
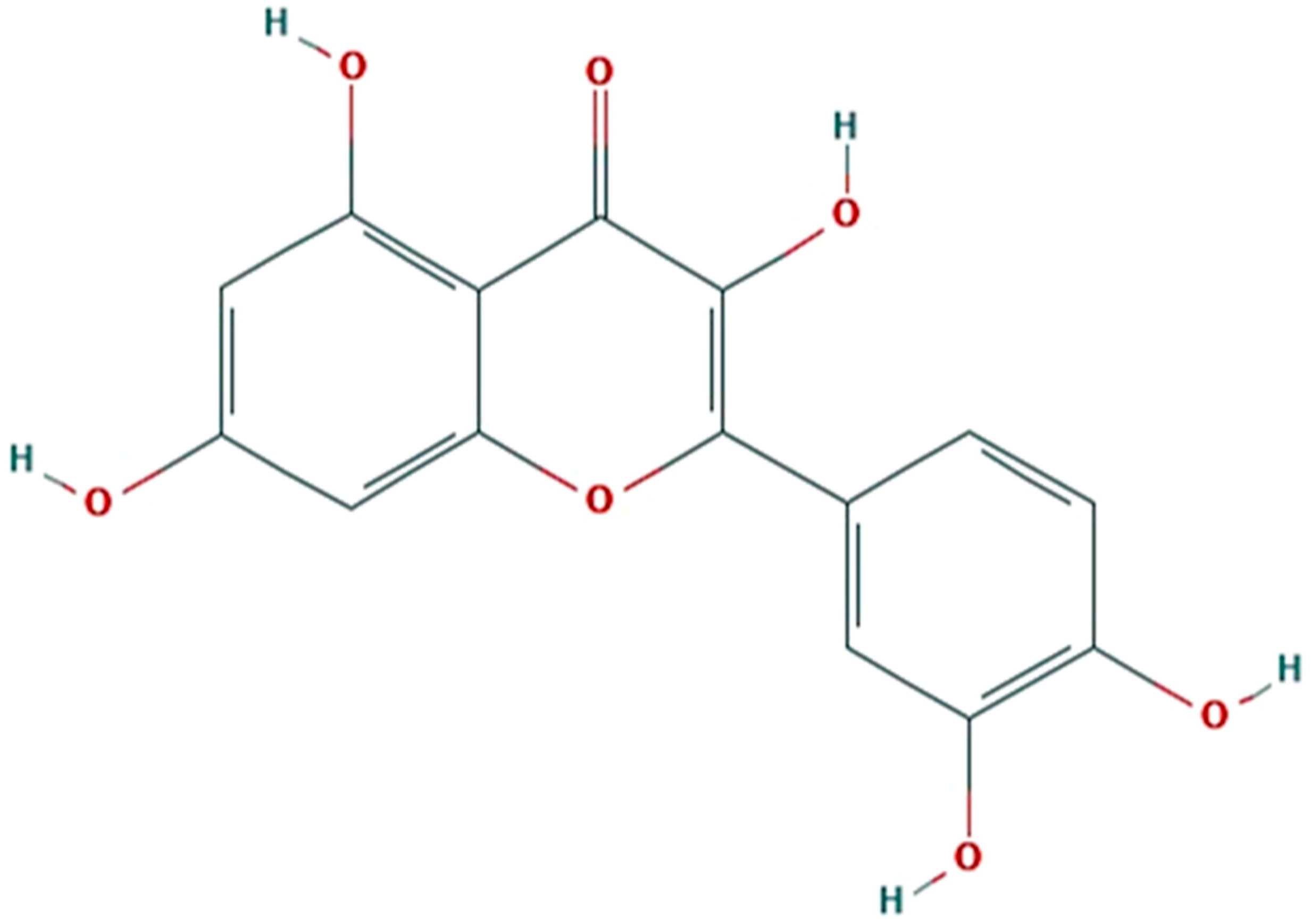
Potential of Quercetin to Reduce Herbivory without Disrupting Natural Enemies and Pollinators


Abstract
:1. Introduction
2. Effects of Quercetin on Herbivores
2.1. Hemiptera (True Bugs)
2.2. Coleoptera (Beetles)
2.3. Lepidoptera (Moths/Butterflies)
2.4. Diptera (True Flies)
2.5. Orthoptera (Grasshoppers)
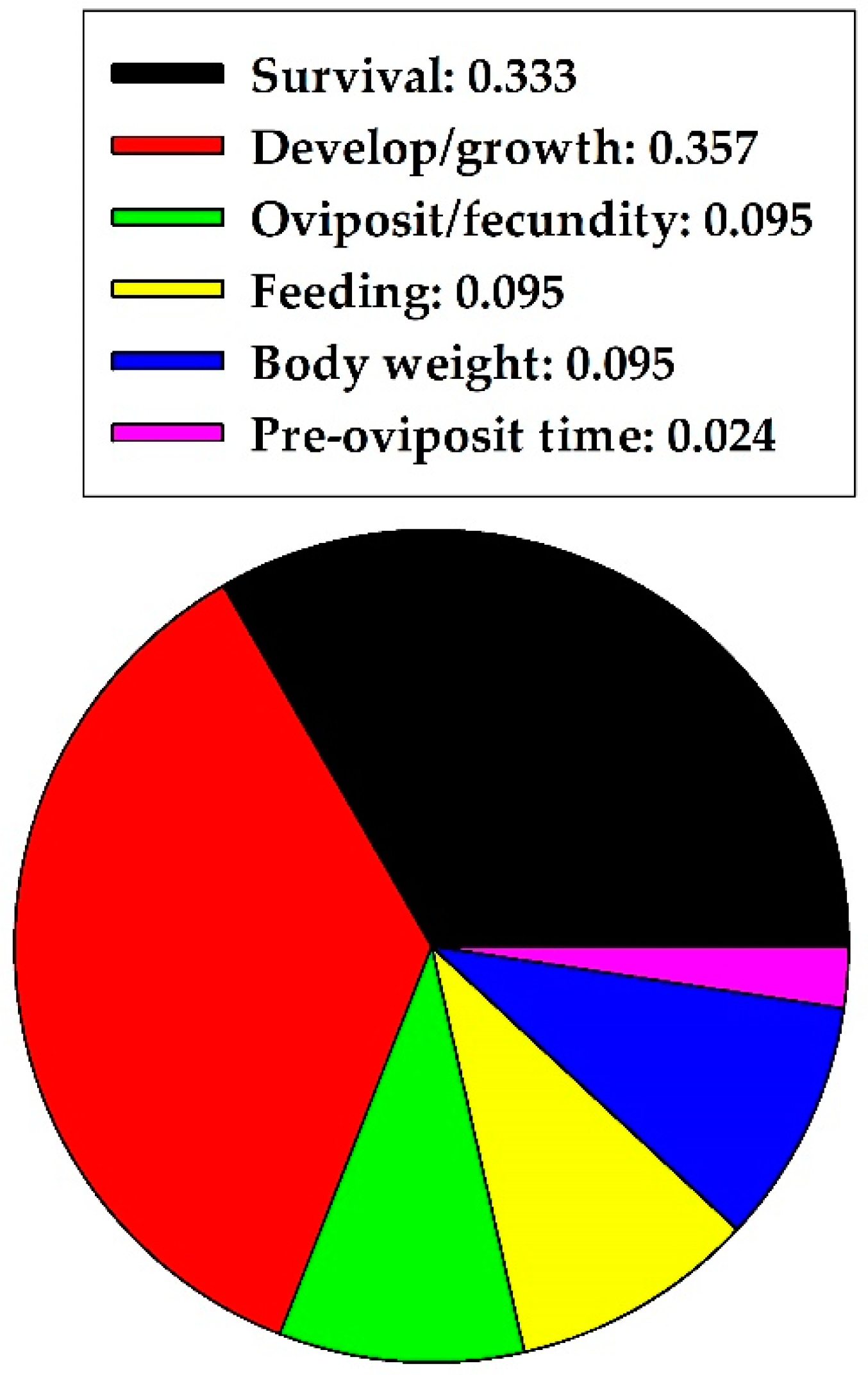
2.6. Summary of Herbivores
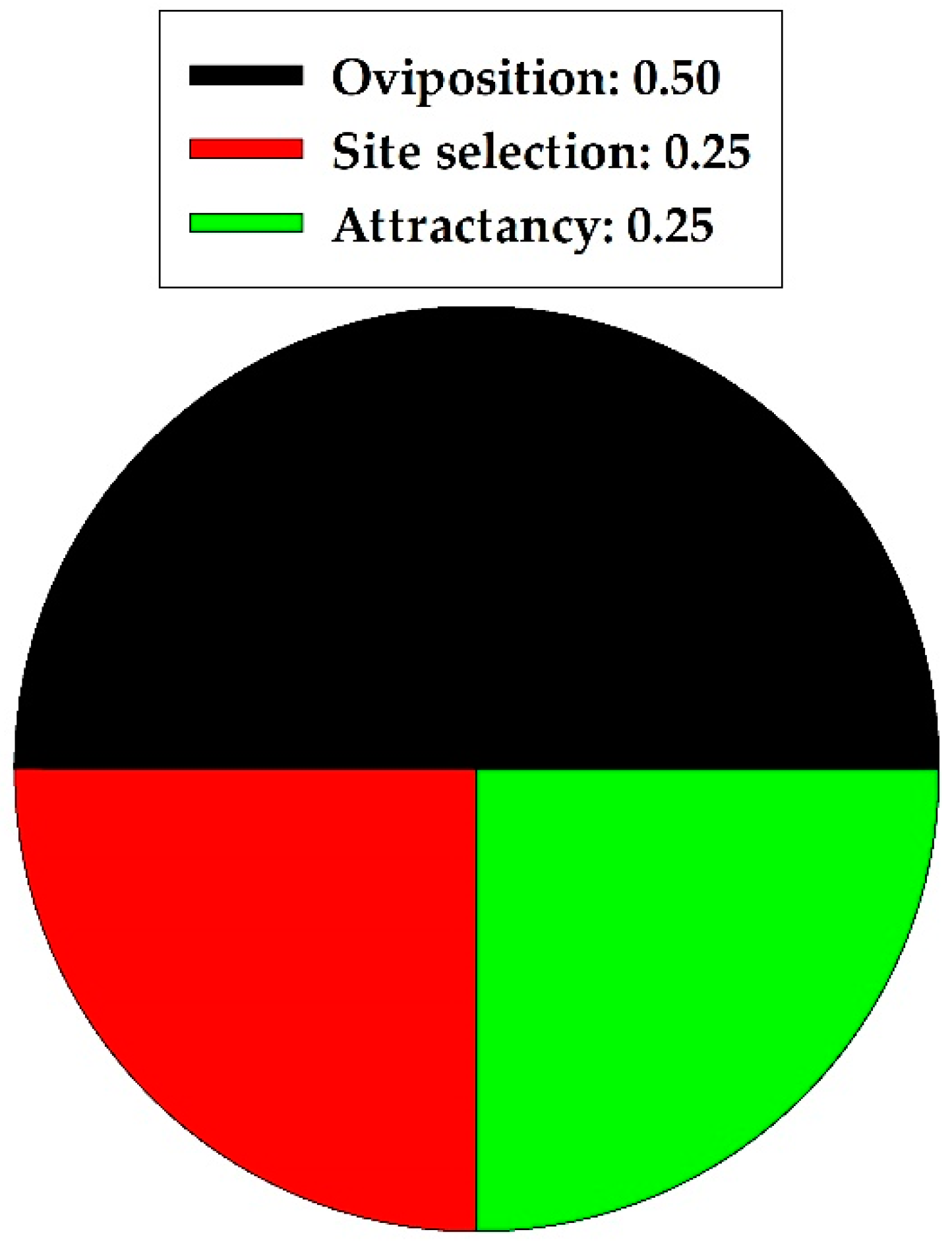
3. Natural Enemies
Predators and Parasitoids
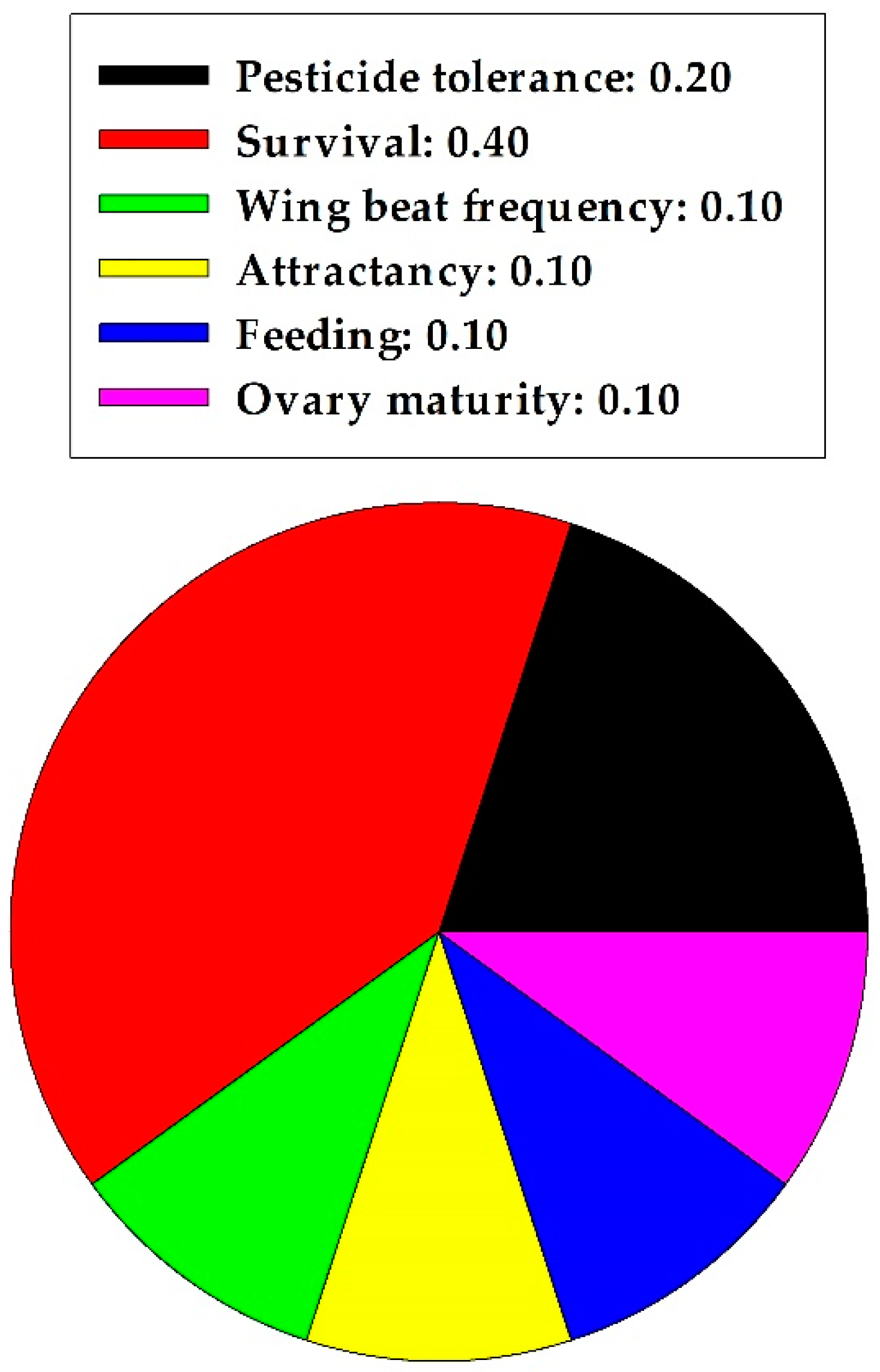
4. Pollinators
Domesticated Honeybee (Hymenoptera)
5. Synthesis and Conclusions
5.1. Synthesis
5.2. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [PubMed]
- Diaz Napal, G.N.; Palacios, S.M. Bioinsecticidal effect of the flavonoids pinocembrin and quercetin against Spodoptera frugiperda. J. Pest Sci. 2015, 88, 629–635. [Google Scholar] [CrossRef]
- Bentivenha, J.P.F.; Canassa, V.F.; Baldin, E.L.L.; Borguini, M.G.; Lima, G.P.P.; Lourenção, A.L. Role of the rutin and genistein flavonoids in soybean resistance to Piezodorus guildinii (Hemiptera: Pentatomidae). Arthropod. Plant Interact. 2018, 12, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Kariyat, R.R.; Gaffoor, I.; Sattar, S.; Dixon, C.W.; Frock, N.; Moen, J.; De Moraes, C.M.; Mescher, M.C.; Thompson, G.A.; Chopra, S. Sorghum 3-deoxyanthocyanidin flavonoids confer resistance against corn leaf aphid. J. Chem. Ecol. 2019, 45, 502–514. [Google Scholar] [CrossRef]
- Harborne, J.B. Biochemistry of plant pollination. In Introduction to Ecological Biochemistry; Academic Press Inc.: London, UK, 1977; Chapter 2; pp. 28–57. [Google Scholar]
- Haribal, M.; Renwick, J.A.A. Oviposition stimulants for the monarch butterfly: Flavonol glycosides from Asclepias curassavica. Phytochemistry 1996, 41, 139–144. [Google Scholar] [CrossRef]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Simmonds, M.S.J. Importance of flavonoids in insect-plant interactions: Feeding and oviposition. Phytochemistry 2001, 56, 245–252. [Google Scholar] [CrossRef]
- Simmonds, M.S.J. Flavonoid-insect interactions: Recent advances in our knowledge. Phytochemistry 2003, 64, 21–30. [Google Scholar] [CrossRef]
- Goławska, S.; Sprawka, I.; Łukasik, I.; Goławski, A. Are naringenin and quercetin useful chemicals in pest-management strategies? J. Pest Sci. 2014, 87, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Perić-Mataruga, V.; Hackenberger, B.K.; Vlahović, M.; Ilijin, L.; Mrdaković, M. Potential improvement of Lymantria dispar L. management by quercetin. Arch. Biol. Sci. 2014, 66, 1125–1129. [Google Scholar] [CrossRef] [Green Version]
