Ecosyst. Environ. 2012, 148, 22–28. [Google Scholar] [CrossRef]
- Blagodatskaya, E.V.; Blagodatsky, S.A.; Anderson, T.H.; Kuzyakov, Y. Priming effects in Chernozem induced by glucose and N in relation to microbial growth strategies. Appl. Soil Ecol. 2007, 37, 95–105. [Google Scholar] [CrossRef]
- Henriksen, T.M.; Breland, T.A. Carbon mineralization, fungal and bacterial growth, and enzyme activities as affected by contact between crop residues and soil. Biol. Fertil. Soils 2002, 35, 41–48. [Google Scholar] [CrossRef]
- Geisseler, D.; Horwath, W.R.; Scow, K.M. Soil moisture and plant residue addition interact in their effect on extracellular enzyme activity. Pedobiologia 2011, 54, 71–78. [Google Scholar] [CrossRef]
- Zhao, S.C.; Qiu, S.J.; Xu, X.P.; Ciampitti, I.A.; Zhang, S.Q.; He, P. Change in straw decomposition rate and soil microbial community composition after straw addition in different long-term fertilization soils. Appl. Soil Ecol. 2019, 138, 123–133. [Google Scholar] [CrossRef]
- Kamble, P.N.; Baath, E. Comparison of fungal and bacterial growth after alleviating induced N-limitation in soil. Soil Biol. Biochem. 2016, 103, 97–105. [Google Scholar] [CrossRef]
- Chibuike, G.; Burkitt, L.; Bretherton, M.; Singh, R.; Bishop, P. Dissolved Organic Carbon Concentration and Denitrification Capacity of a New Zealand Hill Country Soil After Forage Crop Establishment. Earth Syst. Environ. 2021, 1–14. [Google Scholar] [CrossRef]
- Zhao, D.S.; Zheng, D.; Wu, S.H.; Wu, Z.F. Climate changes in northeastern China during last four decades. Chinese Geogr. Sci. 2007, 17, 317–324. [Google Scholar] [CrossRef]
- Sun, W.M.; Xiao, E.Z.; Pu, Z.L.; Krumins, V.; Dong, Y.R.; Li, B.Q. Paddy soil microbial communities driven by environment- and microbe-microbe interactions: A case study of elevation-resolved microbial communities in a rice terrace. Sci. Total Environ. 2018, 612, 884–893. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Qin, H.L.; Chen, Z.; Wu, J.S.; Wei, W.X. Effect of long-term fertilization on bacterial composition in rice paddy soil. Biol. Fertil. Soils 2011, 47, 397–405. [Google Scholar] [CrossRef]
- Kikuchi, H.; Watanabe, T.; Jia, Z.; Kimura, M.; Asakawa, S. Molecular analyses reveal stability of bacterial communities in bulk soil of a Japanese paddy field: Estimation by denaturing gradient gel electrophoresis of 16S rRNA genes amplified from DNA accompanied with RNA. Soil Sci. Plant Nutr. 2007, 53, 448–458. [Google Scholar] [CrossRef]
- Xia, Y.H.; Chen, X.B.; Zheng, X.D.; Deng, S.H.; Hu, Y.J.; Zheng, S.M. Preferential uptake of hydrophilic and hy-drophobic compounds by bacteria and fungi in upland and paddy soils. Soil Biol. Biochem. 2020, 148, 107879. [Google Scholar] [CrossRef]
- Wei, L.; Ge, T.; Zhu, Z.; Luo, Y.; Yang, Y.; Xiao, M. Comparing carbon and nitrogen stocks in paddy and upland soils: Accumulation, stabilization mechanisms, and environmental drivers. Geoderma 2021, 398, 115121. [Google Scholar] [CrossRef]
