
Fertilization Regulates Accumulation and Allocation of Biomass and Nutrients in Phoebe bournei Seedlings

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Materials
2.3. Experimental Design
2.4. Experimental Management
2.5. Seedling Measurement Parameters
2.6. Data Processing and Analysis
2.6.1. Calculation of Comprehensive Index
2.6.2. Data Analysis
3. Results
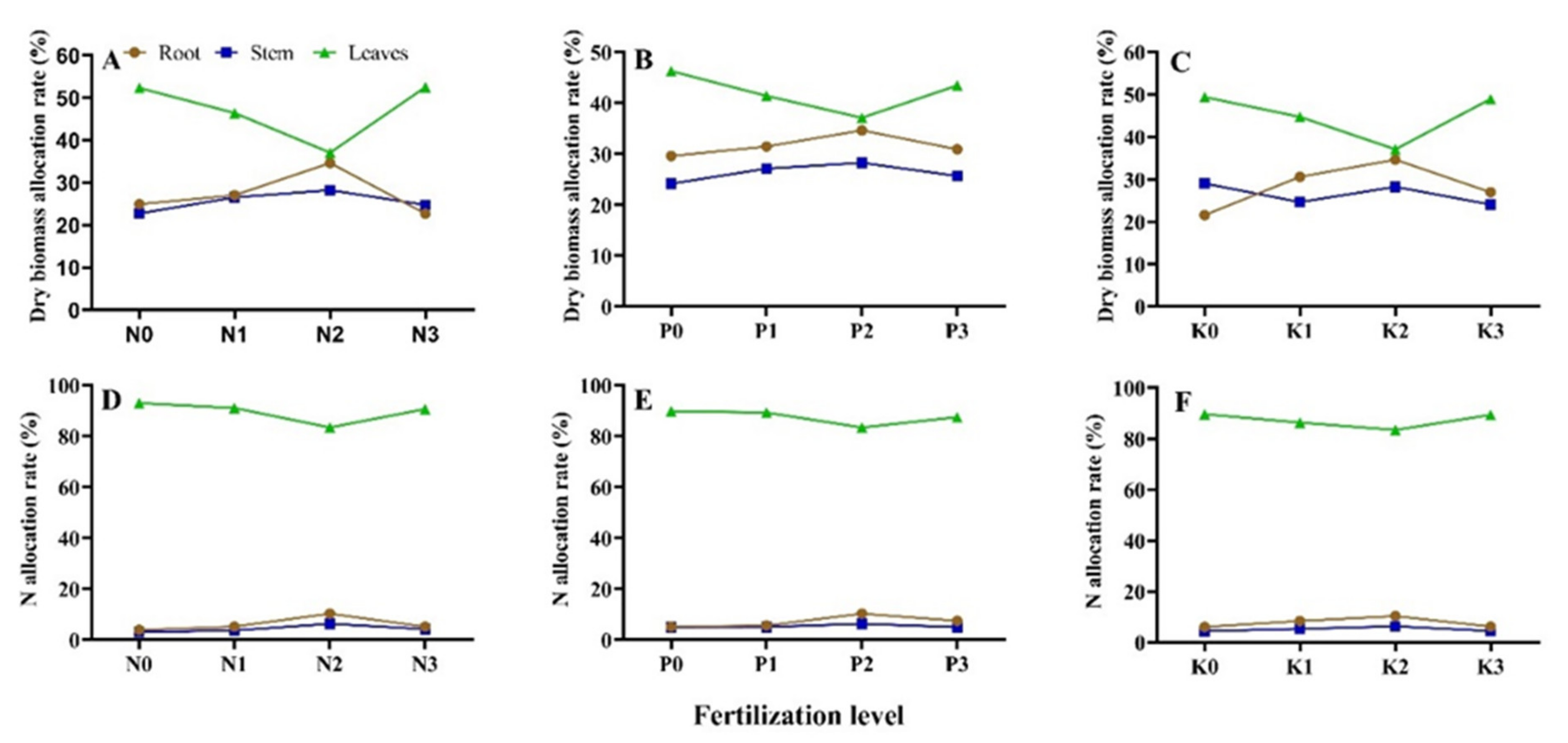
3.1. Seedlings Dry Biomass Allocation under Different N-P-K Fertilization
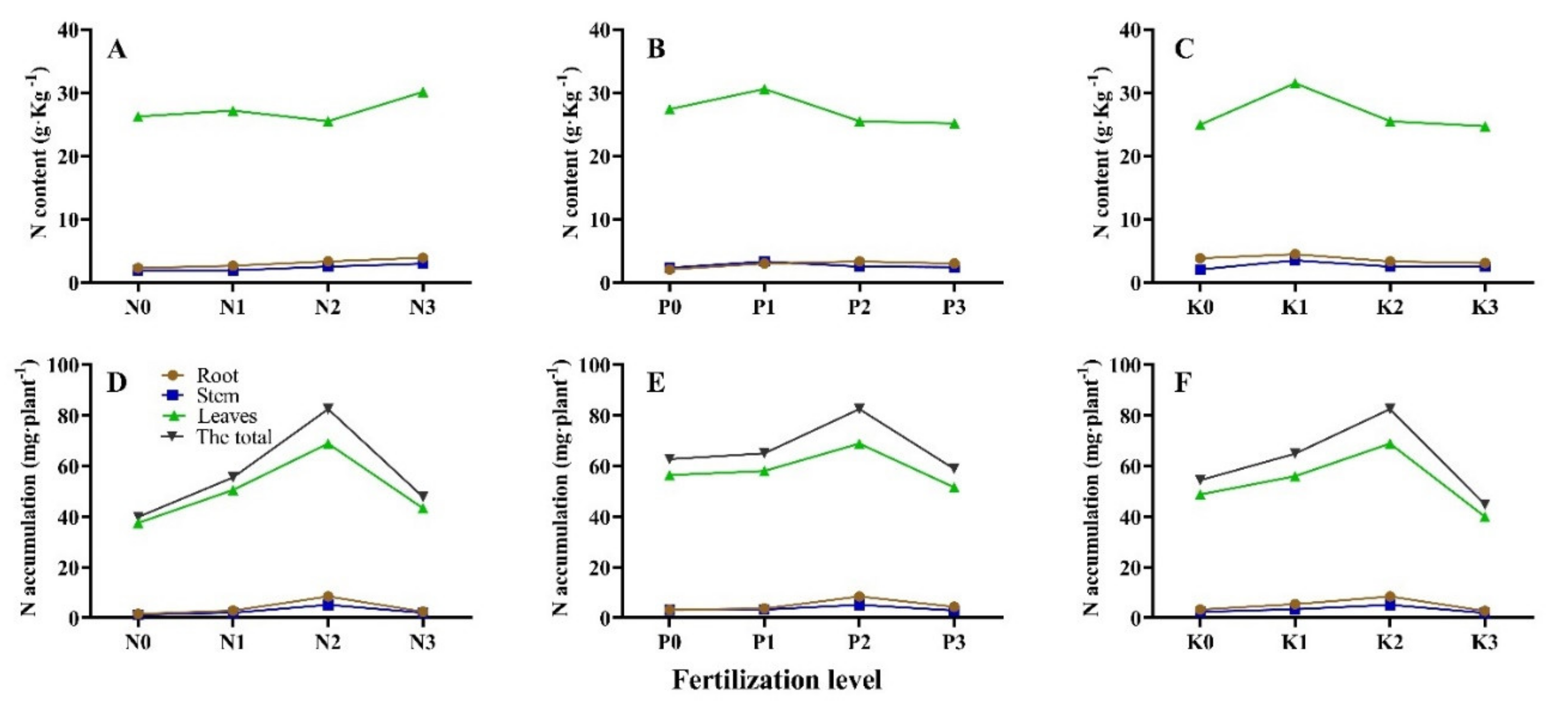
3.2. Seedling N Accumulation and Allocation under Different N-P-K Fertilization
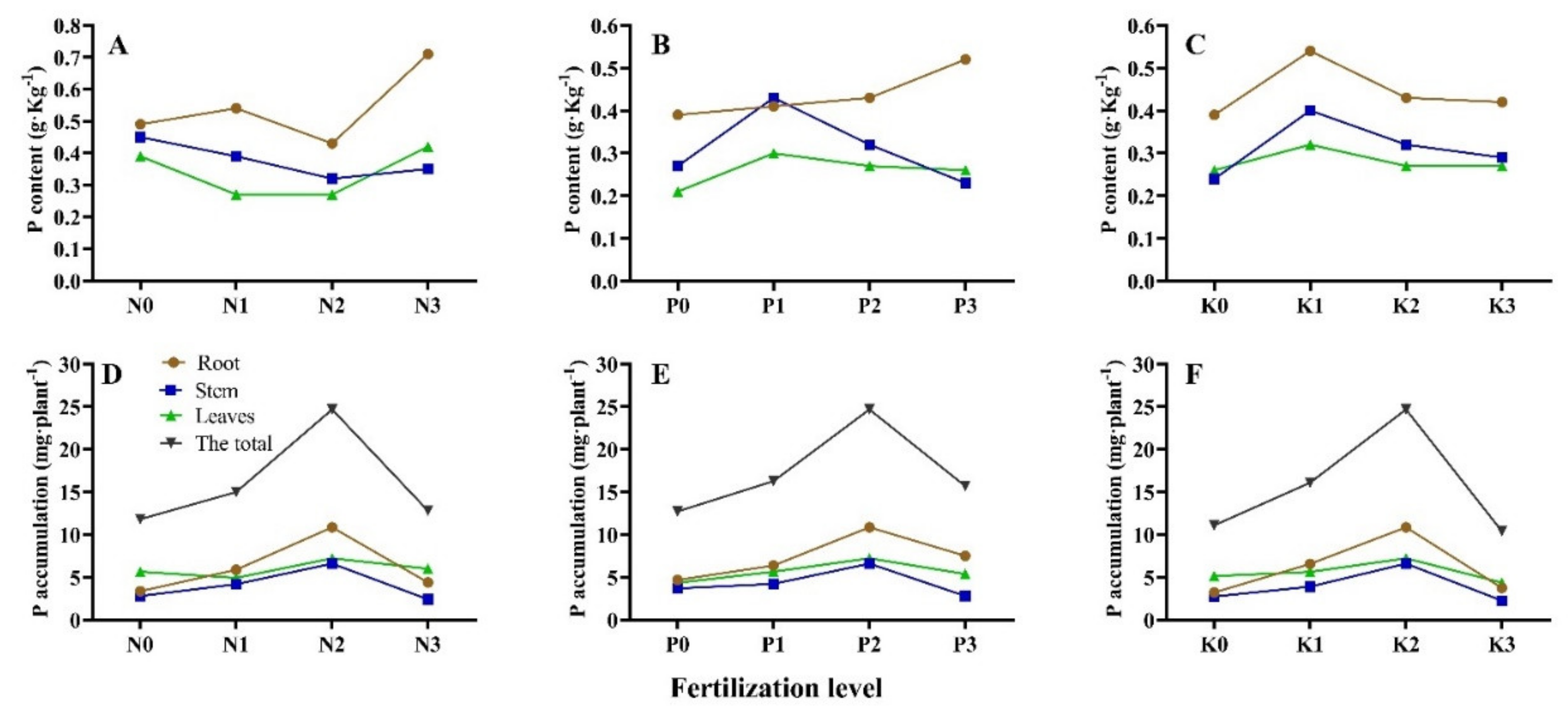
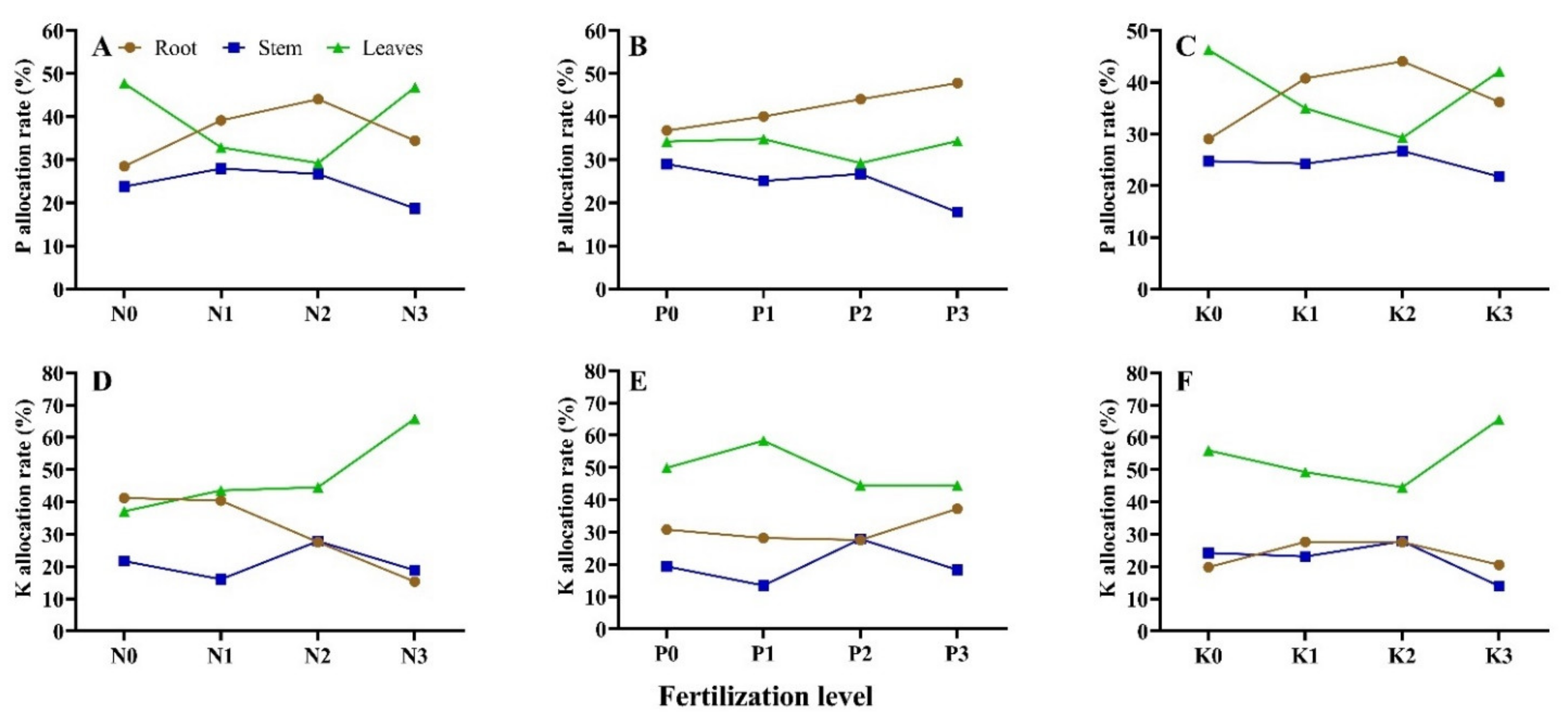
3.3. Seedling P Accumulation and Allocation under Different N-P-K Fertilization
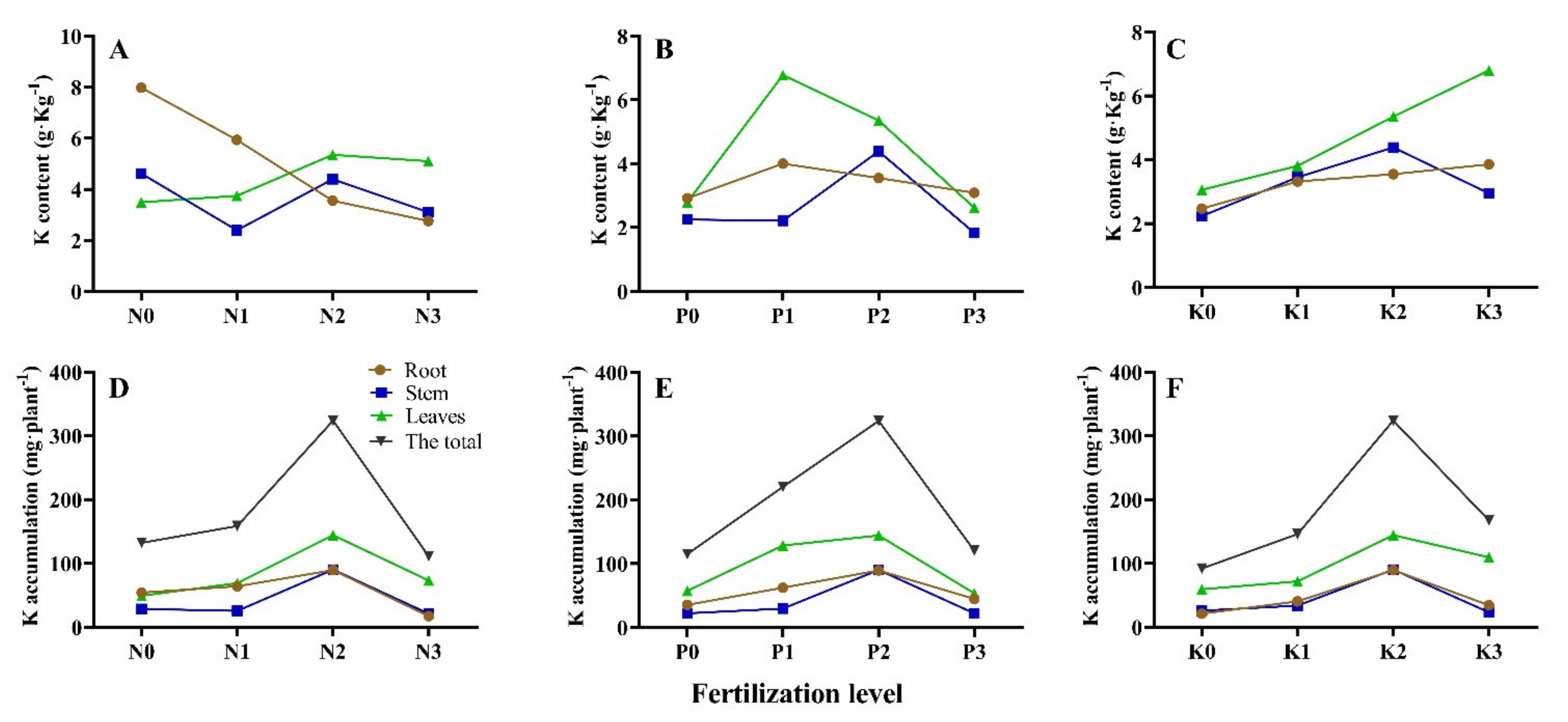
3.4. Seedling K Accumulation and Allocation under Different N-P-K Fertilization
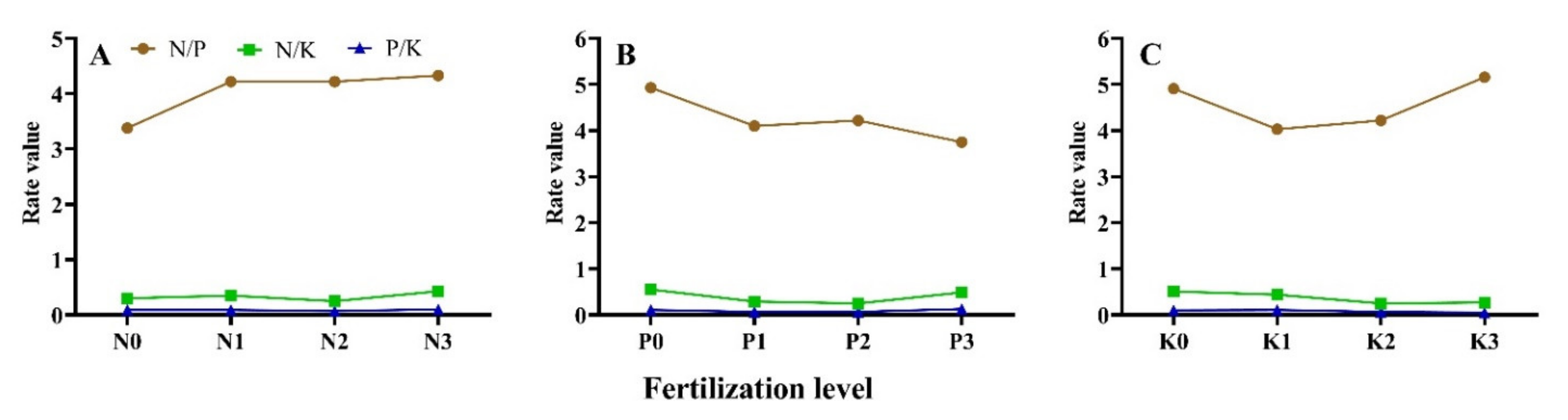
3.5. Seedling N, P, and K Element Absorption Balance under Different N-P-K Fertilization
3.6. Stepwise Regression and Path Analysis of Various Indexes under Different N-P-K Fertilization
3.7. Correlation Analysis between Dry Biomass Allocation Rate in Each Organ and Important Indexes of P. bournei Seedlings under N-P-K Fertilization
3.8. Seedling Range Analysis of Important Indexes under N-P-K Fertilization
4. Discussion
4.1. Effect of N Fertilizer on Nutrients Accumulation and Allocation in P. bournei Seedlings
4.2. Effect of P Fertilizer on Nutrients Accumulation and Allocation in P. bournei Seedlings
4.3. Effect of K Fertilizer on Nutrients Accumulation and Allocation in P. bournei Seedlings
4.4. N, P, and K Fertilizer Regulate Dry Biomass Accumulation and Allocation of P. bournei Seedlings
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Chrysargyris, A.; Panayiotou, C.; Tzortzakis, N. Nitrogen and phosphorus levels affected plant growth, essential oil composition and antioxidant status of lavender plant (Lavandula angustifolia Mill.). Ind. Crop. Prod. 2016, 83, 577–586. [Google Scholar] [CrossRef]