- Lundgren, J.G.; Wiedenmann, R.N. Nutritional suitability of corn pollen for the predator Coleomegilla maculata (Coleoptera: Coccinellidae). J. Ins. Physiol. 2004, 50, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Özcan, M.M.; Aljuhaimi, F.; Babiker, E.E.; Uslu, N.; Ceylan, D.A.; Ghafoor, K.; Özcan, M.M.; Dursun, N.; Ahmed, I.M.; Jamiu, F.G.; et al. Determination of antioxidant activity, phenolic compound, mineral contents and fatty acid compositions of bee pollen grains collected from different locations. J. Apicul. Sci. 2019, 63, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Liu, M.; Wang, K.; Yang, Y.; Su, N.; Huang, W.; Wu, Y. Chemical and cytological evaluation of honeybee pollen antioxidant ability. J. Food Sci. 2020, 85, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Diaz Napal, G.N.; Defagó, M.T.; Valladares, G.R.; Palacios, S.M. Response of Epilachna paenulata to two flavonoids, pinocembrin and quercetin, in a comparative study. J. Chem. Ecol. 2010, 36, 898–904. [Google Scholar] [CrossRef]
- Riddick, E.W.; Wu, Z.; Eller, F.J.; Berhow, M.A. Do bioflavonoids in Juniperus virginiana heartwood stimulate oviposition in the ladybird Coleomegilla maculata? Int. J. Insect. Sci. 2018, 10, 1–13. [Google Scholar] [CrossRef]
- Riddick, E.W.; Wu, Z.; Eller, F.J.; Berhow, M.A. Utilization of quercetin as an oviposition stimulant by lab-cultured Coleomegilla maculata in the presence of conspecifics and a tissue substrate. Insects 2018, 9, 77. [Google Scholar] [CrossRef] [Green Version]
- Riddick, E.W. Volatile and non-volatile organic compounds stimulate oviposition by aphidophagous predators. Insects 2020, 11, 683. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.Y.; Huang, X.B.; Li, S.; Hao, K.; Chang, B.H.; Tu, X.B.; Pang, B.P.; Zhang, Z.H. Quercetin affects the growth and development of the grasshopper Oedaleus asiaticus (Orthoptera: Acrididae). J. Econ. Entomol. 2019, 112, 1175–1182. [Google Scholar] [CrossRef]
- Zahedi, A.; Razmjou, J.; Raflee-Dastjerdi, H.; Leppla, N.C.; Golizadeh, A.; Hassanpour, M.; Ebadollahi, A. Tritrophic interactions of cucumber cultivar, Aphis gossypii (Hemiptera: Aphididae), and its predator Hippodamia variegata (Coleoptera: Coccinellidae). J. Econ. Entomol. 2019, 112, 1774–1779. [Google Scholar] [CrossRef]
- Gao, J.; Zhao, G.Y.; Yu, Y.S.; Liu, F.L. High concentration of nectar quercetin enhances worker resistance to queen’s signals in bees. J. Chem. Ecol. 2010, 36, 1241–1243. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.; Wu, W.; Berenbaum, M.R. Impacts of dietary phytochemicals in the presence and absence of pesticides on longevity of honey bees (Apis mellifera). Insects 2017, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, L.; Wu, W.; Berenbaum, M.R. Behavioral responses of honey bees (Apis mellifera) to natural and synthetic xenobiotics in food. Sci. Rep. 2017, 7, 15924. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.; Dharma, K.; Kumar, N.R. Insecticidal effects of aqueous extracts of wild pomegranate peel and seed (Punica granatum L.) against rose aphids, Macrosiphum rosaeformis. J. Appl. Nat. Sci. 2017, 9, 1397–1405. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-Q.; Guo, X.-R.; Li, K.-B.; Yin, J.; Cao, Y.-Z. Resistance of wheat varieties (lines) to Sitobion miscanthi (Takahashi) (Aphidoidea: Aphididae). Acta Entomol. Sin. 2006, 49, 963–968. [Google Scholar]