- Li, Y.H.; Shahbaz, M.; Zhu, Z.K.; Deng, Y.W.; Tong, Y.Y.; Chen, L. Oxygen availability determines key regulators in soil organic carbon mineralisation in paddy soils. Soil Biol. Biochem. 2021, 153, 108106. [Google Scholar] [CrossRef]
- Chen, Z.; Luo, X.Q.; Hu, R.G.; Wu, M.N.; Wu, J.S.; Wei, W.X. Impact of Long-Term Fertilization on the Composition of Denitrifier Communities Based on Nitrite Reductase Analyses in a Paddy Soil. Microb. Ecol. 2010, 60, 850–861. [Google Scholar] [CrossRef]
- Yang, H.S.; Feng, J.X.; Zhai, S.L.; Dai, Y.J.; Xu, M.M.; Wu, J.S. Long-term ditch-buried straw return alters soil water potential, temperature, and microbial communities in a rice-wheat rotation system. Soil Tillage Res. 2016, 163, 21–31. [Google Scholar] [CrossRef]
- Zhao, S.C.; Li, K.J.; Zhou, W.; Qiu, S.J.; Huang, S.W.; He, P. Changes in soil microbial community, enzyme activities and organic matter fractions under long-term straw return in north-central China. Agric. Ecosyst. Environ. 2016, 216, 82–88. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Adams, R.I.; Miletto, M.; Taylor, J.W.; Bruns, T.D. Dispersal in microbes: Fungi in indoor air are dominated by outdoor air and show dispersal limitation at short distances. ISME J. 2013, 7, 1262–1273. [Google Scholar] [CrossRef] [Green Version]
- Gardes, M.; Bruns, T.D. Its Primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feeley, K.J.; Davies, S.J.; Perez, R.; Hubbell, S.P.; Foster, R.B. Directional changes in the species composition of a tropical forest. Ecology 2011, 92, 871–882. [Google Scholar] [CrossRef] [Green Version]
- Buttigieg, P.L.; Ramette, A. A guide to statistical analysis in microbial ecology: A community-focused, living review of multivariate data analyses. FEMS Microbiol. Ecol. 2014, 90, 543–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, S.H.; Tian, L.; Ma, L.N.; Tian, C.J. Community Structure of Rhizomicrobiomes in Four Medicinal Herbs and Its Im-plication on Growth Management. Microbiology 2018, 87, 425–436. [Google Scholar] [CrossRef]
- Oksanen, J. Multivariate Analysis of Ecological Communities in R: Vegan Tutorial. 2011. Available online: https://www.yumpu.com/en/document/view/41285419/multivariate-analysis-of-ecological-communities-in-r-vegan-tutorial (accessed on 18 June 2021).
- Yang, Y.Y.; Wang, W.B. Benzyldimethyldodecyl ammonium chloride shifts the proliferation of functional genes and microbial community in natural water from eutrophic lake. Environ. Pollut. 2018, 236, 355–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revelle, W.; Revelle, M.W. Package ‘Psych’. The Comprehensive R Archive Network. 2015. Available online: http://www.test.personality-project.org/r/psych/psych-manual.pdf (accessed on 20 January 2021).
