
Biological Indicators of Soil Condition on the Kabanyolo Experimental Field, Uganda

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
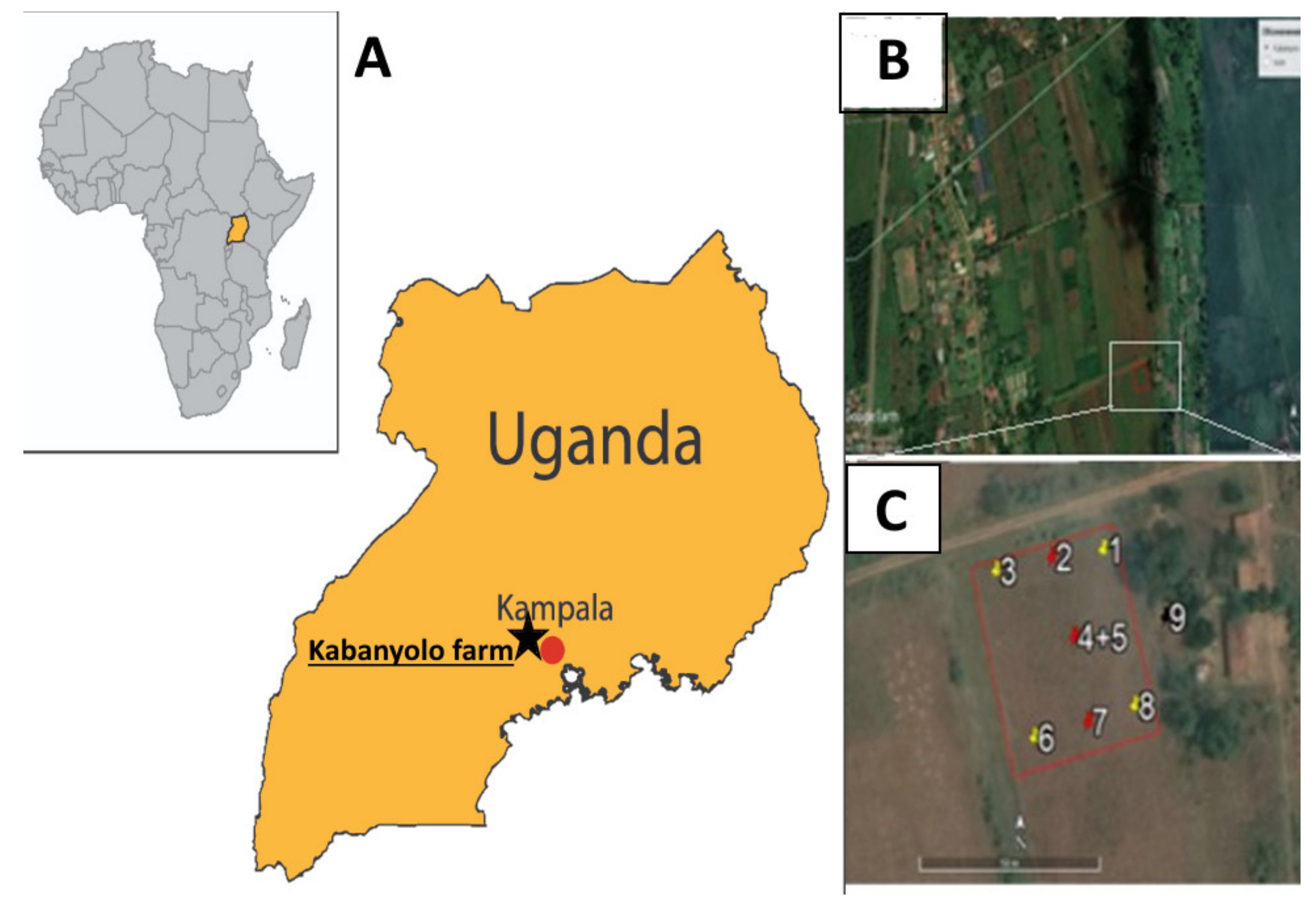
2.1. Study Area
2.2. Sampling
2.3. Soil Chemical Analyses
2.4. Analysis of the Intensity of the Nitrogen and Carbon Cycle Processes
2.4.1. Pre-Preparation of Soil Samples
2.4.2. The Processes of the C Cycle
2.4.3. Processes of the N Cycle
2.4.4. Computation of Additional Parameters of the State of Soil Microbial Communities
2.4.5. Estimation of Prokaryotic Components in the Soil Microbial Complex by Quantitative PCR (qPCR)
2.4.6. Statistical Processing of Results
3. Results
3.1. Soil Properties
3.1.1. Natural Soil Properties and Classification
3.1.2. Cultivated Topsoil Properties
3.2. Indicators of Biological Activity
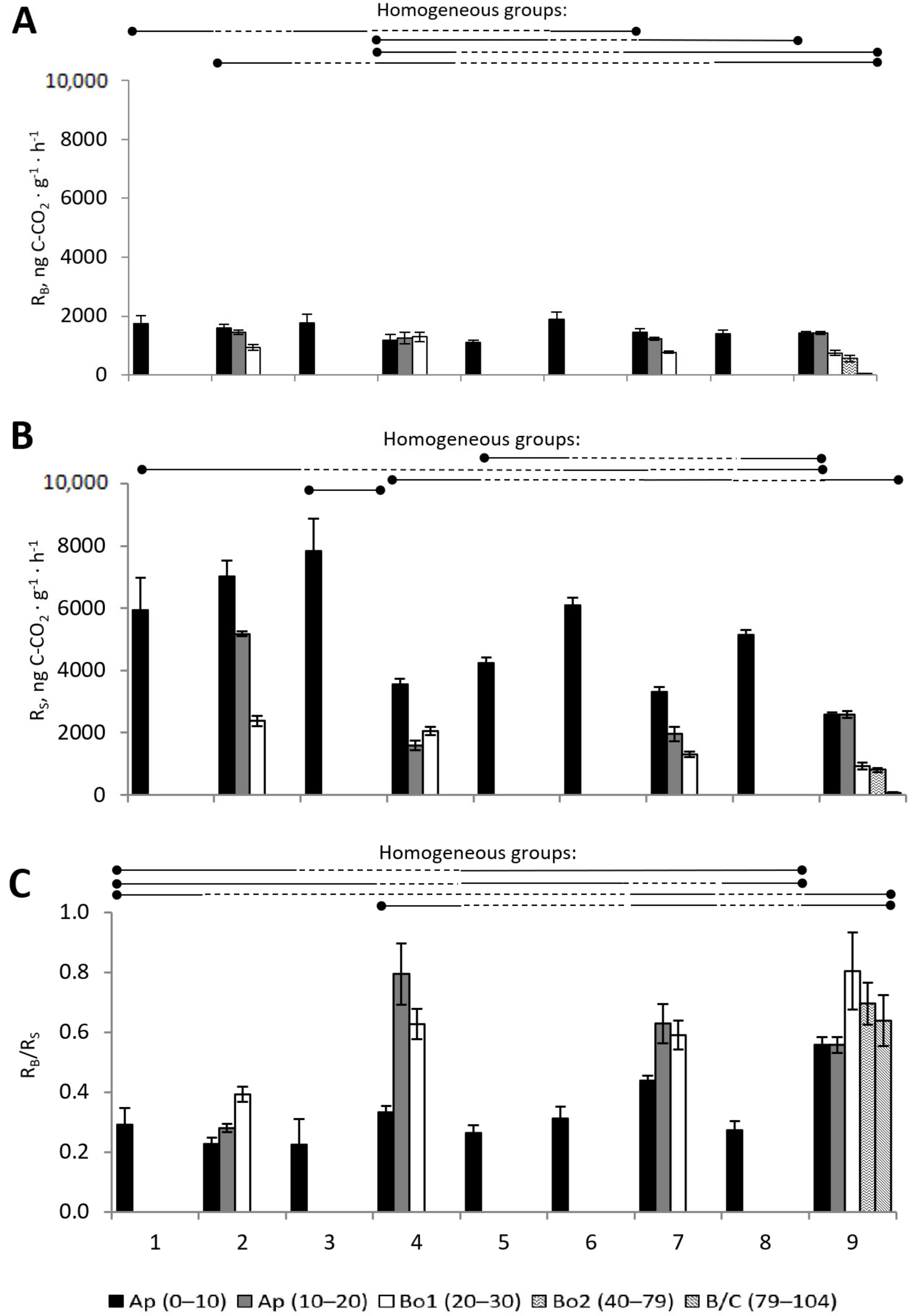
3.2.1. Basal and Substrate-Induced Respiration
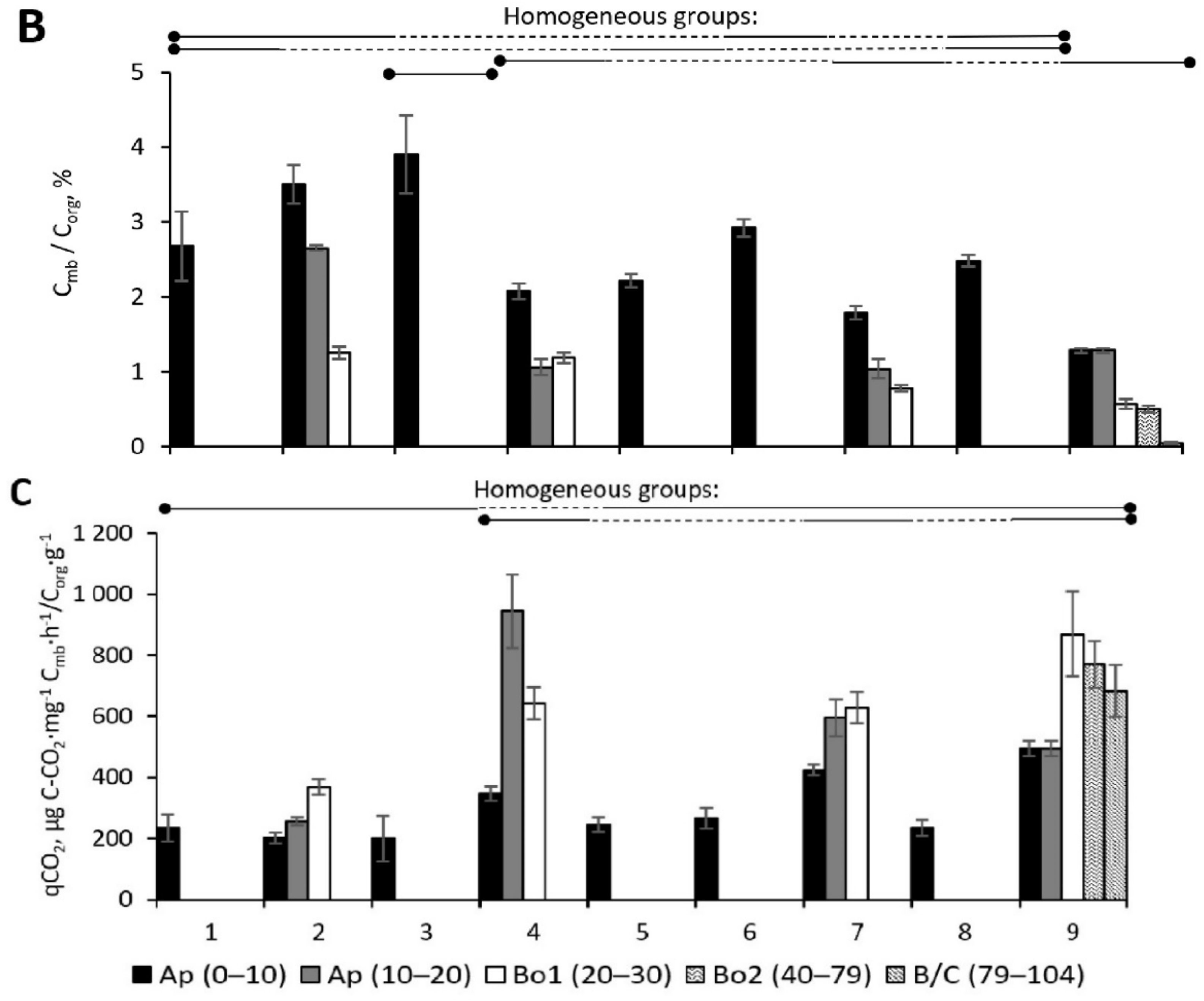
3.2.2. Microbial Biomass C Stock
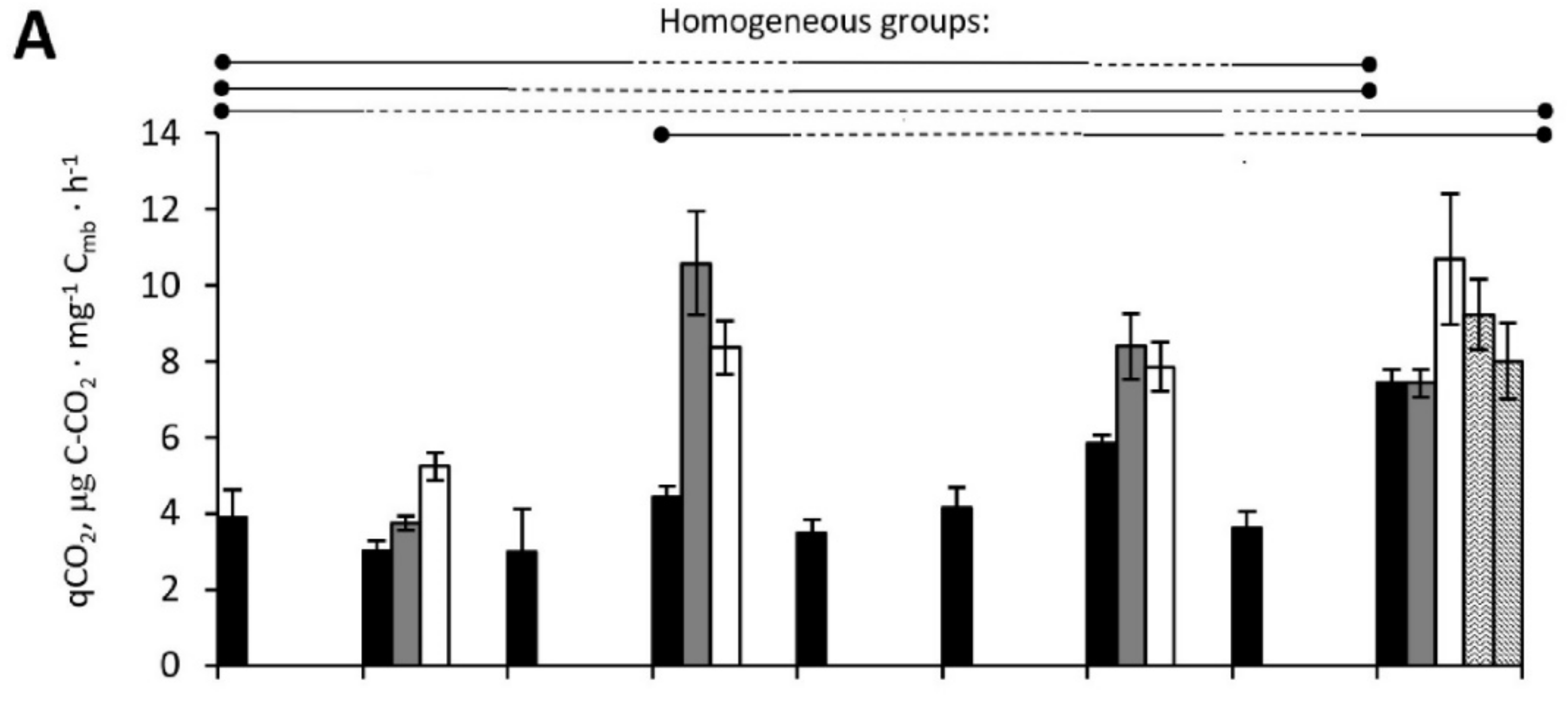
3.2.3. Microbial Metabolic Coefficient qCO2
3.2.4. CH4 Production
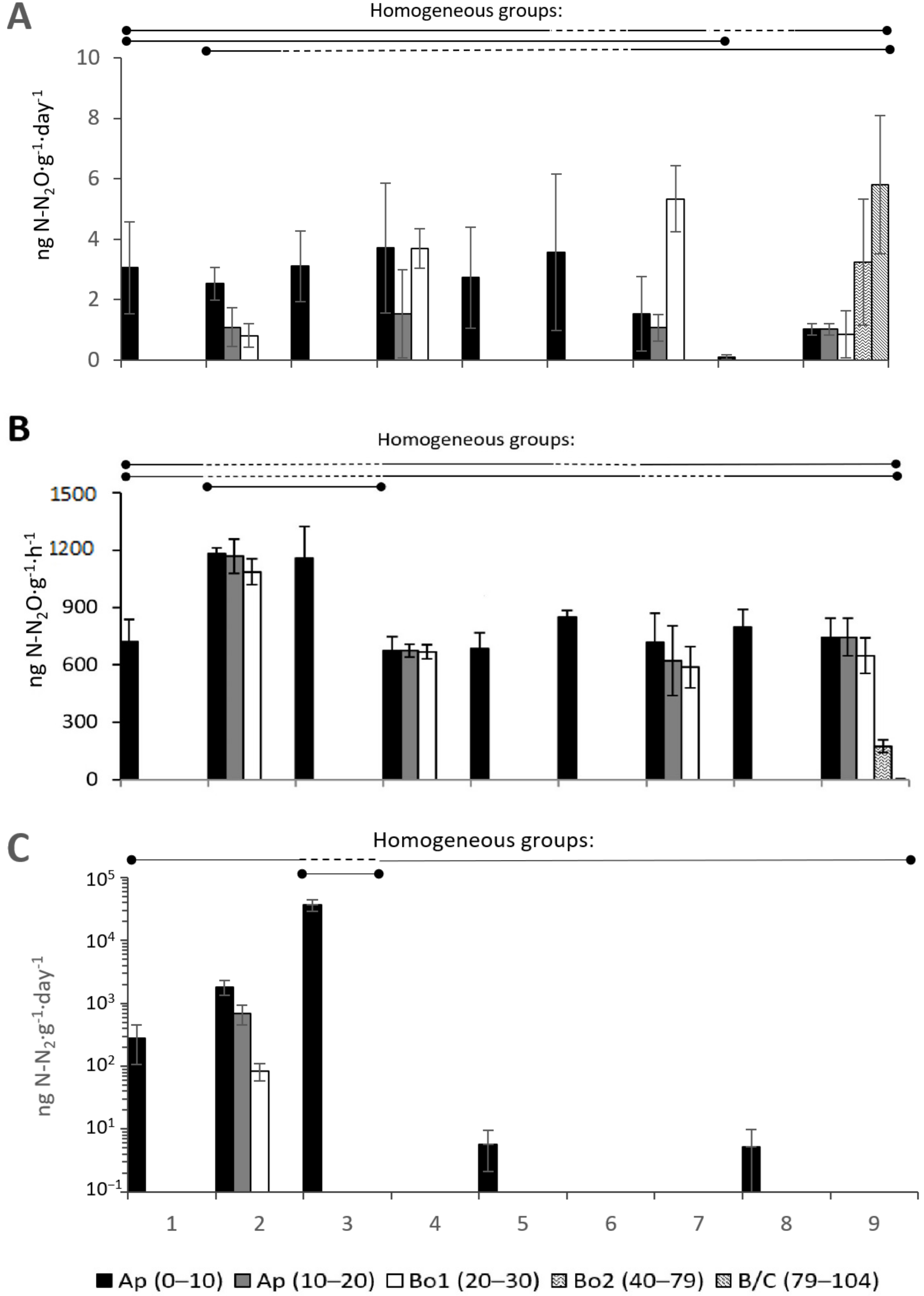
