
Enhanced Ruminal Fermentation Parameters and Altered Rumen Bacterial Community Composition by Formulated Rumen Buffer Agents Fed to Dairy Cows with a High-Concentrate Diet


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care
2.2. Experiment 1
2.2.1. Animals, Feeding, and Experimental Design
2.2.2. Rumen Fluid Collection and Analyses for In Vitro Rumen Fermentation and Buffering Capacity
2.3. Experiment 2
2.3.1. Formulation of Buffer Agents Used for In Vitro and In Vivo Experiments
2.3.2. Rumen Fluid Collection, In Vitro Rumen Fermentation, Buffering Capacity, and Parameter Analyses
2.3.3. Experimental Animals, Feeding, Design, and Effect of Treatments on the In Vivo Study
2.3.4. 16S rRNA Amplicon Sequencing and Metataxonomic Analyses
2.4. Statistical Analysis
3. Results
3.1. Experiment 1
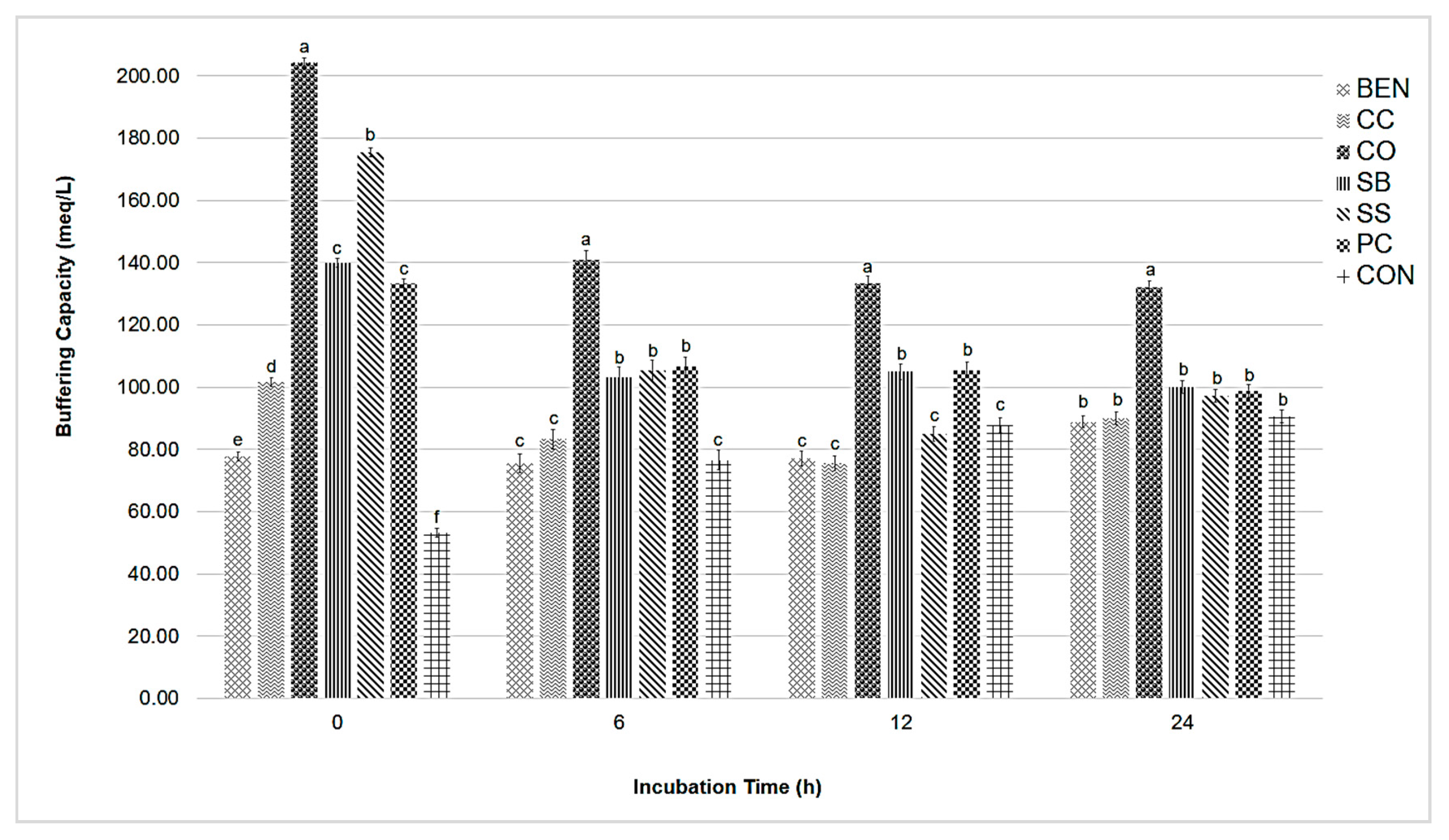
Effects of Individual Buffer Agents on In Vitro Rumen Fermentation Parameters
3.2. Experiment 2
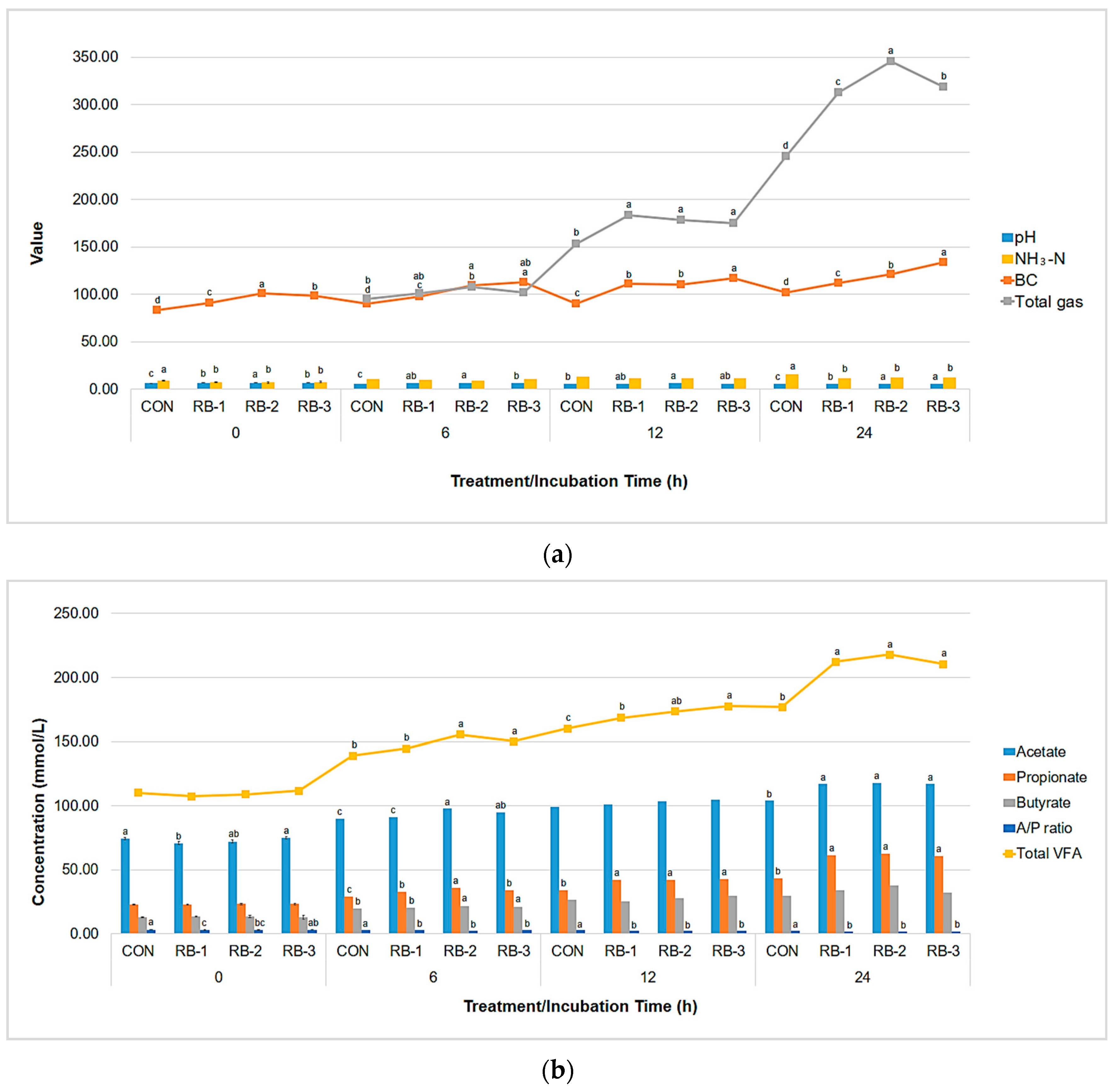
3.2.1. Effect of Combination of Buffer Agents on In Vitro Rumen Fermentation Parameters
3.2.2. Effects of Combination of Buffer Agents on In Vivo Rumen Fermentation Parameters
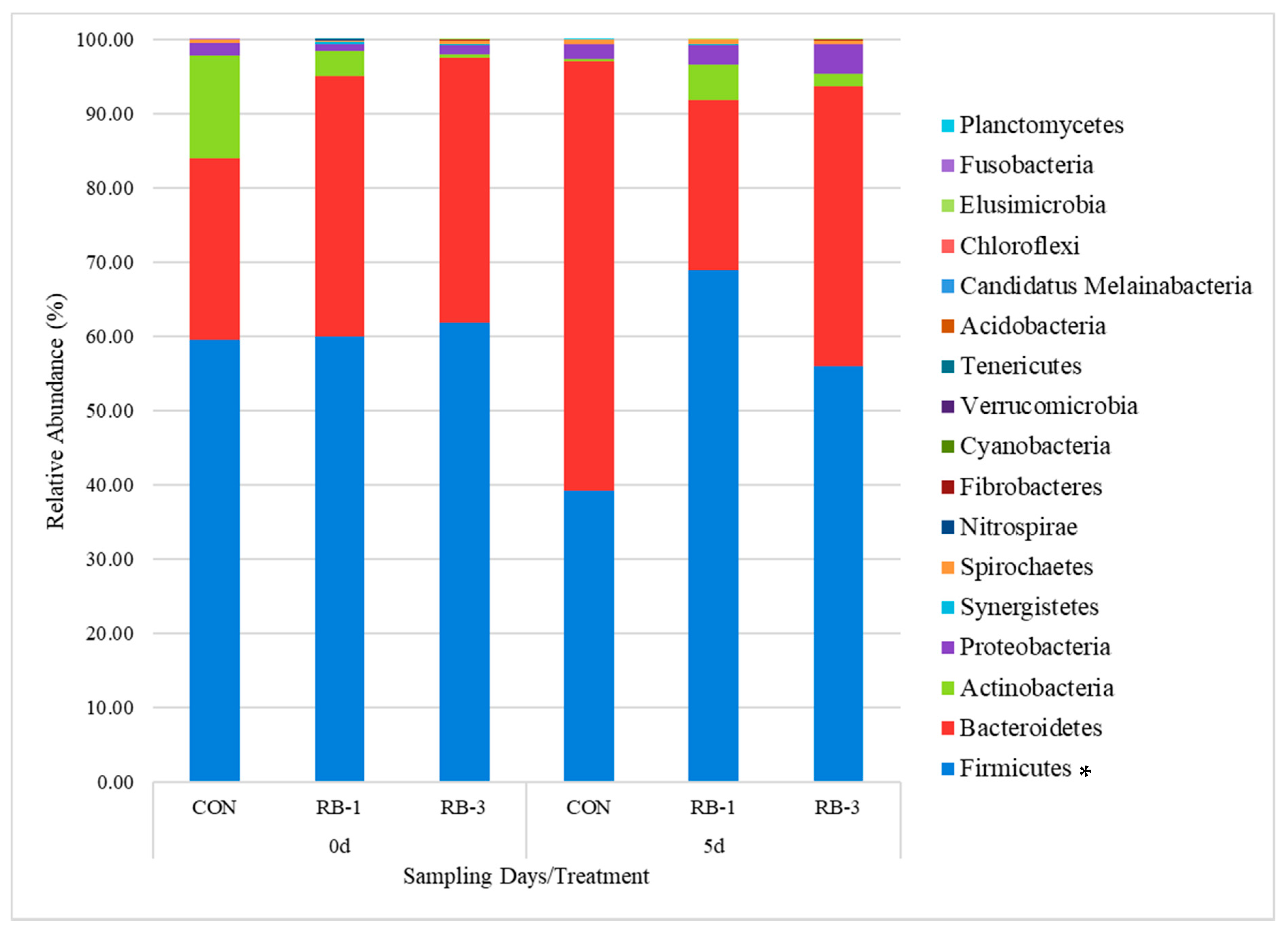
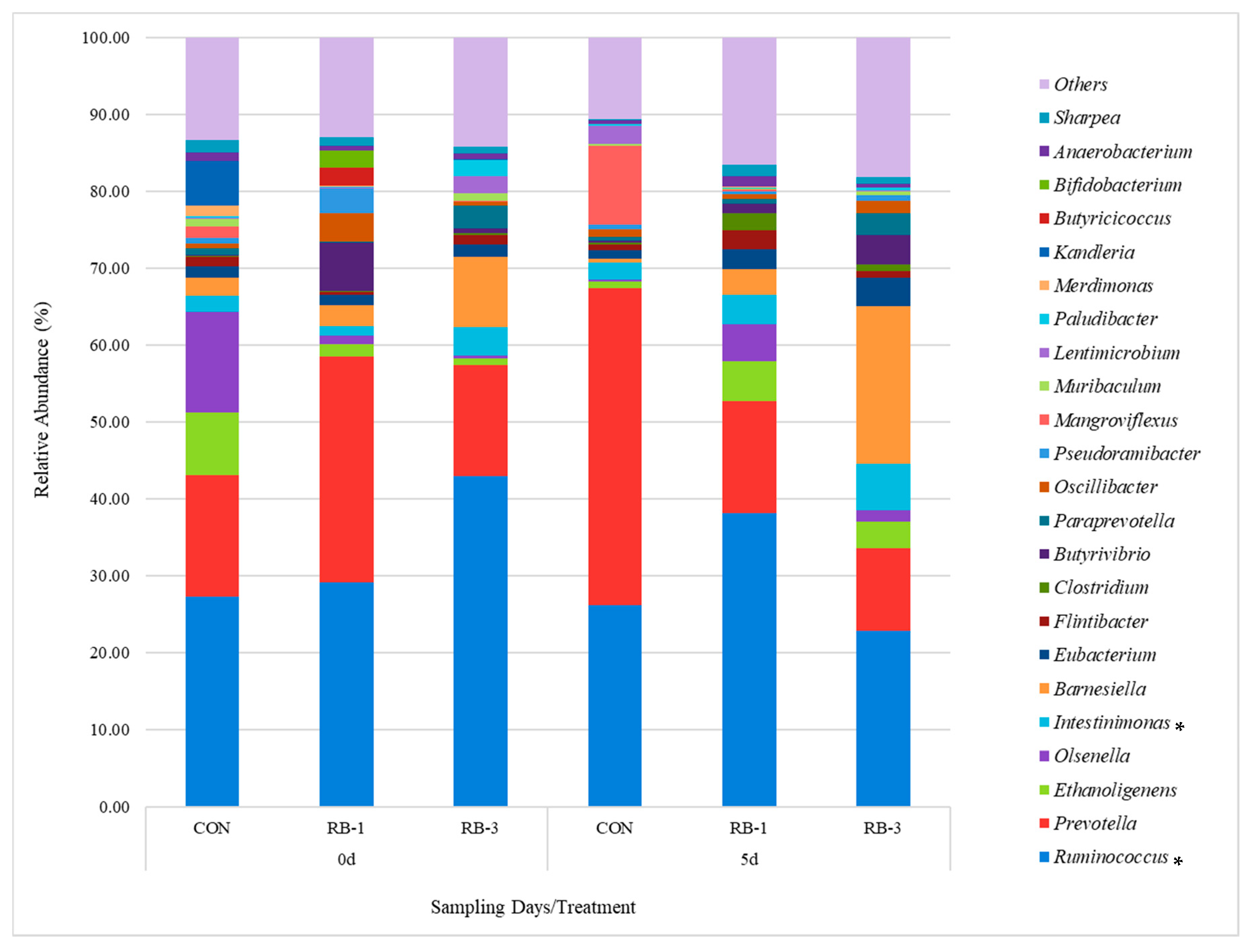
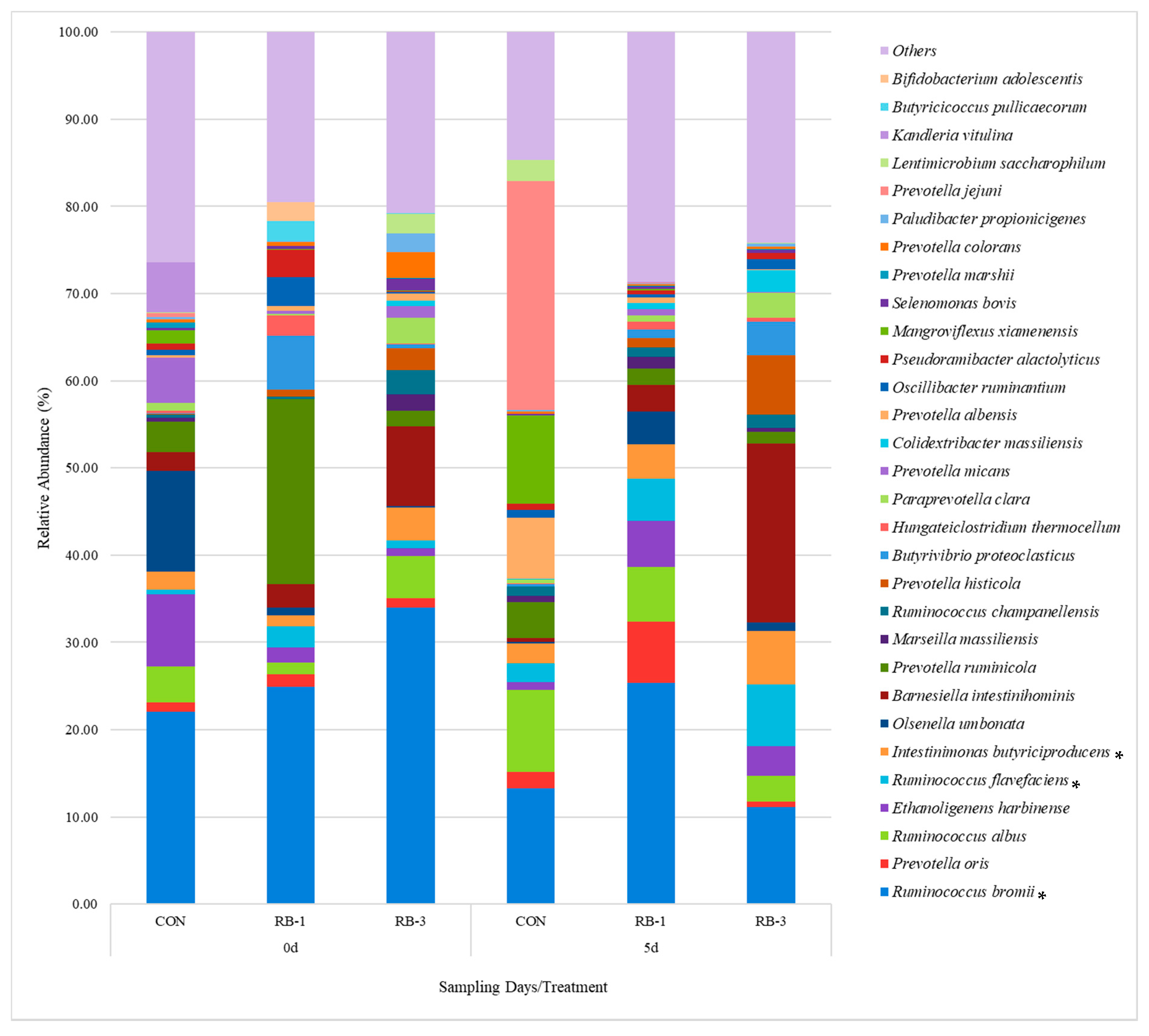
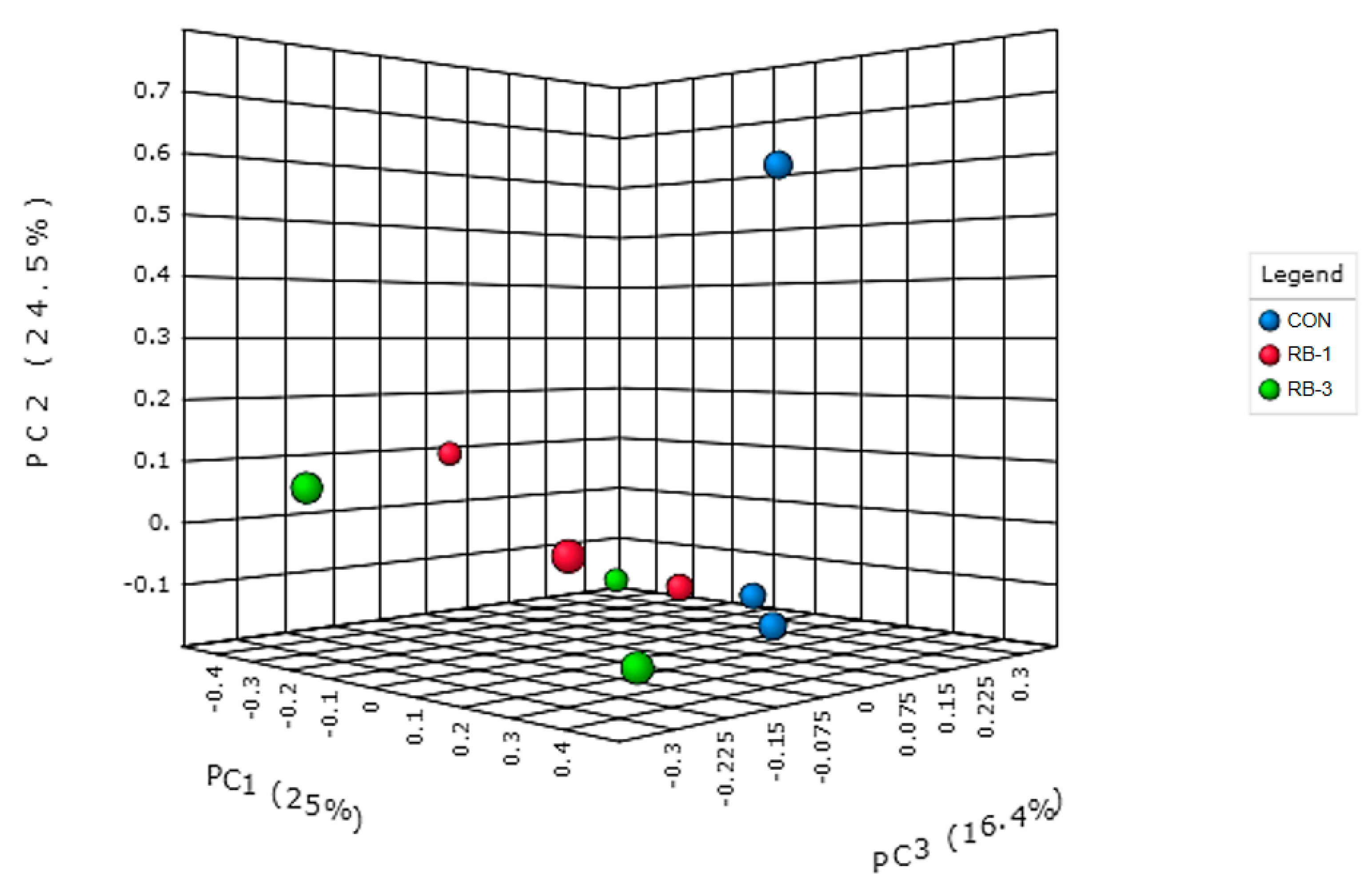
3.2.3. General Rumen Bacterial Community Composition
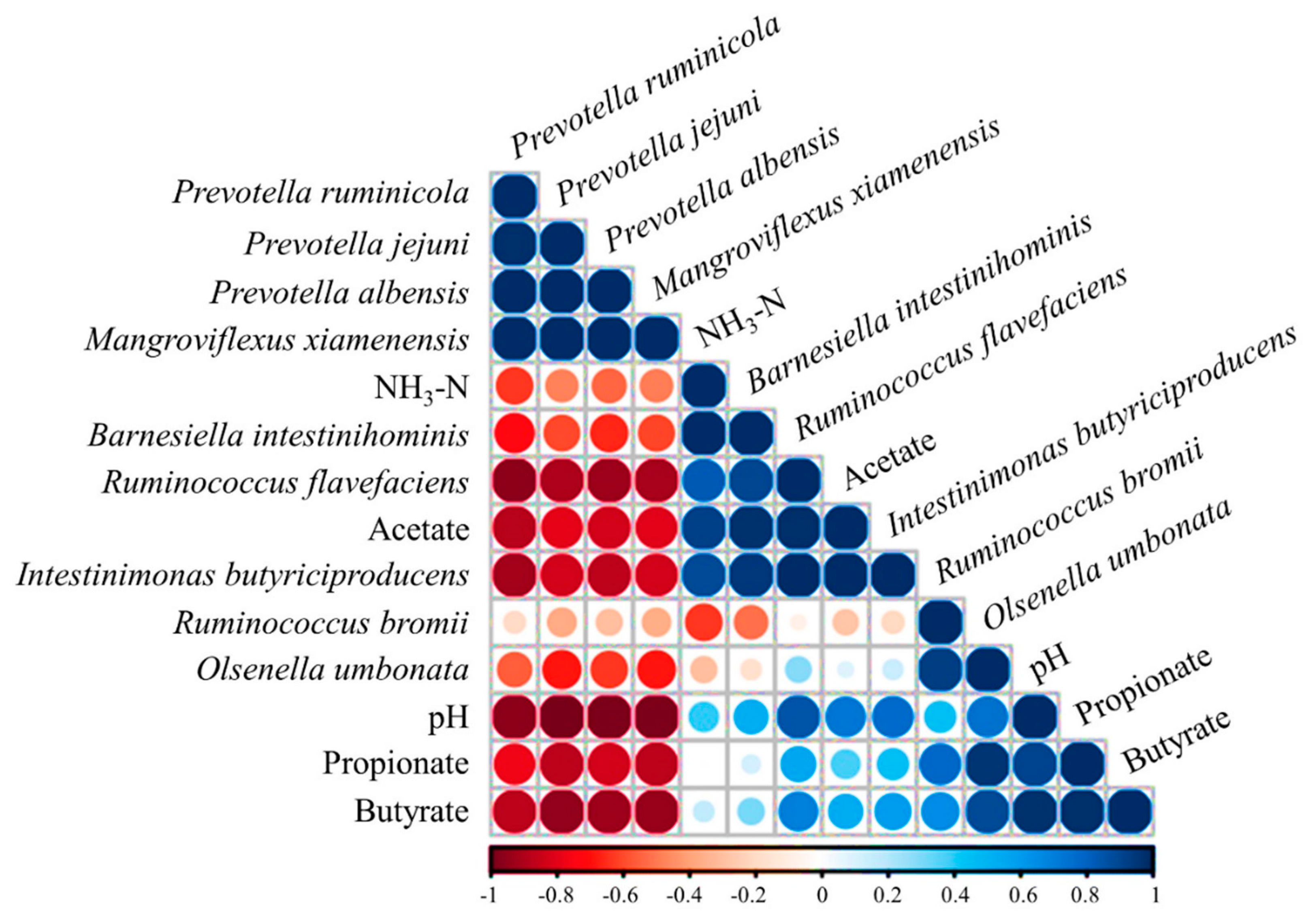
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hook, S.E.; Steele, M.A.; Northwood, K.S.; Dijkstra, J.; France, J.; Wright, A.