- Sun, W.M.; Xiao, E.Z.; Xiao, T.F.; Krumins, V.; Wang, Q.; Haggblom, M. Response of Soil Microbial Communities to Elevated Antimony and Arsenic Contamination Indicates the Relationship between the Innate Microbiota and Contaminant Fractions. Environ. Sci. Technol. 2017, 51, 9165–9175. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.P.; Ma, H.; Zhao, Q.L.; Zhang, S.R.; Wei, W.L.; Ding, X.D. Changes in soil bacterial community and enzyme activity under five years straw returning in paddy soil. Eur. J. Soil Biol. 2020, 100, 103215. [Google Scholar] [CrossRef]
- Yan, S.S.; Song, J.M.; Fan, J.S.; Yan, C.; Dong, S.K.; Ma, C.M. Changes in soil organic carbon fractions and microbial community under rice straw return in Northeast China. Glob. Ecol. Conserv. 2020, 22, e00962. [Google Scholar] [CrossRef]
- Bu, R.Y.; Ren, T.; Lei, M.J.; Liu, B.; Li, X.K.; Cong, R.H. Tillage and straw-returning practices effect on soil dissolved organic matter, aggregate fraction and bacteria community under rice-rice-rapeseed rotation system. Agric. Ecosyst. Environ. 2020, 287, 106681. [Google Scholar] [CrossRef]
- Stemmer, M.; Watzinger, A.; Blochberger, K.; Haberhauer, G.; Gerzabek, M.H. Linking dynamics of soil microbial phospholipid fatty acids to carbon mineralization in a C-13 natural abundance experiment: Impact of heavy metals and acid rain. Soil Biol. Biochem. 2007, 39, 3177–3186. [Google Scholar] [CrossRef]
- Paterson, E.; Sim, A.; Osborne, S.M.; Murray, P.J. Long-term exclusion of plant-inputs to soil reduces the functional capacity of microbial communities to mineralise recalcitrant root-derived carbon sources. Soil Biol. Biochem. 2011, 43, 1873–1880. [Google Scholar] [CrossRef]
- Marschner, P.; Umar, S.; Baumann, K. The microbial community composition changes rapidly in the early stages of decomposition of wheat residue. Soil Biol. Biochem. 2011, 43, 445–451. [Google Scholar] [CrossRef]
- Pastorelli, R.; Vignozzi, N.; Landi, S.; Piccolo, R.; Orsini, R.; Seddaiu, G. Consequences on macroporosity and bacterial diversity of adopting a no-tillage farming system in a clayish soil of Central Italy. Soil Biol. Biochem. 2013, 66, 78–93. [Google Scholar] [CrossRef]
- Song, X.Y.; Spaccini, R.; Pan, G.; Piccolo, A. Stabilization by hydrophobic protection as a molecular mechanism for organic carbon sequestration in maize-amended rice paddy soils. Sci. Total Environ. 2013, 458–460, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.S.; Ge, T.D.; Liu, J.Y.; Chen, X.B.; Hu, Y.J.; Wu, J.S. Effects of biotic and abiotic factors on soil organic matter mineralization: Experiments and structural modeling analysis. Eur. J. Soil Biol. 2018, 84, 27–34. [Google Scholar] [CrossRef]
- de Boer, W.; Folman, L.B.; Summerbell, R.C.; Boddy, L. Living in a fungal world: Impact of fungi on soil bacterial niche development. FEMS Microbiol. Rev. 2005, 29, 795–811. [Google Scholar] [CrossRef] [Green Version]