- Roda, A.L.; Oldham, N.J.; Svatos, A.; Baldwin, I.T. Allometric analysis of the induced flavonols on the leaf surface of wild tobacco (Nicotiana attenuata). Phytochemistry 2003, 62, 527–536. [Google Scholar] [CrossRef]
- Salunke, B.K.; Kotkar, H.M.; Mendki, P.S.; Upasani, S.M.; Maheshwari, V.L. Efficacy of flavonoids in controlling Callosobruchus chinensis (L.) (Coleoptera: Bruchidae), a post-harvest pest of grain legumes. Crop. Prot. 2005, 24, 888–893. [Google Scholar] [CrossRef]
- Matsumoto, H.; Tebayashi, S.; Kuwahara, Y.; Matsuyama, S.; Suzuki, T.; Fujii, K. Identification of taxifolin present in the Azuki bean as an oviposition stimulant of the Azuki bean weevil. J. Pest Sci. 1994, 19, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Adeyemi, M.M.; Agbaji, A.S.; Adebote, D.A.; Amupitan, J.O.; Oyewale, A.O. Antifeedant activity of quercetin isolated from the stem bark of Bobgunnia madagascariensis (Desv.) J.H.Kirkbr & Wiersema (Caesalpiniaceae). Aust. J. Basic. Appl. Sci. 2010, 4, 3342–3346. [Google Scholar]
- Skrzecz, I.; Sowińska, A.; Janiszewski, W. Effects of botanical antifeedants on Melolontha melolontha grub feeding on Scots pine roots. Folia For. Pol. Series A 2014, 56, 135–140. [Google Scholar] [CrossRef]
- Fulcher, A.F.; Ranney, T.G.; Burton, J.D.; Walgenbach, J.F.; Danehower, D.A. Role of foliar phenolics in host plant resistance of Malus taxa to adult Japanese beetles. Hortscience 1998, 33, 862–865. [Google Scholar] [CrossRef] [Green Version]
- Patton, C.A.; Ranney, T.G.; Burton, J.D.; Walgenbach, J.F. Feeding responses of Japanese beetle to naturally occurring metabolites found in rosaceous plants. J. Environ. Hort. 1997, 15, 222–227. [Google Scholar] [CrossRef]
- Dowd, P.F. Responses of Carpophilus hemipterus larvae and adults to selected secondary metabolites of maize. Entomol. Exp. Appl. 1990, 54, 29–36. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, Z.; Cheng, X.; Liu, S.; Wei, Q.; Scott, I.M. Conifer flavonoid compounds inhibit detoxification enzymes and synergize insecticides. Pest Biochem. Physiol. 2016, 127, 1–7. [Google Scholar] [CrossRef]
- Matsuda, K. Feeding stimulation of flavonoids for various leaf beetles (Coleoptera: Chrysomelidae). Appl. Entomol. Zool. 1978, 13, 228–230. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, F.G.; Jenkins, J.N.; Parrott, W.L. Influence of constituents of the cotton plant on feeding, oviposition, and development of the boll weevil. J. Econ. Entomol. 1967, 60, 1294–1297. [Google Scholar] [CrossRef]
- Hedin, P.A.; Miles, L.R.; Thompson, A.C.; Minyard, J.P. Constituents of a cotton bud formulation of a boll weevil feeding stimulant mixture. J. Agric. Food. Chem. 1968, 16, 505–513. [Google Scholar] [CrossRef]
- Chen, C.; Yan, W.; Wang, S.; Shi, X.; Gao, X.; Han, P.; Zhou, X.; Desneux, N. Uptake of quercetin reduces larval sensitivity to lambda-cyhalothrin in Helicoverpa armigera. J. Pest Sci. 2018, 91, 919–926. [Google Scholar] [CrossRef]
