
Isolation and Identification Rust Pathogens and the Study of Antioxidant Enzyme Activity and Gene Expression under Rust Infection in Zoysia japonica


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Zoysiagrass Rust Pathogen
2.2. Infection of Zoysiagrass Rust Pathogen in Different Germplasm of Z. japonica
2.3. Antioxidant Enzyme Activity and Gene Expression Analyses
2.4. Statistical Analysis
3. Results
3.1. Morphological Identification of Zoysiagrass Rust Pathogen
3.2. Sequence Homology Analysis of Zoysiagrass Rust Pathogen 18S rDNA
3.3. The Disease Severity and Infection Type Survey
3.4. Morphological Observations of Z. japonica Leaves Inoculated with Rust Pathogen
3.5. Antioxidant Enzyme Activity
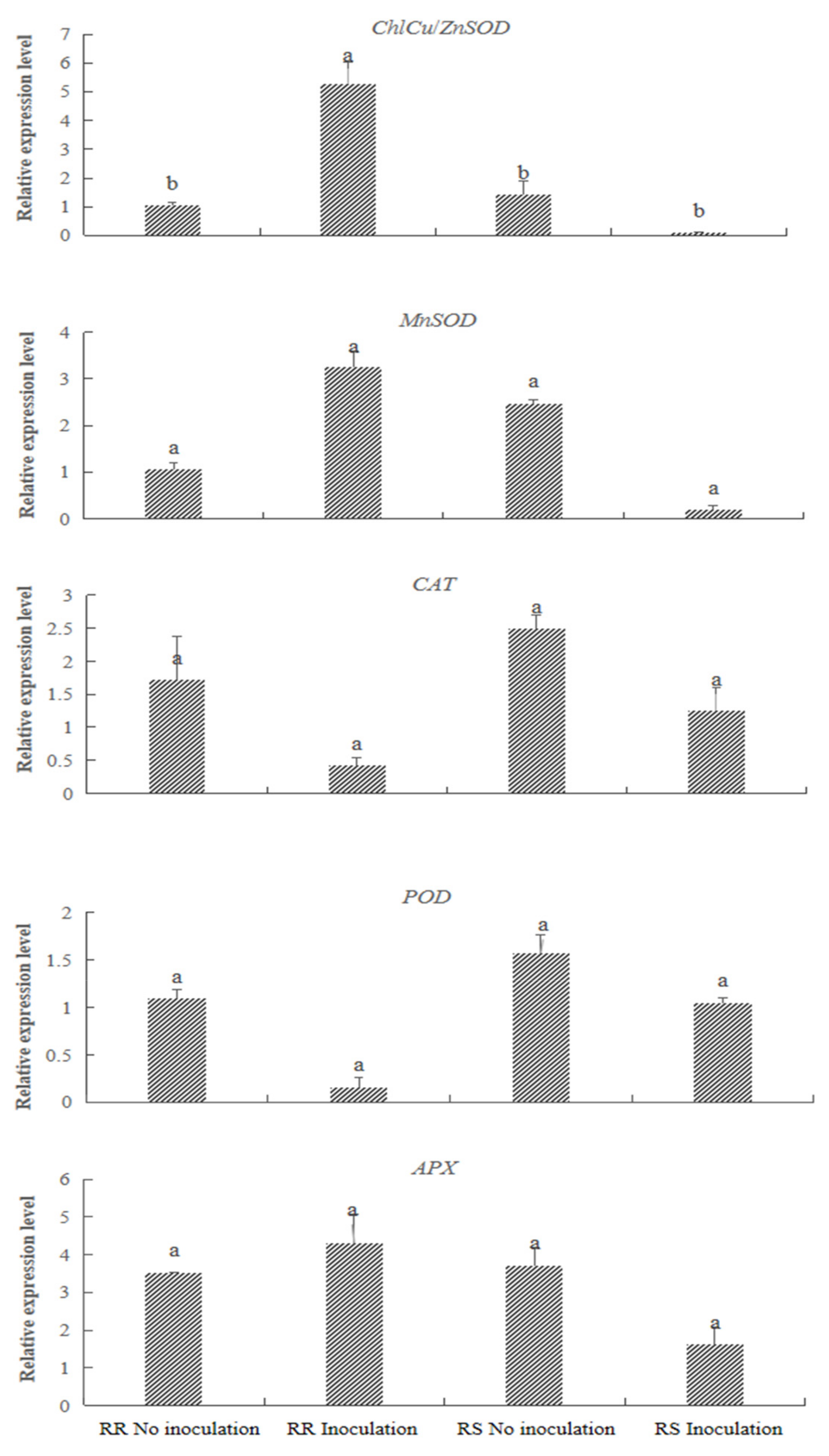
3.6. Gene Expression Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhardwaj, S.C.; Gangwar, O.P.; Prasad, P. Immunity to rusts in wheat: Theory, fact and practice. Indian Phytopathol. 2021, 74, 355–363. [Google Scholar] [CrossRef]
- He, Q.; Liu, J. Advancement in rust research on warm season turfgrass in China. Zhongguo Yesheng Zhiwu Ziyuan 2005, 24, 8–10. [Google Scholar]
- Yan, Y.; Li, W.P.; Gao, W.J.; Shen, Z.Y.; Wang, H.H.; Chen, L.X. Application of rDNA ITS sequence analysis in fungus identification. Zhongguo Weisheng Jianyan 2008, 18, 1958–1961. [Google Scholar]
- Iwen, P.; Hinrichs, S.; Rupp, M. Utilization of the internal transribed spacer regions as molecular targets to detect and identify human fungal oathogens. Med. Mycol. 2002, 40, 87–109. [Google Scholar] [CrossRef]
- Hou, J.; Chen, J.; Huang, J. Advances in molecular probes for biological detection of red tide. Zhongguo Gonggong Weisheng 2004, 20, 1127–1129. [Google Scholar]
- Commenil, P.; Brunet, L.; Audran, J.C. The development of the grape berry cuticle in relation to susceptibility to bunch rot disease. J. Exp. Bot. 1997, 48, 1599–1607. [Google Scholar] [CrossRef] [Green Version]
- Diaz, B.M.; Fereres, A. Ultraviolet-blocking materials as a physical barrier to control insect pests and plant pathogens in protected crops. Pest Technol. 2007, 1, 85–95. [Google Scholar]
- Li, M.; Li, W.; Sun, Y.; Mao, P.; Qi, X.; Wang, Y. Analysis of leaf tissue structures between rust-resistant and rust-susceptible Zoysiagrass (Zoysia japonica). Acta Physiol. Plant 2018, 40, 75. [Google Scholar] [CrossRef]
- Gill, S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Maksimović, J.; Zhang, J.; Zeng, F.; Shabala, L.; Zhou, M.; Shabala, S. Linking oxidative and salinity stress tolerance in barley: Can root antioxidant enzyme activity be used as a measure of stress tolerance? Plant Soil 2013, 365, 141–155. [Google Scholar] [CrossRef]