- Torsvik, V.; Ovreas, L.; Thingstad, T.F. Prokaryotic diversity-Magnitude, dynamics, and controlling factors. Science 2002, 296, 1064–1066. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.J.; Sui, Y.Y.; Yu, Z.H.; Yao, Q.; Shi, Y.; Chu, H.Y. Diversity and distribution patterns of acidobacterial communities in the black soil zone of northeast China. Soil Biol. Biochem. 2016, 95, 212–222. [Google Scholar] [CrossRef]
- Shahbaz, M.; Kuzyakov, Y.; Sanaullah, M.; Heitkamp, F.; Zelenev, V.; Kumar, A. Microbial decomposition of soil organic matter is mediated by quality and quantity of crop residues: Mechanisms and thresholds. Biol. Fertil. Soils 2017, 53, 287–301. [Google Scholar] [CrossRef]
- Han, S.; Zeng, L.Y.; Luo, X.S.; Xiong, X.; Wen, S.L.; Wang, B.R. Shifts in Nitrobacter- and Nitrospira-like nitrite-oxidizing bacterial communities under long-term fertilization practices. Soil Biol. Biochem. 2018, 124, 118–125. [Google Scholar] [CrossRef]
- Ahn, J.H.; Song, J.; Kim, B.Y.; Kim, M.S.; Joa, J.H.; Weon, H.Y. Characterization of the Bacterial and Archaeal Communities in Rice Field Soils Subjected to Long-Term Fertilization Practices. J. Microbiol. 2012, 50, 754–765. [Google Scholar] [CrossRef]
- Gupta, R.S. The phylogeny of proteobacteria: Relationships to other eubacterial phyla and eukaryotes. FEMS Microbiol. Rev. 2000, 24, 367–402. [Google Scholar] [CrossRef]
- Yi, W.J.; You, J.H.; Zhu, C.; Wang, B.L.; Qu, D. Diversity, dynamic and abundance of Geobacteraceae species in paddy soil following slurry incubation. Eur. J. Soil Biol. 2013, 56, 11–18. [Google Scholar] [CrossRef]
- Wisawapipat, W.; Chooaiem, N.; Aramrak, S.; Chittamart, N.; Nookabkaew, S.; Rangkadilok, N. Sulfur amendments to soil decrease inorganic arsenic accumulation in rice grain under flooded and nonflooded conditions: Insights from temporal dynamics of porewater chemistry and solid-phase arsenic solubility. Sci. Total Environ. 2021, 779, 146352. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.L.; Na, S.; Li, F.B.; Hu, H.W. Impact of sulfate and iron oxide on bacterial community dynamics in paddy soil under alternate watering conditions. J. Hazard. Mater. 2021, 408, 124417. [Google Scholar] [CrossRef] [PubMed]
- Wolinska, A.; Kuzniar, A.; Zielenkiewicz, U.; Izak, D.; Szafranek-Nakonieczna, A.; Banach, A. Bacteroidetes as a sensitive biological indicator of agricultural soil usage revealed by a culture-independent approach. Appl. Soil Ecol. 2017, 119, 128–137. [Google Scholar] [CrossRef]
- Lydell, C.; Dowell, L.; Sikaroodi, M.; Gillevet, P.; Emerson, D. A population survey of members of the phylum Bacteroidetes isolated from salt marsh sediments along the East Coast of the United States. Microb. Ecol. 2004, 48, 263–273. [Google Scholar] [CrossRef]