- Xu, J.; Wu, L.; Tong, B.; Yin, J.; Huang, Z.; Li, W.; Li, X. Magnesium supplementation alters leaf metabolic pathways for higher flavor quality of oolong tea. Agriculture 2021, 11, 120. [Google Scholar] [CrossRef]
- Hermans, C.; Hammond, J.; White, P.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant. Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef]
- Shalizi, M.N.; Goldfarb, B.; Burney, O.T.; Shear, T.H. Effects of five growing media and two fertilizer levels on polybag—Raised Camden Whitegum (Eucalyptus benthamii Maiden & Cambage) seedling morphology and drought hardiness. Forests 2019, 10, 543. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Ma, H.; Xia, J.; Hou, F.; Shi, X.; Hao, X.; Hafeez, A.; Han, H.; Luo, H. Optimal pre-plant irrigation and fertilization can improve biomass accumulation by maintaining the root and leaf productive capacity of cotton crop. Sci. Rep. 2017, 7, 17168. [Google Scholar] [CrossRef] [Green Version]
- Vandamme, E.; Wissuwa, M.; Rose, T.; Ahouanton, K.; Saito, K. Strategic phosphorus (P) application to the nursery bed increases seedling growth and yield of transplanted rice at low P supply. Field Crop. Res. 2016, 186, 10–17. [Google Scholar] [CrossRef]
- Oliet, J.A.; Puértolas, J.; Planelles, R.; Jacobs, D.F. Nutrient loading of forest tree seedlings to promote stress resistance and field performance: A Mediterranean perspective. New For. 2013, 44, 649–669. [Google Scholar] [CrossRef]
- Bernstein, N.; Gorelick, J.; Zerahia, R.; Koch, S. Impact of N, P, K, and humic acid supplementation on the chemical profile of medical Cannabis (Cannabis sativa L.). Front. Plant. Sci. 2019, 10, 736. [Google Scholar] [CrossRef] [Green Version]
- Ladha, J.; Reddy, P.M. Nitrogen fixation in rice systems: State of knowledge and future prospects. Plant. Soil 2003, 252, 151–167. [Google Scholar] [CrossRef]