3.2.5. N2O Formation and Nitrogen Fixation Activity
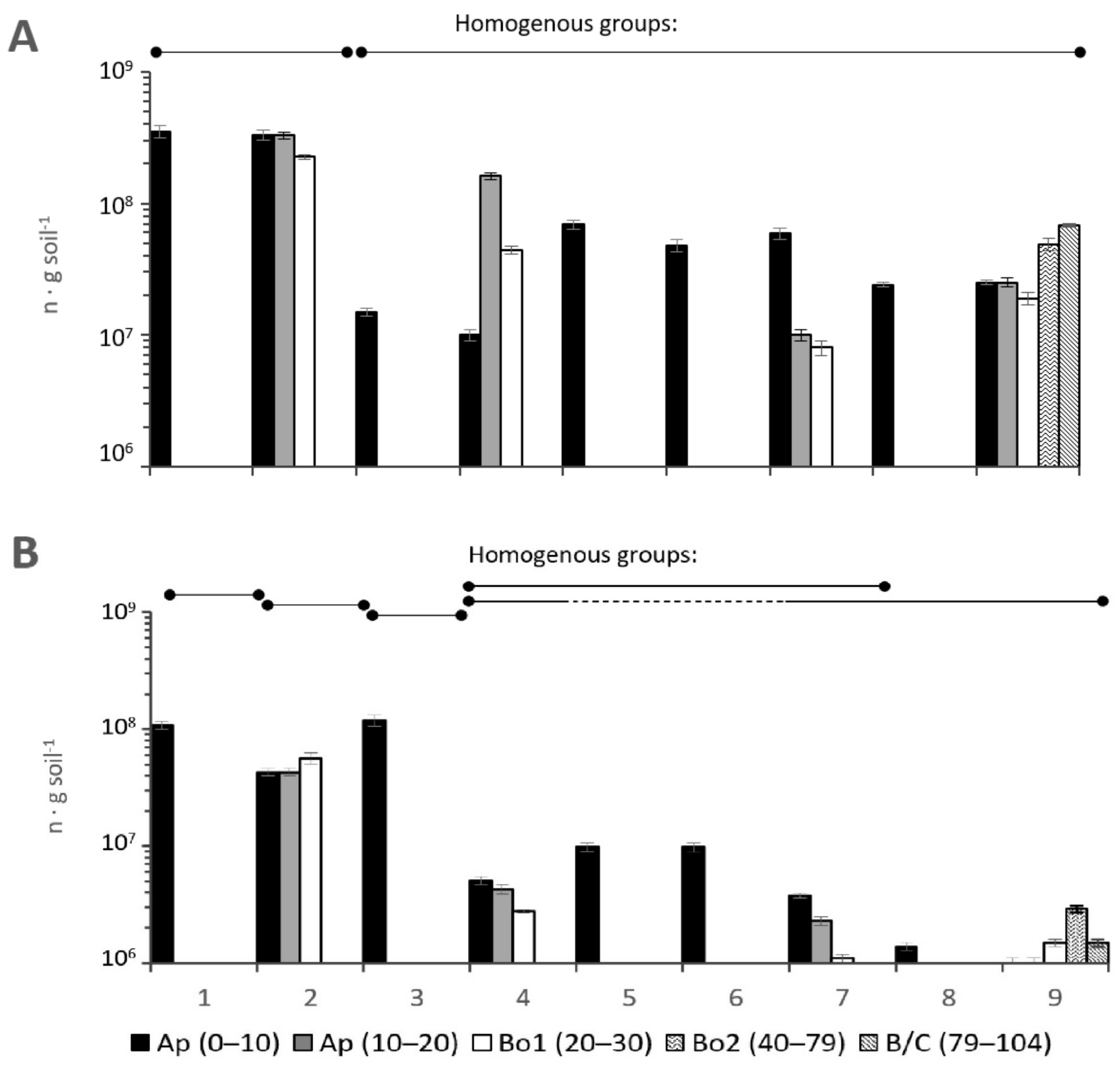
3.2.6. The Number of Copies of Genes of Prokaryotes
3.2.7. ANOVA for Microbial Processes
4. Discussion
4.1. Chemical Parameters of the Habitat of the Soil Microbial Pool
4.2. Participation of the Soil Microbiome in the C Cycle
4.3. Participation of the Soil Microbiome in the N Cycle
4.4. The Number of Copies of Prokaryotic Genes (Bacteria and Archaea)
4.5. Soil Biological Indicators and Fertilisers
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hartemink, A.E. Soil Fertility Decline in the Tropics: With Case Studies on Plantations; CABI Publishing: Wallingford, UK, 2003; p. 360. [Google Scholar]
- Sanchez, P.A. Soil fertility and hunger in Africa. Science 2002, 295, 2019–2020. [Google Scholar] [CrossRef] [Green Version]
- Stewart, Z.P.; Pierzynski, G.M.; Middendorf, B.J.; Prasad, P.V. Approaches to improve soil fertility in sub-Saharan Africa. J. Exp. Bot. 2020, 71, 632–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baah-Ofori, R.N.; Amoakohene, M. A review of soil fertility management communication in sub-Saharan Africa. J. Agric. Rural. Dev. Trop. Subtrop. (JARTS) 2021, 122, 1–12. [Google Scholar] [CrossRef]