-D.G.; McBride, B.W. Impact of subacute ruminal acidosis (SARA) adaptation and recovery on the density and diversity of bacteria in the rumen of dairy cows. FEMS Microbiol. Ecol. 2011, 78, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Plaizier, J.C.; Li, S.; Danscher, A.M.; Derakshani, H.; Andersen, P.H.; Khafipour, E. Changes in Microbiota in Rumen Digesta and Feces Due to a Grain-Based Subacute Ruminal Acidosis (SARA) Challenge. Microb. Ecol. 2017, 74, 485–495. [Google Scholar] [CrossRef]
- Nagata, R.; Kim, Y.H.; Ohkubo, A.; Kushibiki, S.; Ichijo, T.; Sato, S. Effects of repeated subacute ruminal acidosis challenges on the adaptation of the rumen bacterial community in Holstein bulls. J. Dairy Sci. 2018, 101, 4424–4436. [Google Scholar] [CrossRef]
- Plaizier, J.C.; Khafipour, E.; Li, S.; Gozho, G.N.; Krause, D.O. Subacute ruminal acidosis (SARA), endotoxins and health consequences. Anim. Feed Sci. Technol. 2012, 172, 9–21. [Google Scholar] [CrossRef]
- Kleen, J.L.; Cannizzo, C. Incidence, prevalence and impact of SARA in dairy herds. Anim. Feed Sci. Technol. 2012, 172, 4–8. [Google Scholar] [CrossRef]
- Plaizier, J.C.; Krause, D.O.; Gozho, G.N.; McBride, B.W. Subacute ruminal acidosis in dairy cows: The physiological causes, incidence and consequences. Vet. J. 2008, 176, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Rabelo, E.; Rezende, R.L.; Bertics, S.J.; Grummer, R.R. Effects of transition diets varying in dietary energy density on lactation performance and ruminal parameters of dairy cows. J. Dairy Sci. 2003, 86, 916–925. [Google Scholar] [CrossRef] [Green Version]
- Nagaraja, T.G.; Titgemeyer, E.C. Ruminal acidosis in beef cattle: The current microbiological and nutritional outlook. J. Dairy Sci. 2007, 90, E17–E38. [Google Scholar] [CrossRef] [Green Version]