- Raza, M.A.; Bin Khalid, M.H.; Zhang, X.; Feng, L.Y.; Khan, I.; Hassan, M.J.; Ahmed, M.; Ansar, M.; Chen, Y.K.; Fan, Y.F.; et al. Effect of planting patterns on yield, nutrient accumulation and distribution in maize and soybean under relay intercropping systems. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Harmand, J.-M.; Njiti, C.F.; Bernhard-Reversat, F.; Puig, H. Aboveground and belowground biomass, productivity and nutrient accumulation in tree improved fallows in the dry tropics of Cameroon. For. Ecol. Manag. 2004, 188, 249–265. [Google Scholar] [CrossRef]
- Deng, S.; Shi, K.; Ma, J.; Zhang, L.; Ma, L.; Jia, Z. Effects of fertilization ratios and frequencies on the growth and nutrient uptake of Magnolia wufengensis (Magnoliaceae). Forests 2019, 10, 65. [Google Scholar] [CrossRef] [Green Version]
- Pogrzeba, M.; Rusinowski, S.; Krzyżak, J. Macroelements and heavy metals content in energy crops cultivated on contaminated soil under different fertilization—Case studies on autumn harvest. Environ. Sci. Pollut. Res. 2018, 25, 12096–12106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.; Jiang, X.; Ma, M.; Zhou, B.; Guan, D.; Zhao, B.; Zhou, J.; Cao, F.; Li, L.; Li, J. Effect of 35 years inorganic fertilizer and manure amendment on structure of bacterial and archaeal communities in black soil of northeast China. Appl. Soil Ecol. 2016, 105, 187–195. [Google Scholar] [CrossRef]
- Chen, B.; Wang, Q.; Ye, Z.; Stiles, S.; Feng, G. Optimisation of phosphorus fertilization promotes biomass and phosphorus nutrient accumulation, partitioning and translocation in three cotton (Gossypium hirsutum) genotypes. Crop. Pasture Sci. 2020, 71, 56–69. [Google Scholar] [CrossRef]
- Szczepanek, M.; Siwik-Ziomek, A. P and K accumulation by rapeseed as affected by biostimulant under different NPK and S ferti-lization doses. Agronomy 2019, 9, 477. [Google Scholar] [CrossRef] [Green Version]