- Selin-Rani, S.; Senthil-Nathan, S.; Thanigaivel, A.; Vasantha-Srinivasan, P.; Edwin, E.-S.; Ponsankar, A.; Lija-Escaline, J.; Kalaivani, K.; Abdel-Megeed, A.; Hunter, W.B.; et al. Toxicity and physiological effect of quercetin on generalist herbivore, Spodoptera litura Fab. and a non-target earthworm Eisenia fetida Savigny. Chemosphere 2016, 165, 257–267. [Google Scholar] [CrossRef]
- Li, Z.; Guan, X.; Zhang, Q.; Liu, X.; Michaud, J.P. Quercetin interacts with Cry1Ac protein to affect larval growth and survival of Helicoverpa armigera. Pest Manag. Sci. 2016, 72, 1359–1365. [Google Scholar] [CrossRef]
- Liu, D.; Yuan, Y.; Li, M.; Qiu, X. Effects of dietary quercetin on performance and cytochrome P450 expression of the cotton bollworm, Helicoverpa armigera. Bull Entomol. Res. 2015, 105, 771–777. [Google Scholar] [CrossRef]
- Chacón-Fuentes, M.; Parra, L.; Rodriguez-Saona, C.; Seguel, I.; Ceballos, R.; Quiroz, A. Domestication in Murtilla (Ugni molinae) reduced defensive flavonol levels but increased resistance against a native herbivorous insect. Environ. Entomol. 2015, 44, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-E.; Ma, H.-J.; Feng, D.-D.; Lai, X.-F.; Chen, Z.-M.; Xu, M.-Y.; Yu, Q.-Y.; Zhang, Z. Induction of detoxification enzymes by quercetin in the silkworm. J. Econ. Entomol. 2012, 105, 1024–1042. [Google Scholar] [CrossRef] [PubMed]
- Abou-Zaid, M.M.; Beninger, C.W.; Arnason, J.T.; Nozzolillo, C. The effect of one flavone, two catechins and four flavonols on mortality and growth of the European corn borer (Ostrinia nubilalis Hubner). Biochem. Syst. Ecol. 1993, 21, 415–420. [Google Scholar] [CrossRef]
- Gould, F. Stress specificity of maternal effects in Heliothis virescens (Boddie) (Lepidoptera, Noctuidae) larvae. Mem. Entomol. Soc. Canada 1988, 146, 191–197. [Google Scholar] [CrossRef]
- Shaver, T.N.; Lukefahr, M.J.; Garcia, J. Food utilisation, ingestion, and growth of larvae of the bollworm and tobacco budworm on diets containing gossypol. J. Econ. Entomol. 1970, 63, 1544–1546. [Google Scholar] [CrossRef]
- Shaver, T.N.; Lukefahr, M.J. Effect of flavonoid pigments and gossypol on growth and development of the bollworm, tobacco budworm, and pink bollworm. J. Econ. Entomol. 1969, 62, 643–646. [Google Scholar] [CrossRef]
- Lukefahr, M.J.; Martin, D.F. Cotton-plant pigments as a source of resistance to the bollworm and tobacco budworm. J. Econ. Entomol. 1966, 59, 176–179. [Google Scholar] [CrossRef]
- Sharma, R.; Sohal, S.K. Oviposition response of melon fruit fly, Bactrocera cucurbitae (Coquillett) to different phenolic compounds. J. Biopest. 2016, 9, 46–51. [Google Scholar]
- Sharma, R.; Sohal, S.K. Bioefficacy of quercetin against melon fruit fly. Bull Insectol. 2013, 66, 79–83. [Google Scholar]
- Pree, D.J. Resistance to development of larvae of the apple maggot in crab apples. J. Econ. Entomol. 1977, 70, 611–614. [Google Scholar] [CrossRef]
- Saric, A.; Kalafatic, M.; Rusak, G.; Kovacevic, G.; Franjevic, D.; Gutzeit, H.O. Postembryonic development of Drosophila melanogaster Meigen, 1830 under the influence of quercetin. Entomol. News 2007, 118, 235–240. [Google Scholar] [CrossRef]