- Petri, R.M.; Schwaiger, T.; Penner, G.B.; Beauchemin, K.A.; Forster, R.J.; McKinnon, J.J.; McAllister, T.A. Characterization of the core rumen microbiome in cattle during transition from forage to concentrate as well as during and after an acidotic challenge. PLoS ONE 2013, 8, e83424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, S.Y.; Zhang, R.Y.; Wang, D.S.; Zhu, W.Y. Impact of subacute ruminal acidosis (SARA) adaptation on rumen microbiota in dairy cattle using pyrosequencing. Anaerobe 2013, 24, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Khafipour, E.; Li, S.; Tun, H.M.; Derakhshani, H.; Moossavi, S.; Plaizier, K.J.C. Effects of grain feeding on microbiota in the digestive tract of cattle. Anim. Front. 2016, 6, 13–19. [Google Scholar] [CrossRef]
- Steele, M.A.; Croom, J.; Kahler, M.; Alzahal, O.; Hook, S.E.; Plaizier, K.; Mcbride, B.W. Bovine rumen epithelium undergoes rapid structural adaptations during grain-induced subacute ruminal acidosis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R1515–R1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zebeli, Q.; Metzler-Zebeli, B.U. Interplay between rumen digestive disorders and diet-induced inflammation in dairy cattle. Res. Vet. Sci. 2012, 93, 1099–1108. [Google Scholar] [CrossRef]
- Russell, J.B.; Rychlik, J.L. Factors that alter rumen microbial ecology. Science (80-.) 2001, 292, 1119–1122. [Google Scholar] [CrossRef]
- Krause, D.O.; Nagaraja, T.G.; Wright, A.D.G.; Callaway, T.R. Board-invited review: Rumen microbiology: Leading the way in microbial ecology. J. Anim. Sci. 2013, 91, 331–341. [Google Scholar] [CrossRef]
- Grant, R.H.; Mertens, D.R. Influence of Buffer pH and Raw Corn Starch Addition on In Vitro Fiber Digestion Kinetics. J. Dairy Sci. 1992, 75, 2762–2768. [Google Scholar] [CrossRef]