- Zhang, Е.; Zhan, J.; Zhang, Q.; Yao, L.; Liu, W. Impacts of land use/cover change on terrestrial carbon stocks in Uganda. Phys. Chem. Earth 2017, 101, 195–203. [Google Scholar] [CrossRef]
- Abawi, G.S.; Widmer, T.L. Impact of soil health management practices on soilborne pathogens, nematodes and root diseases of vegetable crops. Appl. Soil Ecol. 2000, 15, 37–47. [Google Scholar] [CrossRef]
- Kibblewhite, M.G.; Ritz, K.; Swift, M.J. Soil health in agricultural systems. Philos. Trans. R. Soc. B Biol. Sci. 2007, 363, 685–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kihara, J.; Bolo, P.; Kinyua, M.; Nyawira, S.S.; Sommer, R. Soil health and ecosystem services: Lessons from sub-Sahara Africa (SSA). Geoderma 2020, 370, 114342. [Google Scholar] [CrossRef]
- Wade, J.; Culman, S.W.; Hurisso, T.T.; Miller, R.O.; Baker, L.; Horwath, W.R. Sources of variability that compromise smineralisable carbon as a soil health indicator. Soil Sci. Soc. Am. J. 2018, 82, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Van Bruggen, A.H.; Semenov, A.M. In search of biological indicators for soil health and disease suppression. Appl. Soil Ecol. 2000, 15, 13–24. [Google Scholar] [CrossRef]
- Arias, M.E.; González-Pérez, J.A.; González-Vila, F.J.; Ball, A.S. Soil health: A new challenge for microbiologists and chemists. Int. Microbiol. 2005, 8, 13–21. [Google Scholar]
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fertil. Soils 2012, 48, 489–499. [Google Scholar] [CrossRef]
- Singh, B. Are nitrogen sfertilisers deleterious to soil health? Agronomy 2018, 8, 48. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, S.; Srivastava, P.; Devi, R.S.; Bhadouria, R. Influence of synthetic sfertilisers and pesticides on soil health and soil microbiology. In Agrochemicals Detection, Treatment and Remediation; Vara Prasad, M.N., Ed.; Butterworth-Heinemann: Oxford, UK, 2020; pp. 25–54. [Google Scholar] [CrossRef]