- Schramm, D.D.; Collins, H.E.; Hawley, R.S.; German, J.B. Unaltered meiotic chromosome segregation in Drosophila melanogaster raised on a 5% quercetin diet. Food Chem. Toxicol. 1998, 36, 585–589. [Google Scholar] [CrossRef]
- Xu, B.; Wang, Y.; Liu, X.; Yuan, F.; Su, N.; Chen, Y.; Wu, Y.; Zhang, Q. Effects of CryIAc and secondary metabolites in Bt transgenic cottonseed on Lycoriella pleuroti Yang et Zhang (Diptera: Sciaridae). Environ. Entomol. 2006, 35, 807–810. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, X.; Chang, B.H.; Zhang, Z. Growth performance and enzymatic response of the grasshopper, Calliptamus abbreviatus (Orthoptera: Acrididae), to six plant-derived compounds. J. Insect Sci. 2020, 20, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Westcott, N.D.; Hinks, C.F.; Olfert, O. Dietary effects of secondary plant compounds on nymphs of Melanoplus sanguinipes (Orthoptera: Acrididae). Ann. Entomol. Soc. Am. 1992, 85, 304–309. [Google Scholar] [CrossRef]
- Rani, P.U.; Sambangi, P.; Sandhyarani, K. Impact of plant phenolics as semiochemicals on the performance of Trichogramma chilonis Ishii. J. Insect. Behav. 2017, 30, 16–31. [Google Scholar] [CrossRef]
- Ardalani, H.; Vidkjær, N.H.; Laursen, B.B.; Kryger, P.; Fomsgaard, I.S. Dietary quercetin impacts the concentration of pesticides in honey bees. Chemosphere 2021, 262, 1–8. [Google Scholar] [CrossRef]
- Liao, L.-H.; Pearlstein, D.J.; Wu, W.-Y.; Kelley, A.G.; Montag, W.M.; Hsieh, E.M.; Berenbaum, M.R. Increase in longevity and amelioration of pesticide toxicity by natural levels of dietary phytochemicals in the honey bee, Apis mellifera. PLoS ONE 2020, 15, e0243364. [Google Scholar] [CrossRef]
- Liao, L.-H.; Wu, W.-Y.; Dad, A.; Berenbaum, M.R. Fungicide suppression of flight performance in the honeybee (Apis mellifera) and its amelioration by quercetin. Proc. R. Soc. B 2019, 286, 20192041. [Google Scholar] [CrossRef]
- Wong, M.J.; Liao, L.-H.; Berenbaum, M.R. Biphasic concentration-dependent interaction between imidacloprid and dietary phytochemicals in honey bees (Apis mellifera). PLoS ONE 2018, 13, e0206625. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Schuler, M.A.; Berenbaum, M.R. Disruption of quercetin metabolism by fungicide affects energy production in honey bees (Apis mellifera). Proc. Nat. Acad. Sci. USA 2017, 114, 2538–2543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guseman, A.J.; Miller, K.; Kunkle, G.; Dively, G.P.; Pettis, J.S.; Evans, J.D.; van Engelsdorp, D.; Hawthorne, D.J. Multi-drug resistance transporters and a mechanism-based strategy for assessing risks of pesticide combinations to honey bees. PLoS ONE 2016, 11, e0148242. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.M.; Mao, W.F.; Pollock, H.S.; Niu, G.D.; Schuler, M.A.; Berenbaum, M.R. Ecologically appropriate xenobiotics induce cytochrome P450s in Apis mellifera. PLoS ONE. 2012, 7, e31051. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, S.; Yu, O. Metabolic engineering of flavonoids in plants and microorganisms. Appl. Microbiol. Biotechnol. 2011, 91, 949–956. [Google Scholar] [CrossRef]