- Jangid, K.; Williams, M.A.; Franzluebbers, A.J.; Sanderlin, J.S.; Reeves, J.H.; Jenkins, M.B. Relative impacts of land-use, management intensity and fertilization upon soil microbial community structure in agricultural systems. Soil Biol. Biochem. 2008, 40, 2843–2853. [Google Scholar] [CrossRef]
- Konishi, M.; Homma, Y.; Kubonouchi, M.; Kato, Y. Agent Used for Promoting Growth of Plant e.g., Tomato, Comprises Microorganism Contained in Bacteroidetes, Microorganism Contained in Chloroflexi, Microorganism Contained in Gemmatimonadetes, and Microorganisms contained in Verrucomicrobia. 2020. Available online: http://apps.webofknowledge.com/full_record.do?colName=DIIDW&recordID=202088488Q&log_event=no&search_mode=GeneralSearch&qid=1&log_event=yes&product=UA&SID=5F1w4BAxnjaP2n2ScBk&viewType=fullRecord&doc=1&page=1 (accessed on 10 September 2020).
- Raio, A.; Puopolo, G. Pseudomonas chlororaphis metabolites as biocontrol promoters of plant health and improved crop yield. World J. Microbiol. Biotechnol. 2021, 37, 99. [Google Scholar] [CrossRef]
- Fiori, A.C.G.; Schwan-Estrada, K.R.F.; Stangarlin, J.R.; Vida, J.B.; Scapim, C.A.; Cruz, M.E.S. Antifungal activity of leaf extracts and essential oils of some medicinal plants against Didymella bryoniae. J. Phytopathol. 2000, 148, 483–487. [Google Scholar] [CrossRef]
- Wei, X.; Wang, X.; Cao, P.; Gao, Z.; Chen, A.J.; Han, J. Microbial Community Changes in the Rhizosphere Soil of Healthy and Rusty Panax ginseng and Discovery of Pivotal Fungal Genera Associated with Rusty Roots. Biomed. Res. Int. 2020, 2020, 8018525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, S.; Nuccio, E.E.; Shi, Z.J.; He, Z.; Zhou, J.; Firestone, M.K. The interconnected rhizosphere: High network complexity dominates rhizosphere assemblages. Ecol. Lett. 2016, 19, 926–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montoya, J.M.; Pimm, S.L.; Sole, R.V. Ecological networks and their fragility. Nature 2006, 442, 259–264. [Google Scholar] [CrossRef]
- Xiao, X.; Liang, Y.T.; Zhou, S.; Zhuang, S.Y.; Sun, B. Fungal community reveals less dispersal limitation and potentially more connected network than that of bacteria in bamboo forest soils. Mol. Ecol. 2018, 27, 550–563. [Google Scholar] [CrossRef]
- Zhang, B.G.; Zhang, J.; Liu, Y.; Shi, P.; Wei, G.H. Co-occurrence patterns of soybean rhizosphere microbiome at a continental scale. Soil Biol. Biochem. 2018, 118, 178–186. [Google Scholar] [CrossRef]
- Berry, D.; Widder, S. Deciphering microbial interactions and detecting keystone species with co-occurrence networks. Front. Microbiol. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]




| Straw Addition | Chao 1 | ACE | Simpson | Shannon | The Number of OTUs | |
|---|---|---|---|---|---|---|
| Bacteria | 0% | 4097.23 ± 19.13 a | 4023.71 ± 153.68 a | 0.9981 ± 0.002 a | 10.48 ± 0.04 a | 3396.67 ± 32.25 a |