- Soratto, R.P.; Job, A.L.G.; Fernandes, A.; Assunção, N.S.; Fernandes, F.M. Biomass accumulation and nutritional requirements of potato as affected by potassium supply. J. Soil Sci. Plant. Nutr. 2020, 20, 1051–1066. [Google Scholar] [CrossRef]
- Li, M.; Zhang, H.; Yang, X.; Ge, M.; Ma, Q.; Wei, H.; Dai, Q.; Huo, Z.; Xu, K.; Luo, D. Accumulation and utilization of nitrogen, phosphorus and potassium of irrigated rice cultivars with high productivities and high N use efficiencies. Field Crop. Res. 2014, 161, 55–63. [Google Scholar] [CrossRef]
- Ju, X.; Kou, C.; Christie, P.; Dou, Z.; Zhang, F. Changes in the soil environment from excessive application of fertilizers and manures to two contrasting intensive cropping systems on the North China Plain. Environ. Pollut. 2007, 145, 497–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamil, K.R.; Anita, K.; Roman, K.K.; Konieczna, A. Evaluation of a different fertilisation in technology of corn for silage, sugar beet and meadow grasses production and their impact on the environment in Poland. Afr. J. Agric. Res. 2015, 10, 1351–1358. [Google Scholar] [CrossRef] [Green Version]
- Soratto, R.P.; Fernandes, A.M. Phosphorus effects on biomass accumulation and nutrient uptake and removal in two potato culti-vars. Agron. J. 2016, 108, 1225–1236. [Google Scholar] [CrossRef] [Green Version]
- Malhi, S.S.; Johnston, A.M.; Schoenau, J.J.; Wang, Z.L.; Vera, C.L. Seasonal biomass accumulation and nutrient uptake of wheat, barley and oat on a Black Chernozem Soil in Saskatchewan. Can. J. Plant. Sci. 2006, 86, 1005–1014. [Google Scholar] [CrossRef]
- Wang, X.; Wei, X.; Wu, G.; Chen, S. High nitrate or ammonium applications alleviated photosynthetic decline of Phoebe bournei seedlings under elevated carbon dioxide. Forests 2020, 11, 293. [Google Scholar] [CrossRef] [Green Version]