- Okalebo, J.R.; Othieno, C.O.; Nekesa, A.O.; Ndungu-Magiroi, K.W.; Kifuko-Koech, M.N. Potential for agricultural lime on improved soil health and agricultural production in Kenya. In Proceedings of the 9th African Crop Science, Cape Town, South Africa, 28 September–2 October 2009; Volume 9, pp. 339–341. [Google Scholar]
- Kaweesa, S.H.; Ndah, H.T.; Schuler, J.; Melcher, A.; Loiskandl, W. Understanding the conditions of conservation agriculture adoption in Lango region, Uganda. Agroecol. Sustain. Food Syst. 2020, 44, 1260–1279. [Google Scholar] [CrossRef]
- Okiror, P.; Lejju, J.B.; Bahati, J.; Rugunda, G.K.; Sebuuwufu, C.I.; Mulindwa, P.; Ocan, J.J. Suitability of Kabanyolo soils for fruit and vegetable production. Open J. Soil Sci. 2017, 7, 19–33. [Google Scholar] [CrossRef] [Green Version]
- Ministry of Lands, Housing and Urban Development, Republic of Uganda. The National Land Use Policy; Ministry of Lands, Housing and Urban Development, Republic of Uganda: Kampala, Uganda, 2013; p. 62.
- Magunda, M.K.; Larson, W.E.; Linden, D.R.; Nater, E.A. Changes in microrelief and their effects on infiltration and erosion during simulated rainfall. Soil Technol. 1997, 10, 57–67. [Google Scholar] [CrossRef]