| 50% | 3978.75 ± 151.81 a | 4121.79 ± 98.71 a | 0.9977 ± 0.003 a | 10.47 ± 0.10 a | 3355.00 ± 118.25 a | |
| 75% | 4080.96 ± 98.50 a | 3908.46 ± 65.73 a | 0.9969 ± 0.004 a | 10.39 ± 0.02 a | 3390.67 ± 31.20 a | |
| 100% | 3862.76 ± 69.83 a | 4135.83 ± 22.97 a | 0.9970 ± 0.009 a | 10.35 ± 0.08 a | 3288.00 ± 30.34 a | |
| Fungi | 0% | 1041.67 ± 62.40 a | 1041.82 ± 62.35 a | 0.9667 ± 0.002 a | 6.45 ± 0.16 a | 664.33 ± 54.48 a |
| 50% | 830.33 ± 52.09 ab | 830.33 ± 52.09 ab | 0.9457 ± 0.023 a | 5.73 ± 0.40 a | 480.67 ± 36.23 b | |
| 75% | 776.67 ± 56.04 b | 776.67 ± 56.04 b | 0.9107 ± 0.028 a | 5.16 ± 0.32 a | 470.33 ± 45.32 b | |
| 100% | 781.33 ± 161.40 b | 781.50 ± 161.51 b | 0.8656 ± 0.093 a | 5.26 ± 0.93 a | 484.00 ± 113.53 b |
| Kingdom | Phyla | 0% Straw Returning | 50% Straw Returning | 75% Straw Returning | 100% Straw Returning |
|---|---|---|---|---|---|
| Bacteria | Acidobacteria | 0.1387 ± 0.0045 a | 0.1374 ± 0.0025 a | 0.1284 ± 0.0091 ab | 0.119 ± 0.01 b |
| Actinobacteria | 0.1393 ± 0.0066 a | 0.1582 ± 0.0102 a | 0.1479 ± 0.0234 a | 0.1367 ± 0.0201 a | |
| Armatimonadetes | 0.005 ± 0.0014 a | 0.0045 ± 0.0008 a | 0.0053 ± 0.0014 a | 0.0035 ± 0.001 a | |
| Bacteroidetes | 0.0318 ± 0.0039 c | 0.0448 ± 0.0043 b | 0.0489 ± 0.0035 ab | 0.055 ± 0.0021 a | |
| Chlorobi | 0.0134 ± 0.002 a | 0.0083 ± 0.0012 b | 0.0166 ± 0.0021 a | 0.017 ± 0.0022 a | |
| Chloroflexi | 0.1641 ± 0.0197 ab | 0.1402 ± 0.0126 b | 0.1798 ± 0.0191 a | 0.1482 ± 0.0101 ab | |
| Cyanobacteria | 0.0082 ± 0.0037 ab | 0.0105 ± 0.0027 a | 0.0035 ± 0.0009 b | 0.0036 ± 0.0003 b | |
| Elusimicrobia | 0.0016 ± 0.0002 a | 0.0013 ± 0.0003 a | 0.0012 ± 0.0004 a | 0.0012 ± 0.0002 a | |
| Fibrobacteres | 0.0001 ± 0.0001 b | 0.0004 ± 0.0001 a | 0.0002 ± 0 b | 0.0003 ± 0.0001 b | |
| Firmicutes | 0.0052 ± 0.0004 b | 0.0075 ± 0.0009 b | 0.024 ± 0.0112 a | 0.0096 ± 0.0004 b | |
| Gemmatimonadetes | 0.0523 ± 0.0021 a | 0.0357 ± 0.0041 b | 0.0372 ± 0.0062 b | 0.0349 ± 0.0028 b | |
| Nitrospirae | 0.0316 ± 0.0079 a | 0.0207 ± 0.0032 ab | 0.0261 ± 0.0062 ab | 0.0176 ± 0.0024 b | |
| Planctomycetes | 0.0017 ± 0.0002 b | 0.0028 ± 0.0007 a | 0.0012 ± 0.0002 b | 0.0011 ± 0.0002 b | |
| Proteobacteria | 0.3929 ± 0.0223 bc | 0.4137 ± 0.0063 ab | 0.3675 ± 0.0213 c | 0.4382 ± 0.0147 a | |
| Spirochaetes | 0.0016 ± 0.0001 a | 0.0014 ± 0.0004 a | 0.0018 ± 0.0003 a | 0.0022 ± 0.0008 a | |
| Verrucomicrobia | 0.0125 ± 0.0029 a | 0.0127 ± 0.001 a | 0.0104 ± 0.0037 a | 0.0119 ± 0.0036 a | |
| Fungi | Ascomycota | 0.5885 ± 0.1015 a | 0.5954 ± 0.0183 a | 0.7125 ± 0.1118 a | 0.5713 ± 0.1201 a |
| Basidiomycota | 0.2583 ± 0.0571 a | 0.3034 ± 0.0485 a | 0.25 ± 0.0951 a | 0.3282 ± 0.1343 a | |
| Chytridiomycota | 0.0004 ± 0.0002 a | 0.0002 ± 0 a | 0.0001 ± 0.0001 a | 0.0003 ± 0.0002 a | |
| Mortierellomycota | 0.1487 ± 0.0459 a | 0.0943 ± 0.0343 ab | 0.0309 ± 0.0142 b | 0.0881 ± 0.0105 ab | |