- Ding, W.; Liping, N.; Xing, H.; Wei, Z.; Zhoua, Q.; Nong, R.; Chen, J. Essential oil extracted from leaf of Phoebe bournei (Hemsl.) yang: Chemical constituents, antitumor, antibacterial, hypoglycemic activities. Nat. Prod. Res. 2018, 34, 2524–2527. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Shang, H.; Chen, Z.; Tian, Y.; Yu, H. Effects of elevated ozone on stoichiometry and nutrient pools of Phoebe bournei (Hemsl.) Yang and Phoebe Zhennan, S. Lee et F. N. Wei seedlings in subtropical China. Forests 2016, 7, 78. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhu, Y.; Pan, Y.; Huang, H.; Li, C.; Li, G.; Tong, Z. Transcriptomic profiling and identification of candidate genes in two Phoebe bournei ecotypes with contrasting cold stress responses. Trees 2018, 32, 1315–1333. [Google Scholar] [CrossRef]
- He, S.; Ma, Y.; Fei, Y. Research on the species composition and features of Phoebe plant community in Laifeng County, Hubei Province. J. Mt. Agri. Biol. 2017, 36, 45–49. [Google Scholar]
- Wang, W.; Wei, X.A.; Liao, W.; Blanco, J.; Liu, Y.; Liu, S.; Liu, G.; Zhang, L.; Guo, X.; Guo, S. Evaluation of the effects of forest management strategies on carbon sequestration in evergreen broad-leaved (Phoebe bournei) plantation forests using FORECAST ecosystem model. For. Ecol. Manag. 2013, 300, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Fan, R.; Sun, J.; Yang, F.; Quanlin, Z.; Zheng, Y.; Zhong, Q.; Cheng, D. Divergent scaling of respiration rates to nitrogen and phosphorus across four woody seedlings between different growing seasons. Ecol. Evol. 2017, 7, 8761–8769. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Zhu, X.; Lin, X.; Xu, Z. Effects of fertilization on the growth of three Lauraceae seedlings in containers. North. Hortic. 2011, 13, 73–77. [Google Scholar]
- Li, J.; Zhang, M.; Chen, F.; Yao, B. Improving rice modeling success rate with ternary non-structural fertilizer response model. Sci. Rep. 2018, 8, 9071. [Google Scholar] [CrossRef]