- Van Reeuwijk, L.P. Procedures for Soil Analysis; ISRIC Technical paper 9; ISRIC: Wageningen, The Netherlands, 2002; p. 90. [Google Scholar]
- Bashour, I.I.; Sayegh, A.H. Methods of Analysis for Soils of Arid and Semi-Arid Region; Food and Agriculture Organization of the United Nations: Rome, Italy, 2007; p. 119. Available online: https://msibsri4313.files.wordpress.com/2013/11/methods-of-soil-analysis-for-arid-semiarid-regions.pdf (accessed on 11 October 2021).
- Anderson, J.P.E.; Domsch, K.H. Application of eco-physiological quotients (qCO2 and qD) on microbial biomasses from soils of different cropping histories. Soil Biol. Biochem. 1990, 22, 251–255. [Google Scholar] [CrossRef]
- Berg, B.; Wessén, B.; Ekbohm, G. Nitrogen level and decomposition in Scots pine needle litter. Oikos 1982, 38, 291–296. [Google Scholar] [CrossRef]
- Anderson, J.P.E.; Domsch, K.H. A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biol. Biochem. 1978, 10, 215–221. [Google Scholar] [CrossRef]
- SsoFast EvaGreen Supermix. Available online: http://www.bio-rad.com/webroot/web/pdf/lsr/literature/10014647A.pdf (accessed on 8 July 2021).