- Han, D. Screening Cultivars of Resistant to Rust and the Resistant Mechanism to Rust of Adzuki Bean. Master’s Thesis, Heilongjiang Bayi Agricultural University, Daqing, China, 2017. [Google Scholar]
- Yang, X.; Guo, H. Evaluation on trampling tolerance of two warm-season turfgrass varieties. Caoye Kexue 2018, 12, 54–62. [Google Scholar]
- Nakamura, N. The improvements of Zoysia and other warm-season grasses. Shibakusa Kenkyu 1980, 9, 113–117. [Google Scholar]
- Dong, H.; Gong, L.; Wang, Y.; Chen, Z.; Zhang, X.; Zhang, M. Ecology of Zoysia japonica and the Exploitation and Application of Its Resources in China; China Forestry Publishing House: Beijing, China, 2001. [Google Scholar]
- Chen, L.; Chen, L. A study on rust of Zoysia tenuifolia. Shanghai Nongxueyuan Xuebao 1997, 15, 133–136. [Google Scholar]
- Zhang, L.M. Identification of Zoysiagrass (Zoysia japonica Steud.) Germplasm to Rust and Screening of SSR Markers Linked with Resistant Genes. Master’s Thesis, China Agricultural University, Beijing, China, 2010. [Google Scholar]
- Nyassé, S.; Despréaux, D.; Cilas, C. Validity of a leaf inoculation test to assess the resistance to Phytophthora megakarya in a cocoa (Theobroma cacao L.) diallel mating design. Euphytica 2002, 123, 395–399. [Google Scholar] [CrossRef]
- Wang, Y.; Zhuang, J. Flora Fungorum Sinicorum; Science Press: Beijing, China, 1998. [Google Scholar]
- Chen, Y.Q.; Wang, N.; Zhou, H.; Qu, L.H.; Yang, L.F. Molecular identification and origin analysis on “red-tide” related Phaeocystis causative species. Haiyang Xuebao (Zhongwenban) 2002, 24, 99–103. [Google Scholar]
- Fan, S.X. Multi-Location Evaluation of Zoysiagrass (Zoysia japonica Steud.) Resistant Germplasm and Screening of SSR Markers with Resistant Genes. Master’s Thesis, China Agricultural University, Beijing, China, 2012. [Google Scholar]
- Gu, G. Germplasm Evaluation for Rust Resistance and Polymorphic Analysis of SSR Markers between Resistant and Susceptible Parents in Zoysiagrass (Zoysia japonica Steud.). Master’s Thesis, China Agricultural University, Beijing, China, 2013. [Google Scholar]
- Fang, Z. Research Methods for Plant Diseases; China Agriculture Press: Beijing, China, 1979. [Google Scholar]