- Russell, J.B.; Wilson, D.B. Why Are Ruminal Cellulolytic Bacteria Unable to Digest Cellulose at Low pH? J. Dairy Sci. 1996, 79, 1503–1509. [Google Scholar] [CrossRef]
- Li, F.; Cao, Y.; Liu, N.; Yang, X.; Yao, J.; Yan, D. Subacute ruminal acidosis challenge changed in situ degradability of feedstuffs in dairy goats. J. Dairy Sci. 2014, 97, 5101–5109. [Google Scholar] [CrossRef] [Green Version]
- Khorasani, R. Dietary Buffering Requirements of Lactating Dairy Cows; Department of Agricultural Food and Nutritional Sciences, University of Alberta: Edmonton, AB, Canada, 2018. [Google Scholar]
- Tucker, W.B.; Aslam, M.; Lema, M.; Shin, I.S.; Le Ruyet, P.; Hogue, J.F.; Buchanan, D.S.; Miller, T.P.; Adams, G.D. Sodium bicarbonate or multielement buffer via diet or rumen: Effects on performance and acid-base status of lactating cows. J. Dairy Sci. 1992, 75, 2409–2420. [Google Scholar] [CrossRef]
- Zamarreño, A.M.; García-Mina, J.M.; Cantera, R.G. A new methodology for studying the performance of products against ruminal acidosis. J. Sci. Food Agric. 2003, 83, 1607–1612. [Google Scholar] [CrossRef]
- Stokes, M.R.; Vandemark, L.L.; Bull, L.S. Effects of sodium bicarbonate, magnesium oxide, and a commercial buffer mixture in early lactation cows fed hay crop silage. J. Dairy Sci. 1986, 69, 1595–1603. [Google Scholar] [CrossRef]
- Stallings, C.C. Sodium Bicarbonate and Magnesium Oxide in Dairy Cattle Rations; Virginia Cooperative Extension Program: Blacksburg, VA, USA, 1992; Available online: https://vtechworks.lib.vt.edu/bitstream/handle/10919/75835/VCE404_114.pdf?sequence=1&isAllowed=y (accessed on 12 June 2020).