- Yu, Y.; Lee, C.; Hwang, S. Analysis of community structures in anaerobic processes using a quantitative real-time PCR method. Water Sci. Technol. 2005, 52, 85–91. [Google Scholar] [CrossRef]
- Manucharova, N.A.; Ksenofontova, N.A.; Belov, A.A.; Kamenskiy, N.N.; Arzamazova, A.V.; Zenova, G.M.; Kinzhaev, R.R.; Trofimov, S.Y.; Stepanov, A.L. Prokaryotic component of oil-contaminated oligotrophic peat soil under different levels of mineral nutrition: Biomass, diversity, and activity. Euras. Soil Sci. 2021, 54, 89–97. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, J.A.; Vilgalys, R.; Jacksson, R.B. Assessment of soil microbial community structure by use of taxon-specific quantitative PCR assays. Appl. Environ. Microbiol. 2005, 71, 4117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenywa, J.S.; Amoding-Katushabe, A.; Jumba, F. Distribution of nitrate and related nutrient stocks in a fertilised horticultural valley soil in central Uganda. Afr. J. Rural. Dev. 2020, 4, 413–420. Available online: http://afjrd.org/jos/index.php/afjrd/article/view/2118 (accessed on 11 October 2021).
- Pidgeon, J.D. The measurement and prediction of available water capacity of ferrallitic soils in Uganda. J. Soil Sci. 1972, 23, 431–441. [Google Scholar] [CrossRef]
- Banage, W.B.; Visser, S.A. Soil moisture and temperature levels and fluctuations in one year in a Uganda soil catena. East Afr. Agric. For. J. 1967, 32, 450–455. [Google Scholar] [CrossRef]
- Twongyirwe, R.; Sheil, D.; Majaliwa, J.G.M.; Ebanyat, P.; Tenywa, M.M.; van Heist, M.; Kumar, L. Variability of soil organic carbon stocks under different land uses: A study in an afro-montane landscape in southwestern Uganda. Geoderma 2013, 193, 282–289. [Google Scholar] [CrossRef]
- Musinguzi, P.; Tenywa, J.S.; Ebanyat, P.; Basamba, T.A.; Tenywa, M.M.; Mubiru, D.N.; Zinn, Y.L. Soil organic fractions in cultivated and uncultivated Ferralsols in Uganda. Geoderma Reg. 2015, 4, 108–113. [Google Scholar] [CrossRef]
- Tumwebaze, S.B.; Byakagaba, P. Soil organic carbon stocks under coffee agroforestry systems and coffee monoculture in Uganda. Agric. Ecosyst. Environ. 2016, 216, 188–193. [Google Scholar] [CrossRef]
- Musinguzi, P.; Ebanyat, P.; Tenywa, J.S.; Basamba, T.A.; Tenywa, M.M.; Mubiru, D.N. Critical soil organic carbon range for optimal crop response to mineral fertiliser nitrogen on a Ferralsol. Exp. Agric. 2016, 52, 635–653. [Google Scholar] [CrossRef]
- Mallarino, A.P.; Sawyer, J.E.; Barnhart, S.K. A General Guide for Crop Nutrient and Limestone Recommendations in Iowa; Extension and Outreach Publications 82; Iowa State University, University Extension: Ames, IA, USA, 2013; pp. 1–12. Available online: https://lib.dr.iastate.edu/extension_pubs/82 (accessed on 11 October 2021).