- Yoon, J.-A.; Kim, B.-G.; Lee, W.J.; Lim, Y.; Chong, Y.; Ahn, J.-H. Production of a novel quercetin glycoside through metabolic engineering of Escherichia coli. Appl. Environ. Microbiol. 2012, 78, 4256–4262. [Google Scholar] [CrossRef] [Green Version]
- Sheng, H.; Sun, X.; Yan, Y.; Yuan, Q.; Wang, J.; Shen, X. Metabolic engineering of microorganisms for the production of flavonoids. Front. Bioeng. Biotechnol. 2020, 8, 589069. [Google Scholar] [CrossRef]
- Bernklau, E.; Bjostad, L.; Hogeboom, A.; Carlisle, A.; Arathi, H.S. Dietary phytochemicals, honey bee longevity and pathogen tolerance. Insects 2019, 10, 14. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Bioassay Method | 1 Effects on Behavior and Life History | 2 Effective Concn. | Reference |
|---|---|---|---|---|
| Herbivore: Hemiptera; true bugs | ||||
| Macrosiphum rosae, nymphs and adults (Aphididae) | Treated red rose (Rosa) foliage | Survival (−−) | 1 mg/mL | [25] |
| Acyrthosiphon pisum, nymphs and adults (Aphididae) | In artificial diet | Development (−−), Pre-oviposition time (−−), Fecundity (−−), Survival (−−) | 1–10 mg/mL, 0.1–10 mg/mL, 1–10 mg/mL, 0.01–10 mg/mL | [11] |
| Sitobion miscanthi, adults (Aphididae) | Innate resistance in wheat ears in field | Fecundity (−−) | 0.199 mg/mL | [26] |
| Tupiocoris notatus, nymphs (Miridae) | Treated tobacco (Nicotiana) leaves | Attractancy (++) | 0.09 μg | [27] |
| Herbivore: Coleoptera; beetles | ||||
| Callosobruchus chinensis, eggs and adults (Bruchidae) | On filter paper and in plastic jar | Survival (−−), Oviposition (−−) | 5.0 mg/mL, 5.0 mg/mL | [28] |
| C. chinensis, adults | On glass beads | Oviposition (oo) | 0.001–1.0 mg/mL | [29] |
| Tribolium castaneum, adults (Tenebrionidae) | On wheat wafer discs | Feeding (−−) | 2.0 mg/mL | [30] |
| Melolontha melolontha, larvae (Scarabaeidae) | In potted soil, in field | Survival (oo) | 20.0 mg/mL | [31] |
| Popillia japonica, adults (Scarabaeidae) | In artificial diet | Feeding (++) | 30.2 mg/mL | [32] |
| P. japonica, adults | In artificial diet | Feeding (++) | 0.302–3.02 mg/mL | [33] |
| Carpophilus hemipterus, larvae and adults (Nitidulidae) | In artificial diet | Feeding (++) | 0.025 mg/mL | [34] |
| Leptinotarsa decemlineata, larvae (Chrysomelidae) | In artificial diet plus insecticide | Survival (−−) | 0.1 mg/mL | [35] |
| Phaedon brassicae, adults (Chrysomelidae) | Treated filter paper | Feeding (−−) | 3.02 mg/mL | [36] |
| Oulema oryzae, adults (Chrysomelidae) | Treated filter paper | Feeding (−−) | 3.02 mg/mL | [36] |
| Plagiodera versicolora, adults (Chrysomelidae) | Treated filter paper | Feeding (++) | 3.02 mg/mL | [36] |
| Altica oleracea, adults (Chrysomelidae) | Treated filter paper | Feeding (+ +) | 3.02 mg/mL | [36] |
| Altica nipponica, adults | Treated filter paper | Feeding (++) | 3.02 mg/mL | [36] |
| Anthonomus grandis, larvae and adults (Curculionidae) | In artificial diet | Feeding (oo), Oviposition (oo), Body weight (++) | 1–10 mg/mL, 1–10 mg/mL, 6 mg/mL | [37] |