| Mucoromycota | 0.0001 ± 0 b | 0 ± 0 b | 0.0002 ± 0.0001 b | 0.0106 ± 0.008 a | |
| Rozellomycota | 0.0039 ± 0.0012 ab | 0.0067 ± 0.001 a | 0.0062 ± 0.0033 a | 0.0014 ± 0.0009 b |
| Kingdom | Phylum | Genus | 0% Straw Returning | 50% Straw Returning | 75% Straw Returning | 100% Straw Returning |
|---|---|---|---|---|---|---|
| Bacteria | Actinobacteria | Arthrobacter | 0.1182 ± 0.0041 a | 0.1641 ± 0.0128 a | 0.1729 ± 0.0232 a | 0.15 ± 0.0435 a |
| Terracoccus | 0.0193 ± 0.0006 a | 0.0135 ± 0.0013 b | 0.0159 ± 0.0008 b | 0.0104 ± 0.0021 c | ||
| Nocardioides | 0.042 ± 0.0019 a | 0.0403 ± 0.004 ab | 0.0354 ± 0.0024 b | 0.0273 ± 0.0016 c | ||
| Acidobacteria | Candidatussolibacter | 0.0436 ± 0.0041 a | 0.0337 ± 0.0027 ab | 0.0287 ± 0.0068 b | 0.0254 ± 0.0036 b | |
| Candidatuskoribacter | 0.0317 ± 0.0019 b | 0.0381 ± 0.0029 a | 0.0256 ± 0.0014 c | 0.0206 ± 0.0036 c | ||
| Geothrix | 0.0058 ± 0.0007 b | 0.0086 ± 0.0015 b | 0.0129 ± 0.0016 a | 0.0093 ± 0.0017 b | ||
| Bacteroidetes | Flavobacterium | 0.0045 ± 0.0022 b | 0.0245 ± 0.0078 a | 0.0202 ± 0.0054 a | 0.0262 ± 0.0049 a | |
| Paludibacter | 0.0003 ± 0.0002 c | 0.0006 ± 0.0003 c | 0.0136 ± 0.0025 a | 0.0049 ± 0.0006 b | ||
| Chloroflexi | Anaerolinea | 0.0393 ± 0.0147 a | 0.0328 ± 0.0066 a | 0.0404 ± 0.0144 a | 0.0321 ± 0.0124 a | |
| Cyanobacteria | Leptolyngbya | 0.0076 ± 0.0012 b | 0.0172 ± 0.005 a | 0.0045 ± 0.001 b | 0.006 ± 0.0025 b | |
| Firmicutes | Clostridium | 0.0058 ± 0.001 b | 0.0074 ± 0.001 b | 0.012 ± 0.0024 a | 0.0084 ± 0.0015 ab | |
| Nitrospirae | Nitrospira | 0.0248 ± 0.0047 a | 0.0157 ± 0.0044 b | 0.0127 ± 0.0025 b | 0.0121 ± 0.0029 b | |
| Proteobacteria | Geobacter | 0.0951 ± 0.0083 b | 0.074 ± 0.0126 b | 0.1459 ± 0.0044 a | 0.1461 ± 0.0303 a | |
| Crenothrix | 0.0044 ± 0.0006 b | 0.0039 ± 0.0003 b | 0.0089 ± 0.0008 b | 0.0568 ± 0.0076 a | ||
| Anaeromyxobacter | 0.0675 ± 0.0055 a | 0.0613 ± 0.0073 a | 0.0726 ± 0.0063 a | 0.0595 ± 0.006 a | ||
| Rhodoplanes | 0.0912 ± 0.0016 a | 0.0569 ± 0.0081 b | 0.0428 ± 0.0064 b | 0.0504 ± 0.0081 b | ||
| Pseudomonas | 0.0019 ± 0.0007 b | 0.0087 ± 0.0027 ab | 0.0122 ± 0.0033 ab | 0.0228 ± 0.012 a | ||
| Methylosinus | 0.0208 ± 0.0025 b | 0.0358 ± 0.005 a | 0.0196 ± 0.0034 b | 0.0279 ± 0.0031 ab | ||
| Syntrophobacter | 0.0568 ± 0.0014 a | 0.053 ± 0.0061 a | 0.0317 ± 0.0036 b | 0.0347 ± 0.004 b | ||
| Methylibium | 0.0189 ± 0.0026 a | 0.0167 ± 0.0012 a | 0.0094 ± 0.0019 b | 0.0156 ± 0.0011 a | ||
| Desulfobacca | 0.0265 ± 0.0034 a | 0.0219 ± 0.0028 a | 0.024 ± 0.0037 a | 0.014 ± 0.0009 b | ||
| Phenylobacterium | 0.0164 ± 0.0032 b | 0.0257 ± 0.0031 a | 0.0078 ± 0.0014 c | 0.0138 ± 0.0017 b | ||
| Variovorax | 0.0093 ± 0.001 a | 0.0116 ± 0.0005 a | 0.0087 ± 0.0033 a | 0.0117 ± 0.0006 a | ||
| Thiobacillus | 0.0338 ± 0.0156 a | 0.0135 ± 0.0038 a | 0.0166 ± 0.0031 a | 0.019 ± 0.0061 a | ||