- Erdman, R.A.; Douglass, L.W.; Hemken, R.W.; Teh, T.H.; Mann, L.M. Effects of Sodium Bicarbonate on Palatability and Voluntary Intake of Concentrates Fed Lactating Dairy Cows. J. Dairy Sci. 1982, 65, 1647–1651. [Google Scholar] [CrossRef]
- Shaver, R.D.; Armentano, L.E.; Crowley, J.W. Dietary Buffers for Dairy Cattle; University of Wisconsin: Madison, WA, USA, 1988. [Google Scholar]
- Herod, E.L.; Bechtle, R.M.; Bartley, E.E.; Dayton, A.D. Buffering ability of several compounds in vitro and the effect of a selected buffer combination on ruminal acid production in vivo. J. Dairy Sci. 1978, 61, 1114–1122. [Google Scholar] [CrossRef]
- Ramos, S.C.; Jeong, C.D.; Mamuad, L.L.; Kim, S.H.; Kang, S.H.; Kim, E.T.; Cho, Y.I.; Lee, S.S.; Lee, S.S. Diet transition from high-forage to high-concentrate alters rumen bacterial community composition, epithelial transcriptomes and ruminal fermentation parameters in dairy cows. Animals 2021, 11, 838. [Google Scholar] [CrossRef]
- Hattori, K.; Matsui, H. Diversity of fumarate reducing bacteria in the bovine rumen revealed by culture dependent and independent approaches. Anaerobe 2008, 14, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Soriano, A.P.; Mamuad, L.L.; Kim, S.H.; Choi, Y.J.; Jeong, C.D.; Bae, G.S.; Chang, M.B.; Lee, S.S. Effect of Lactobacillus mucosae on in vitro rumen fermentation characteristics of dried brewers grain, methane production and bacterial diversity. Asian-Australasian J. Anim. Sci. 2014, 27, 1562–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamuad, L.L.; Kim, S.H.; Choi, Y.J.; Soriano, A.P.; Cho, K.K.; Lee, K.; Bae, G.S.; Lee, S.S. Increased propionate concentration in Lactobacillus mucosae–fermented wet brewers grains and during in vitro rumen fermentation. J. Appl. Microbiol. 2017, 123, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Chaney, A.L.; Marbach, E.P. Modified reagents for determination of urea and ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [CrossRef]