- Hall, N.M.; Kaya, B.; Dick, J.; Skiba, U.; Niang, A.; Tabo, R. Effect of improved fallow on crop productivity, soil fertility and climate-forcing gas emissions in semi-arid conditions. Biol. Fertil. Soils 2006, 42, 224–230. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.C.; Singh, J.S. Effect of cultivation on microbial carbon and nitrogen in dry tropical forests soil. Biol. Fertil. Soils 1989, 8, 343–348. [Google Scholar] [CrossRef]
- Gyapong, K.A.B.; Ayisi, C.L. The effect of organic manures on soil fertility and microbial biomass carbon, nitrogen and phosphorus under maise-cowpea intercropping system. Discourse J. Agric. Food Sci. 2015, 3, 65–77. [Google Scholar]
- Bastida, F.; Zsolnay, A.; Hernández, T.; García, C. Past, present and future of soil quality indices: A biological perspective. Geoderma 2008, 147, 159–171. [Google Scholar] [CrossRef]
- Susyan, E.A.; Wirth, S.; Ananyeva, N.D.; Stolnikova, E.V. Forest succession on abandoned arable soils in European Russia e Impacts on microbial biomass, fungal-bacterial ratio, and basal CO2 respiration activity. Eur. J. Soil Biol. 2011, 47, 169–174. [Google Scholar] [CrossRef]
- Saviozzi, A.; Levi-Minzi, R.; Cardelli, R.; Riffaldi, R. A comparison of soil quality in adjacent cultivated, forest and native grassland soils. Plant Soil 2001, 233, 251–259. [Google Scholar] [CrossRef]
- Ohigashi, T.; Schloter, M.; Schulz, S.; Munthali, K.; Uchida, Y. Changes in soil prokaryotic diversity in response to land-use changes in Sub-Saharan Africa. Soil Syst. 2021, 5, 62. [Google Scholar] [CrossRef]
- Zhelezova, A.D.; Tkhakakhova, A.K.; Yaroslavtseva, N.V.; Garbuz, S.A.; Lazarev, V.I.; Kogut, B.M.; Kutovaya, O.V.; Kholodov, V.A. Microbiological parameters of aggregates in typical chernozems of long-term field experiments. Euras. Soil Sci. 2017, 50, 701–707. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Group of Organisms/Gene | Primers 1—Forward; 2—Reverse | The Nucleotide Sequence of the Probe (5′-3′3′) |
|---|---|---|
| Archaea | arc915f arc1059r | AGGAATTGGCGGGGGAGCAC GCCATGCACCWCCTCT |
| Bacteria | Eub338 Eub518 | ACT CCT ACG GGA GGC AGCAG ATT ACC GCG GCT GCT GG |
| Horizon, Depth, cm | OC | pHH20 | pHKCl | P2O5 | K2O | Ca2+ | Mg2+ | K+ | Na+ | CEC | BS | Fed |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| g kg−1 | mg kg−1 | cmolc kg−1 | % | |||||||||
| Soil profile outside the field | ||||||||||||
| Ap (0–20) | 15.0 | 5.51 | 4.71 | 58.91 | 109.9 | 4.75 | 1.52 | 0.18 | 0.002 | 8.34 | 77.3 | 2.60 |
| AB (20–40) | 12.3 | 5.46 | 5.12 | 16.60 | 73.2 | 2.79 | 1.16 | 0.05 | 0.011 | 5.00 | 80.1 | 3.12 |
| Bo1 (40–79) | 12.0 | 5.11 | 4.93 | 10.50 | 63.8 | 2.23 | 1.12 | 0.04 | 0.005 | 4.64 | 73.0 | 3.42 |
| Bo2 (79–104) | 11.7 | 4.97 | 4.86 | 13.40 | 38.1 | 1.51 | 0.99 | 0.00 | 0.005 | 3.77 | 66.6 | 3.38 |
| Surficial horizon of the soil on the experimental field (mean values, n = 11) | ||||||||||||
| Ap (0–20) | 14.4 ± 0.9 | 5.52 ± 0.20 | 4.84 ± 0.26 | 54.11 ± 43.98 | 128.27 ± 58.13 | 5.10 ± 1.17 | 1.48 ± 0.20 | 0.26 ± 0.18 | 0.005 ± 0.003 | 8.09 ± 0.60 | 82.74 ± 13.34 | - |
| AB (20–30) | 13.2 ± 1.0 | 5.30 ± 0.61 | 4.97 ± 0.38 | 46.73 ± 42.96 | 94.30 ± 76.69 | 4.38 ± 3.03 | 1.37 ± 0.55 | 0.14 ± 0.13 | 0.010 ± 0.001 | 6.89 ± 2.81 | 81.63 ± 20.82 | - |
| Horizon | Ap | Ap | AB | Bo | BC |
|---|---|---|---|---|---|
| Depth, cm | 0–10 | 10–20 | 20–30 | 40–79 | 79–104 |
| Mean for the experimental field | 403.7 ± 121.6 | 217.8 ± 128.3 | 143.6 ± 36.7 | ||
| Reference profile | 193.2 ± 4.9 | 70.0 ± 8.4 | 60.7 ± 4.7 | 5.81 ± 1.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanova, A.; Denisova, E.; Musinguzi, P.; Opolot, E.; Tumuhairwe, J.B.; Pozdnyakov, L.; Manucharova, N.; Ilichev, I.; Stepanov, A.; Krasilnikov, P. Biological Indicators of Soil Condition on the Kabanyolo Experimental Field, Uganda. Agriculture 2021, 11, 1228. https://doi.org/10.3390/agriculture11121228
Ivanova A, Denisova E, Musinguzi P, Opolot E, Tumuhairwe JB, Pozdnyakov L, Manucharova N, Ilichev I, Stepanov A, Krasilnikov P. Biological Indicators of Soil Condition on the Kabanyolo Experimental Field, Uganda. Agriculture. 2021; 11(12):1228. https://doi.org/10.3390/agriculture11121228
Chicago/Turabian StyleIvanova, Anna, Elizaveta Denisova, Patrick Musinguzi, Emmanuel Opolot, John Baptist Tumuhairwe, Lev Pozdnyakov, Natalia Manucharova, Igor Ilichev, Aleksey Stepanov, and Pavel Krasilnikov. 2021. "Biological Indicators of Soil Condition on the Kabanyolo Experimental Field, Uganda" Agriculture 11, no. 12: 1228. https://doi.org/10.3390/agriculture11121228