- Yang, Z.-J.; Wu, X.-H.; Grossnickle, S.C.; Chen, L.-H.; Yu, X.-X.; El-Kassaby, Y.A.; Feng, J.-L. Formula fertilization promotes Phoebe bournei robust seedling cultivation. Forests 2020, 11, 781. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Chapagain, T.; Ghimire, B.; Pudasaini, R.; Tamang, B.B.; Gurung, K.; Choi, K.; Rai, L.; Magar, S.; Bk, B.; et al. Evaluating the effectiveness of rhizobium inoculants and micronutrients as technologies for nepalese common bean smallholder farmers in the real-world context of highly variable hillside environments and indigenous farming practices. Agriculture 2019, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Guo, X.; Yun, W.; Xia, Y.; You, Z.; Rillig, M. Arbuscular mycorrhiza contributes to the control of phosphorus loss in paddy fields. Plant. Soil 2020, 447, 623–636. [Google Scholar] [CrossRef]
- Artyszak, A.; Gozdowski, D. Influence of various forms of foliar application on root yield and technological quality of sugar beet. Agriculture 2021, 11, 693. [Google Scholar] [CrossRef]
- Wang, X.; Bian, S.; Chang, P.; Wang, N.; Xuan, L.; Zhang, M.; Dong, B.; Zhang, C.; Wu, J.; Ying, Y.; et al. The effects of different nitrogen sources on camptothecin content and related gene expression in Camptotheca acuminata seedlings. J. For. Res. 2019, 31, 1347–1357. [Google Scholar] [CrossRef]
- Barunawati, N.; Giehl, R.F.H.; Bauer, B.; von Wirén, N. The influence of inorganic nitrogen fertilizer forms on micronutrient retranslocation and accumulation in grains of winter wheat. Front. Plant. Sci. 2013, 4, 320. [Google Scholar] [CrossRef] [Green Version]
- Ou, X.; Li, S.; Liao, P.; Cui, X.; Zheng, B.; Yang, Y.; Liu, D.; Zheng, Y. The transcriptome variations of Panax notoginseng roots treated with different forms of nitrogen fertilizers. BMC Genom. 2019, 20, 965. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Ding, M.; Lu, W.; Lu, D. Nitrogen topdressing at the jointing stage affects the nutrient accumulation and translocation in rainfed waxy maize. J. Plant. Nutr. 2019, 42, 657–672. [Google Scholar] [CrossRef]