- Le Ruyet, P.; Tucker, W.B. Ruminal Buffers: Temporal Effects on Buffering Capacity and pH of Ruminal Fluid from Cows Fed a High Concentrate Diet. J. Dairy Sci. 1992, 75, 1069–1077. [Google Scholar] [CrossRef]
- Buffalo, V.S. Scythe: A Bayesian Adapter Trimer. Available online: https://github.com/vsbuffalo/scythe (accessed on 22 October 2020).
- Joshi, N.A.; Fass, J.N. Sickle: A Sliding-Window, Adaptive, Quality-Based Trimming Tool for FastQ Files (Version 1.33) [Software]. Available online: https://github.com/najoshi/sickle (accessed on 22 October 2020).
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high- throughput community sequencing data Intensity normalization improves color calling in SOLiD sequencing. Nat. Publ. Gr. 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- McDonald, D.; Clemente, J.C.; Kuczynski, J.; Rideout, J.R.; Stombaugh, J.; Wendel, D.; Wilke, A.; Huse, S.; Hufnagle, J.; Meyer, F.; et al. The Biological Observation Matrix (BIOM) format or: How I learned to stop worrying and love the ome-ome. Gigascience 2012, 464, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Xu, L.; Gu, Y.Q.; Coleman-Derr, D. MetaCoMET: A web platform for discovery and visualization of the core microbiome. Bioinformatics 2016, 32, 3469–3470. [Google Scholar] [CrossRef] [Green Version]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassambara, A. Correlation Matrix—Online Software: Analysis and Visualization. Available online: http://www.sthda.com/english/rsthda/correlation-matrix.php (accessed on 6 June 2021).