| A. grandis, adults | Treated filter paper | Feeding (++) | 0.5 mg/mL | [38] |
| Epilachna paenulata, larvae (Coccinellidae) | Treated squash (Curcubita) leaves | Feeding (++), Survival (−−) | 0.01 μg/cm2, 10–100 μg/cm2 | [16] |
| E. paenulata, larvae | Treated squash (Cucurbita) leaves | Feeding (oo), Body Weight (oo), Survival (oo) | 0.1–50.0 μg/cm2, 0.1–50.0 μg/cm2, 0.1–50.0 μg/cm2 | [16] |
| Herbivore: Lepidoptera; moths/butterflies | ||||
| Helicoverpa armigera, larvae (Noctuidae) | In artificial diet; leaf-dip toxicity test | Development (−−), Pesticide sensitivity (oo) | 0.1% (w/w), 0.1% (w/w) | [39] |
| Spodoptera litura, larvae (Noctuidae) | Toxicity test | Development (−−), Survival (−−) | 0.005 mg/mL, 0.005 mg/mL | [40] |
| Helicoverpa armigera, larvae (Noctuidae) | In artificial diet | Development (−−), Survival (−−), Pesticide sensitivity (oo) | 16 mg/g, 16 mg/g, 16 mg/g | [41] |
| Helicoverpa armigera, larvae (Noctuidae) | Ingested with liquid solution | Development (−−), Survival (−−) | 3 mg/g, 3 mg/g | [42] |
| Spodoptera frugiperda, larvae (Noctuidae) | Treated foliage (Lettuce) | Feeding (++), Feeding (−−) | 0.01 μg/cm2, 100 μg/cm2 | [3] |
| Chilesia rudis, larvae (Arctiidae) | Treated foliage (cultivated Murtilla) | Feeding (++) | 0.005 mg/mL | [43] |
| Lymantria dispar, larvae (Lymantriidae)(from Quercus forest) | In artificial diet | Survival (−−), Body weight (−−) | 2% (w/w), 2% (w/w) | [12] |
| Bombyx mori, larvae (Bombycidae) | In artificial diet | Body weight/Weight gain (−−) | 0.1% (w/w) | [44] |
| Ostrinia nubilalis, larvae (Pyralidae) | In artificial diet | Development (−−) Survival (−−) | 1 mg/g | [45] |
| Heliothis virescens, larvae (Noctuidae) | In artificial diet | Development (−−) | 0.25% (w/w) | [46] |
| Heliothis virescens, larvae Helicoverpa zea, larvae (Noctuidae) | In artificial diet | Body Weight (−−) Feeding (oo) | 0.10% (w/w) | [47] |
| Pectinophora gossypiella, larvae Heliothis virescens, larvae Helicoverpa zea, larvae (Noctuidae) | In artificial diet | Body Weight (−−) Development (−−) | 0.10% (w/w), P. gossypiella; 0.10% (w/w), H. virescens; 0.20% (w/w), H. zea | [48] |
| Heliothis virescens, larvae Helicoverpa zea, larvae (Noctuidae) | In artificial diet | Development (−−) Survival (−−) | 0.20% (w/w), H. virescens; 0.80% (w/w), H. zea | [49] |
| Herbivore: Diptera; true flies | ||||
| Bactrocera cucurbitae, adults (Tephritidae) | On substrate (pumpkin) | Oviposition (−−) | 0.125 mg/mL | [50] |
| B. cucurbitae, eggs, larvae, and pupae | Dipped in test solution | Development (−−), Development (−−), Development (−−) | 3.125 mg/mL, 0.125 mg/mL, 0.005 mg/mL | [51] |
| Rhagoletis pomonella, larvae (Tephritidae) | In artificial diet | Development (−−) | 1.0 mg/mL | [52] |
| Drosophila melanogaster, larvae (Drosophilidae) | In artificial diet | Development (++) | 1.75% (w/w) | [53] |
| D. melanogaster, adults | In artificial diet | Fecundity (++) | 5% (w/w) | [54] |