| Reyranella | 0.0139 ± 0.0024 a | 0.0096 ± 0.0007 b | 0.0049 ± 0.0009 c | 0.0085 ± 0.0009 b | ||
| Hyphomicrobium | 0.0115 ± 0.0006 a | 0.0103 ± 0.0009 a | 0.0076 ± 0.0007 b | 0.0068 ± 0.0006 b | ||
| Fungi | Ascomycota | Pseudogymnoascus | 0.1055 ± 0.0144 ab | 0.1369 ± 0.0467 ab | 0.2349 ± 0.1072 a | 0.0276 ± 0.0104 b |
| Didymella | 0.1692 ± 0.0623 a | 0.055 ± 0.0224 b | 0.0748 ± 0.0394 b | 0.019 ± 0.0053 b | ||
| Echria | 0.0392 ± 0.0074 b | 0.1942 ± 0.0648 a | 0.0353 ± 0.0227 b | 0.0261 ± 0.0202 b | ||
| Setophoma | 0.0073 ± 0.0044 b | 0.0068 ± 0.0049 b | 0.205 ± 0.163 a | 0.0036 ± 0.0008 b | ||
| Fusicolla | 0.0001 ± 0 b | 0 ± 0 b | 0.0003 ± 0.0003 b | 0.3394 ± 0.2546 a | ||
| Candida | 0.0253 ± 0.0061 a | 0.0098 ± 0.0018 b | 0.0122 ± 0.003 b | 0.0125 ± 0.0057 b | ||
| Xenopolyscytalum | 0.0129 ± 0.0063 a | 0.0048 ± 0.0017 b | 0.0013 ± 0.0002 b | 0.0037 ± 0.0022 b | ||
| Articulospora | 0.0641 ± 0.0382 a | 0.0002 ± 0.0001 b | 0.0002 ± 0.0001 b | 0.016 ± 0.0102 b | ||
| Pyrenochaetopsis | 0.0369 ± 0.0186 a | 0.0268 ± 0.0132 a | 0.0126 ± 0.0015 a | 0.0218 ± 0.0066 a | ||
| Cladosporium | 0.0232 ± 0.0064 a | 0.0142 ± 0.007 a | 0.0449 ± 0.0364 a | 0.0168 ± 0.0051 a | ||
| Microdochium | 0.0139 ± 0.004 a | 0.0035 ± 0.0013 b | 0.0041 ± 0.0022 b | 0.0037 ± 0.0016 b | ||
| Nigrospora | 0.0177 ± 0.0071 a | 0.0069 ± 0.002 b | 0.0027 ± 0.0007 b | 0.0079 ± 0.005 ab | ||
| Aspergillus | 0.0064 ± 0.0026 a | 0.0022 ± 0.0006 a | 0.0047 ± 0.0021 a | 0.0028 ± 0.001 a | ||
| Talaromyces | 0.0033 ± 0.0017 a | 0.0012 ± 0.0002 b | 0.001 ± 0.0003 b | 0.0007 ± 0.0003 b | ||
| Basidiomycota | Papiliotrema | 0.0079 ± 0.003 b | 0.0072 ± 0.0022 b | 0.0122 ± 0.0037 ab | 0.0225 ± 0.0088 a | |
| Guehomyces | 0.1033 ± 0.0324 a | 0.0894 ± 0.0164 a | 0.08 ± 0.0305 a | 0.1095 ± 0.0667 a | ||
| Solicoccozyma | 0.0559 ± 0.0183 a | 0.042 ± 0.0013 a | 0.0661 ± 0.0246 a | 0.1009 ± 0.0566 a | ||
| Psilocybe | 0.0423 ± 0.0153 a | 0.1351 ± 0.0966 a | 0.017 ± 0.0063 a | 0.0258 ± 0.014 a | ||
| Mrakia | 0.0283 ± 0.0065 a | 0.0294 ± 0.0034 a | 0.0713 ± 0.0351 a | 0.0619 ± 0.0257 a | ||
| Mortierellomycota | Mortierella | 0.16 ± 0.05 a | 0.102 ± 0.0345 ab | 0.0312 ± 0.0135 b | 0.0681 ± 0.0135 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, E.; Lin, X.; Tian, L.; Wang, X.; Ji, L.; Jin, F.; Tian, C. Effects of Short-Term Rice Straw Return on the Soil Microbial Community. Agriculture 2021, 11, 561. https://doi.org/10.3390/agriculture11060561
Wang E, Lin X, Tian L, Wang X, Ji L, Jin F, Tian C. Effects of Short-Term Rice Straw Return on the Soil Microbial Community. Agriculture. 2021; 11(6):561. https://doi.org/10.3390/agriculture11060561
Chicago/Turabian StyleWang, Enze, Xiaolong Lin, Lei Tian, Xinguang Wang, Li Ji, Feng Jin, and Chunjie Tian. 2021. "Effects of Short-Term Rice Straw Return on the Soil Microbial Community" Agriculture 11, no. 6: 561. https://doi.org/10.3390/agriculture11060561