- Mainardi, S.R.; Hengst, B.A.; Nebzydoski, S.J.; Nemec, L.M.; Gressley, T.F. Effects of abomasal oligofructose on blood and feces of Holstein steers. J. Anim. Sci. 2011, 89, 2510–2517. [Google Scholar] [CrossRef]
- Teh, T.H.; Hemken, R.W.; Harmon, R.J. Dietary magnesium oxide interactions with sodium bicarbonate on cows in early lactation. J. Dairy Sci. 1985, 68, 881–890. [Google Scholar] [CrossRef]
- Blignaut, D. Alkaline treatment for preventing acidosis in the rumen culture fermenting carbohydrates: An experimental study in vitro. J. Adv. Vet. Anim. Res. 2019, 6, 100–107. [Google Scholar]
- Solorzano, L.C.; Armentano, L.E.; Grummer, R.R.; Dentine, M.R. Effects of sodium bicarbonate or sodium sesquicarbonate on lactating holsteins fed a high grain diet. J. Dairy Sci. 1989, 72, 453–461. [Google Scholar] [CrossRef]
- Snyder, T.J.; Rogers, J.A.; Muller, L.D. Effects of 1.2% sodium icarbonate with two ratios of corn silage:grain on milk production, rumen germentation, and utrient digestion by lactating dairy cows. J. Dairy Sci. 1983, 66, 1290–1297. [Google Scholar] [CrossRef]
- Abdoun, K.; Stumpff, F.; Martens, H. Ammonia and urea transport across the rumen epithelium: A review. Anim. Health Res. Rev. 2006, 7, 43–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohn, R.A.; Dunlap, T.F. Calculation of the buffering capacity of bicarbonate in the rumen and in vitro. J. Anim. Sci. 1998, 76, 1702–1709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emery, R.S.; Brown, L.D. Effect of feeding sodium and potassium bicarbonate on milk fat, rumen pH, and volatile fatty acid production. J. Dairy Sci. 1961, 44, 1899. [Google Scholar] [CrossRef]
- Emery, R.S.; Brown, L.D.; Thomas, J.W. Effect of sodium and calcium carbonates on milk production and composition of milk, blood, and rumen contents of cows fed grain ad libitum with restricted roughage. J. Dairy Sci. 1964, 58, 1325. [Google Scholar] [CrossRef]
- Erdman, R.A.; Botts, R.L.; Hemken, R.W.; Bull, L.S. Effect of dietary sodium bicarbonate and magnesium oxide on production and physiology in early lactation. J. Dairy Sci. 1980, 63, 923–930. [Google Scholar] [CrossRef]
- Wohlt, J.E.; Sniffen, C.J.; Hoover, W.H. Measurement of protein solubility in common feedstuffs. J. Dairy Sci. 1973, 56, 1052. [Google Scholar] [CrossRef]
- Nuñez, A.J.C.; Felix, T.L.; Lemenager, R.P.; Schoonmaker, J.P. Effect of calcium oxide inclusion in beef feedlot diets containing 60% dried distillers grains with solubles on ruminal fermentation, diet digestibility, performance, and carcass characteristics. J. Anim. Sci. 2014, 92, 3954–3965. [Google Scholar] [CrossRef]
- Rauch, R.E.; Robinson, P.H.; Erasmus, L.J. Effects of sodium bicarbonate and calcium magnesium carbonate supplementation on performance of high producing dairy cows. Anim. Feed Sci. Technol. 2012, 177, 180–193. [Google Scholar] [CrossRef]
- Kang, S.; Wanapat, M. Rumen-buffering capacity using dietary sources and in vitro gas fermentation. Anim. Prod. Sci. 2016, 58, 862–870. [Google Scholar] [CrossRef]
- Khattab, I.M.; Salem, A.Z.M.; Abdel-Wahed, A.M.; Kewan, K.Z. Effects of urea supplementation on nutrient digestibility, nitrogen utilisation and rumen fermentation in sheep fed diets containing dates. Livest. Sci. 2013, 155, 223–229. [Google Scholar] [CrossRef]
- Erdman, R.A.; Hemken, R.W.; Bull, L.S. Dietary sodium bicarbonate and magnesium oxide for early postpartum lactating dairy cows: Effects on production, acid-base metabolism, and digestion. J. Dairy Sci. 1982, 65, 712–731. [Google Scholar] [CrossRef]
- Thomas, J.W.; Emery, R.S. Additive Nature of Sodium Bicarbonate and Magnesium Oxide on Milk Fat Concentrations of Milking Cows Fed Restricted-Roughage Rations. J. Dairy Sci. 1969, 52, 1762. [Google Scholar] [CrossRef]
- Thomson, D.J.; Beever, D.E.; Latham, M.J.; Sharpe, M.E. The effect of inclusion of mineral salts in the diet on dilution rate, the pattern of rumen fermentation and the composition of the rumen microflora. J. Agric. Sci. 1978, 91, 1. [Google Scholar] [CrossRef]
- Emmanuel, B.; Lawlor, M.J.; McAleese, D. The effect of phosphate and carbonate-bicarbonate supplements on the rumen buffering systems of sheep. Br. J. Nutr. 1970, 24, 653. [Google Scholar] [CrossRef] [PubMed]
- Fernando, S.C.; Purvis, H.T.; Najar, F.Z.; Sukharnikov, L.O.; Krehbiel, C.R.; Nagaraja, T.G.; Roe, B.A.; De Silva, U. Rumen microbial population dynamics during adaptation to a high-grain diet. Appl. Environ. Microbiol. 2010, 76, 7482–7490. [Google Scholar] [CrossRef] [Green Version]
- Khafipour, E.; Krause, D.O.; Plaizier, J.C. A grain-based subacute ruminal acidosis challenge causes translocation of lipopolysaccharide and triggers inflammation. J. Dairy Sci. 2009, 92, 1060–1070. [Google Scholar] [CrossRef] [Green Version]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Global Rumen Census Collaborators; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef]
- Sato, S. Pathophysiological evaluation of subacute ruminal acidosis (SARA) by continuous ruminal pH monitoring. Anim. Sci. J. 2016, 87, 168–177. [Google Scholar] [CrossRef] [Green Version]