- Zhang, J.; Kirkham, M.B. Drought-stress-induced changes in activities of superoxide dismutase, catalase, and peroxidase in wheat species. Plant Cell Physiol. 1994, 35, 785–791. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Schmittgen, T.; Livak, K. Analyzing real-time PCR data by the comparative CT method. Nat. Protocol. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Yue, X.; McLandsborough, L.; Zou, J.; Gao, X. Separation and identification of two fungal strains in stored Maize. Am. J. Biochem. Biotechnol. 2017, 13, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Li, B. Study on Identification of Pathogens Causing Hyacinth Soft Rot and Rust Symptoms and Disease Control. Master’s Thesis, Shanghai Jiao Tong University, Shanghai, China, 2011. [Google Scholar]
- Zhang, L.M.; Fang, C.; Zhang, X.Y.; Wang, Z.M.; Wang, Y.W. A study on the epidemic pattern of rust disease and germplasm resistance evaluation in Zoysia japonica in Beijing. Caoye Kexue 2011, 28, 279–285. [Google Scholar]
- Lu, J.; Chang, Y.; Li, B.; Shi, B. Determination and analysison 18S rDNA and ITSS sequences of two griseofulvin producing strains. Fujian Nonye Keji 2019, 2, 10–15. [Google Scholar]
- Zhou, S.; Lan, Y. Studies on the rust of Zoysia tenufolia. Nanjing Linye Daxue Xuebao 1991, 15, 40–45. [Google Scholar]
- Pence, H.; Bowling, A.; Breitweiser, L. Visualization of Asian Soybean Rust (Phakopsora pachyrhizi) fungal infections and cell ultrastructure to characterize fungciide action in situ. Microsc. Microanal. 2020, 26 (Suppl. S2), 354–355. [Google Scholar] [CrossRef]
- Yu, K. Molecule Detection of Puccinia polysora and Resistance Mechanism to Southern Corn Rust in Maize. Master’s Thesis, Northwest A & F University, Beijing, China, 2010. [Google Scholar]
- Anam, M.; Shahbaz, T.; Sajid, A.; Aman, U. Salicylic acid and jasmonic acid can suppress green and blue moulds of citrus fruit and induce the activity of polyphenol oxidase and peroxidase. Folia Hortic. 2019, 31, 195–204. [Google Scholar]
- Narayani, S.; Yashodhara, V. Enzymatic analysis of superoxide dismutase (SOD) from hordeum vulgare: Its role in drought stress tolerance. J. Plant Biochem. Physiol. 2019, 7, 1–4. [Google Scholar]
- Zhao, G.; Xu, Y.; Xu, Z.; Zhang, J.; Kong, W. Changes of related defense enzyme activity in resistance to melon powdery mildew. Zhongguo Nongxue Tongbao 2009, 25, 206–209. [Google Scholar]
- Li, Y.; Yu, X.; Bao, M.; Deng, X. Effects of huanglong pathogen infection on the activities of SOD and POD in periwinkles. Guangdong Nonye Kxuexi 2019, 46, 71–78. [Google Scholar]
- Zou, C.; Liu, C.; Wang, L.; Wang, H.; Xin, B.; Jiang, C. Correlation of resistance of different lines of pepper to Phytophthora capsici with activity indices of defense enzymes in leaf and soil enzymes in rhizosphere basis. Jilin Nongye Daxue Xuebao 2015, 37, 153–159. [Google Scholar]