- Bélanger, G.; Ziadi, N.; Lajeunesse, J.; Jouany, C.; Virkajarvi, P.; Sinaj, S.; Nyiraneza, J. Shoot growth and phosphorus–nitrogen relationship of grassland swards in response to mineral phosphorus fertilization. Field Crop. Res. 2017, 204, 31–41. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Nahar, K.; Hossain, M.S.; Mahmud, J.A.; Hossen, M.S.; Masud, A.A.C.; Moumita; Fujita, M. Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Bassi, D.; Menossi, M.; Mattiello, L. Nitrogen supply influences photosynthesis establishment along the sugarcane leaf. Sci. Rep. 2018, 8, 2327. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xie, D.; Ni, J.; Zeng, X. Optimizing phosphate fertilizer application to reduce nutrient loss in a mustard (Brassica juncea var. tumida)-maize (Zea mays L.) rotation system in Three Gorges Reservoir area. Soil Tillage Res. 2019, 190, 78–85. [Google Scholar] [CrossRef]
- Zheng, H.; Pan, X.; Deng, Y.; Wu, H.; Liu, P.; Li, X. AtOPR3 specifically inhibits primary root growth in Arabidopsis under phosphate deficiency. Sci. Rep. 2016, 6, 24778. [Google Scholar] [CrossRef] [Green Version]
- Carstensen, A.; Herdean, A.; Schmidt, S.B.; Sharma, A.; Spetea, C.; Pribil, M.; Husted, S. The impacts of phosphorus deficiency on the photosynthetic electron transport chain. Plant. Physiol. 2018, 177, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Chu, C. Nitrogen–phosphorus interplay: Old story with molecular tale. New Phytol. 2020, 225, 1455–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chrysargyris, A.; Xylia, P.; Botsaris, G.; Tzortzakis, N. Antioxidant and antibacterial activities, mineral and essential oil composition of spearmint (Mentha spicata L.) affected by the potassium levels. Ind. Crop. Prod. 2017, 103, 202–212. [Google Scholar] [CrossRef]