- Kaoutari, A.E.; Armougom, F.; Gordon, J.I.; Raoult, D.; Henrissat, B. The abundance and variety of carbohydrate-active enzymes in the human gut microbiota. Nat. Rev. Microbiol. 2013, 11, 497–504. [Google Scholar] [CrossRef]
- Golder, H.M.; Denman, S.E.; McSweeney, C.; Celi, P.; Lean, I.J. Ruminal bacterial community shifts in grain-, sugar-, and histidine-challenged dairy heifers. J. Dairy Sci. 2014, 97, 5131–5150. [Google Scholar] [CrossRef] [PubMed]
- Klieve, A.V.; O’Leary, M.N.; McMillen, L.; Ouwerkerk, D. Ruminococcus bromii, identification and isolation as a dominant community member in the rumen of cattle fed a barley diet. J. Appl. Microbiol. 2007, 103, 2065–2073. [Google Scholar] [CrossRef]
- Zhao, C.; Gao, Z.; Qin, Q.; Ruan, L. Mangroviflexus xiamenensis gen. nov., sp. nov., a member of the family Marinilabiliaceae isolated from mangrove sediment. Int. J. Syst. Evol. Microbiol. 2012, 62, 1819–1824. [Google Scholar] [CrossRef]
- Sakamoto, L.; Benno, B. Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2015; pp. 1–3. [Google Scholar] [CrossRef]
- Morotomi, M.; Nagai, F.; Sakon, H.; Tanaka, R. Dialister succinatiphilus sp. nov. and Barnesiella intestinihominis sp. nov., isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2008, 58, 2716–2720. [Google Scholar] [CrossRef] [Green Version]
- Opdahl, L.J.; Gonda, M.G.; St-Peter, B. Identification of Uncultured Bacterial Species from Firmicutes, Bacteroidetes and CANDIDATUS Saccharibacteria as Candidate Cellulose Utilizers from the Rumen of Beef Cows. Microorganisms 2018, 6, 17. [Google Scholar] [CrossRef] [Green Version]
- Kläring, K.; Hanske, L.; Bui, N.; Charrier, C.; Blaut, M.; Haller, D.; Plugge, C.M.; Clavel, T. Intestinimonas butyriciproducens gen. nov., sp. nov., a butyrate-producing bacterium from the mouse intestine. Int. J. Syst. Evol. Microbiol. 2013, 63, 4606–4612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehority, B.A. Characterization of several bovine rumen bacteria isolated with a xylan medium. J. Bacteriol. 1966, 91, 1724–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hungate, R.E. The Rumen and Its Microbes, 1st ed.; Academic Press: Cambridge, MA, USA, 1966; ISBN 9781483263625. [Google Scholar]
- Downes, J.; Sutcliffe, I.; Tanner, A.C.R.; Wade, W.G. Prevotella marshii sp. nov. and Prevotella baroniae sp. nov., isolated from the human oral cavity. Int. J. Syst. Evol. Microbiol. 2005, 55, 1551–1555. [Google Scholar] [CrossRef] [Green Version]
- McCann, J.C.; Luan, S.; Cardoso, F.C.; Derakhshani, H.; Khafipour, E.; Loor, J.J. Induction of subacute ruminal acidosis affects the ruminal microbiome and epithelium. Front. Microbiol. 2016, 7, 701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khafipour, E.; Li, S.; Plaizier, J.C.; Krause, D.O. Rumen microbiome composition determined using two nutritional models of subacute ruminal acidosis. Appl. Environ. Microbiol. 2009, 75, 7115–7124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, J.M.; D’Antonio, C.M. Elton revisited: A review of evidence linking diversity and invasibility. Oikos 1999, 87, 15–26. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component (%) | RB-1 | RB-3 |
|---|---|---|
| Calcium | 10.89 | 10.61 |
| Phosphorus | 0.06 | 0.05 |
| Potassium | 2.99 | 1.85 |
| Magnesium | 15.43 | 17.96 |
| Sodium | 10.50 | 7.93 |
| Iron | 0.16 | 0.19 |
| Manganese | 0.01 | 0.01 |
| Index | Day (d) | Treatment 1 | SEM | p-Value | ||
|---|---|---|---|---|---|---|
| CON | RB-1 | RB-3 | ||||
| OTU | 0 | 292.33 | 196.67 | 312.67 | 39.829 | 0.294 |
| 5 | 294.00 | 285.00 | 327.67 | 36.461 | 0.753 | |
| Chao1 | 0 | 323.19 | 207.02 | 347.94 | 46.736 | 0.235 |
| 5 | 333.84 | 320.23 | 358.41 | 42.045 | 0.843 | |
| Shannon Index | 0 | 4.66 | 3.85 | 4.77 | 0.783 | 0.235 |
| 5 | 4.40 | 5.00 | 4.93 | 0.720 | 0.843 | |
| Inverse Simpson | 0 | 0.89 | 0.77 | 0.88 | 0.070 | 0.466 |
| 5 | 0.84 | 0.90 | 0.89 | 0.067 | 0.778 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos, S.C.; Jeong, C.-D.; Mamuad, L.L.; Kim, S.-H.; Son, A.-R.; Miguel, M.A.; Islam, M.; Cho, Y.-I.; Lee, S.-S. Enhanced Ruminal Fermentation Parameters and Altered Rumen Bacterial Community Composition by Formulated Rumen Buffer Agents Fed to Dairy Cows with a High-Concentrate Diet. Agriculture 2021, 11, 554. https://doi.org/10.3390/agriculture11060554
Ramos SC, Jeong C-D, Mamuad LL, Kim S-H, Son A-R, Miguel MA, Islam M, Cho Y-I, Lee S-S. Enhanced Ruminal Fermentation Parameters and Altered Rumen Bacterial Community Composition by Formulated Rumen Buffer Agents Fed to Dairy Cows with a High-Concentrate Diet. Agriculture. 2021; 11(6):554. https://doi.org/10.3390/agriculture11060554
Chicago/Turabian StyleRamos, Sonny C., Chang-Dae Jeong, Lovelia L. Mamuad, Seon-Ho Kim, A-Rang Son, Michelle A. Miguel, Mahfuzul Islam, Yong-Il Cho, and Sang-Suk Lee. 2021. "Enhanced Ruminal Fermentation Parameters and Altered Rumen Bacterial Community Composition by Formulated Rumen Buffer Agents Fed to Dairy Cows with a High-Concentrate Diet" Agriculture 11, no. 6: 554. https://doi.org/10.3390/agriculture11060554