- Zhou, C.; Zhu, C.; Fu, H. Genome-wide investigation of superoxide dismutase (SOD) gene family and their regulatory miRNAs reveal the involvement in abiotic stress and hormone response in tea plant (Camellia sinensis). PLoS ONE 2019, 14, e0223609. [Google Scholar] [CrossRef]
- Lee, W.; In, J.; Lee, B. Molecular characterization and expression of CuZn superoxide dismutase(PSOD1) from Populus alba × Populus glandulosa. Korean J. Plant Res. 2005, 8, 52–59. [Google Scholar]
- Palma, F.; He, C.; Danes, J. Mitochondrial superoxide dismutase: What the established, the intriguing, and the novel reveal about a key cellular redox switch. Antioxid. Redox Signal. 2020, 32, 701–714. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Jing, P.; Liu, P. Research progress on regulation of SOD gene expression activity and resistance to stress in plants. Shengwuxue Jiaoxue 2012, 37, 6–8. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | Source | Incidence Rate (%) | Disease Indexes | Incubation Period (d) | Rust Severity | Infection Type of Material |
|---|---|---|---|---|---|---|
| RS | Liaoning | 21.9 * | 3.90 * | 16–17 | 4 | 4 |
| RR | Shandong | 0 * | 0 * | - | 0 | 0 |
| 0 Days | 5 Days | 10 Days | 15 Days | 20 Days | |
|---|---|---|---|---|---|
| SOD | |||||
| Genotype (G) | ns | ** | ns | * | ** |
| Treatment (T) | ns | ** | ** | ** | ** |
| G × T | ns | ns | ns | ns | * |
| CAT | |||||
| Genotype (G) | ns | ns | * | ns | ** |
| Treatment (T) | ns | ** | ** | ** | ** |
| G × T | ns | ns | ns | ns | ns |
| POD | |||||
| Genotype (G) | ns | ** | ns | ns | ns |
| Treatment (T) | ns | ** | ns | * | ** |
| G × T | ns | ** | ns | ** | ** |
| APX | |||||
| Genotype (G) | ns | * | ** | ** | ns |
| Treatment (T) | ns | ** | ** | ** | ** |
| G × T | ns | ** | ** | ** | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Zeng, H.; Zhao, L.; Yue, J.; Qi, X.; Li, M. Isolation and Identification Rust Pathogens and the Study of Antioxidant Enzyme Activity and Gene Expression under Rust Infection in Zoysia japonica. Agriculture 2021, 11, 1200. https://doi.org/10.3390/agriculture11121200
Zhang D, Zeng H, Zhao L, Yue J, Qi X, Li M. Isolation and Identification Rust Pathogens and the Study of Antioxidant Enzyme Activity and Gene Expression under Rust Infection in Zoysia japonica. Agriculture. 2021; 11(12):1200. https://doi.org/10.3390/agriculture11121200
Chicago/Turabian StyleZhang, Di, Hanguo Zeng, Liyuan Zhao, Jiaming Yue, Xiao Qi, and Manli Li. 2021. "Isolation and Identification Rust Pathogens and the Study of Antioxidant Enzyme Activity and Gene Expression under Rust Infection in Zoysia japonica" Agriculture 11, no. 12: 1200. https://doi.org/10.3390/agriculture11121200