| Lycoriella pleuroti, larvae (Sciaridae) | In artificial culture media | Survival (−−) | 0.1–0.3% (w/w) | [55] |
| Herbivore: Orthoptera; grasshoppers | ||||
| Calliptamus abbreviatus, nymphs (Acrididae) | Sprayed on alfalfa foliage, field cages | Development (−−) Survival (−−) | 0.10 mg/mL | [56] |
| Oedaleus asiaticus, nymphs (Acrididae) | Sprayed on natural host plant foliage, field cages | Development (−−) Survival (−−) | 0.10–10 mg/mL | [20] |
| Melanoplus sanguinipes, nymphs (Acrididae) | In artificial diet | Body weight (oo) Survival (oo) | 0.125–4.0% (w/w) | [57] |
| Category | Bioassay Method | 1 Effects on Behavior and Life History | 2 Effective Concn. | Reference |
|---|---|---|---|---|
| Natural Enemy: Coleoptera; predatory beetles | ||||
| Coleomegilla maculata, adults (Coccinellidae) | Pure powder in plastic cages | Oviposition (++), Site selection (++) | 1 mg, 1 mg | [17,18] |
| Natural Enemy: Hymenoptera; parasitic wasps | ||||
| Trichogramma chilonis, adults (Trichogrammatidae) | Olfactometry, artificial plant model; lab and semi-field | Attractancy (++), Oviposition (++) | 0.01 mg, 0.03 mg | [58] |
| Category | Bioassay Method | 1 Effects on Behavior and Life History | 2 Effective Concn. | Reference |
|---|---|---|---|---|
| Pollinator: Hymenoptera; social bees | ||||
| Apis mellifera, adults (Apidae) | In sucrose-based artificial diet | Pesticide tolerance (++) | 10 mg/g | [59] |
| A. mellifera, adults | In sucrose-based artificial diet | Pesticide tolerance (++), Survival (++) | 0.004–0.075 mg/mL | [60] |
| A. mellifera, adults | In sucrose-based diet with fungicide | Restored wing beat frequency (++) | 0.075 mg/mL | [61] |
| A. mellifera, adults | In sucrose-based artificial diet with insecticide | Survival (++), at low insecticide conc. | 0.075 mg/mL | [62] |
| A. mellifera, adults | In sucrose-based artificial diet | Survival (++) | 0.075 mg/mL | [23] |
| A. mellifera, adults | In sugar-based diet, semi-field bioassay | Attractancy (++), Feeding (++) | 0.003–0.151 mg/mL, 0.003–0.151 mg/mL | [24] |
| A. mellifera, adults | In sucrose paste with fungicide | Energy production (−−) | 0.075 mg/mL | [63] |
| A. mellifera, adults | In sucrose-based diet with acaricide | Survival (−−) | 0.302 mg/mL | [64] |
| A. mellifera, adults | In sucrose paste diet with acaricide | Survival (++) | 10 mg/g | [65] |
| A. mellifera, adults | In artificial nectar solution | Ovarion maturation (++) | 0.1 mg/mL | [22] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riddick, E.W. Potential of Quercetin to Reduce Herbivory without Disrupting Natural Enemies and Pollinators. Agriculture 2021, 11, 476. https://doi.org/10.3390/agriculture11060476
Riddick EW. Potential of Quercetin to Reduce Herbivory without Disrupting Natural Enemies and Pollinators. Agriculture. 2021; 11(6):476. https://doi.org/10.3390/agriculture11060476
Chicago/Turabian StyleRiddick, Eric W. 2021. "Potential of Quercetin to Reduce Herbivory without Disrupting Natural Enemies and Pollinators" Agriculture 11, no. 6: 476. https://doi.org/10.3390/agriculture11060476