- Hu, W.; Jiang, N.; Yang, J.; Meng, Y.; Wang, Y.; Chen, B.; Zhao, W.; Oosterhuis, D.M.; Zhou, Z. Potassium (K) supply affects K accumulation and photosynthetic physiology in two cotton (Gossypium hirsutum L.) cultivars with different K sensitivities. Field Crop. Res. 2016, 196, 51–63. [Google Scholar] [CrossRef]
- Wang, X.; Mohamed, I.; Ali, M.; Abbas, M.H.H.; Shah, G.M.; Chen, F. Potassium distribution in root and non-root zones of two cotton genotypes and its accumulation in their organs as affected by drought and potassium stress conditions. J. Plant. Nutr. Soil Sci. 2018, 182, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Zahoor, R.; Dong, H.; Abid, M.; Zhao, W.; Wang, Y.; Zhou, Z. Potassium fertilizer improves drought stress alleviation potential in cotton by enhancing photosynthesis and carbohydrate metabolism. Environ. Exp. Bot. 2017, 137, 73–83. [Google Scholar] [CrossRef]
- Sheng, X. Growth promotion and increased potassium uptake of cotton and rape by a potassium releasing strain of Bacillus edaphicus. Soil Biol. Biochem. 2005, 37, 1918–1922. [Google Scholar] [CrossRef]
- Rogiers, S.Y.; Coetzee, Z.; Walker, R.R.; Deloire, A.; Tyerman, S. Potassium in the Grape (Vitis vinifera L.) Berry: Transport and function. Front. Plant. Sci. 2017, 8, 1629. [Google Scholar] [CrossRef]
- Bu, M.A.H.A.; Ahmad, R.; Iqbal, J.; Maqbool, M.M.; Ali, A.; Ishaque, M.; Hussain, S. Nutritional and physiological significance of potassium application in maize hybrid crop production. Pak. J. Nutr. 2012, 11, 187–202. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.H.; Jaafar, H.Z.E.; Karimi, E.; Ghasemzadeh, A. Primary, secondary metabolites, photosynthetic capacity and antioxidant activity of the Malaysian herb Kacip Fatimah (Labisia Pumila Benth) exposed to potassium fertilization under greenhouse conditions. Int. J. Mol. Sci. 2012, 13, 15321–15342. [Google Scholar] [CrossRef]
- Wu, J.; Lin, H.; Guo, L.; Dong, J.; Zhang, L.; Fu, W. Biomass and nutrients variation of Chinese fir rooted cuttings under conventional and exponential fertilization regimes of nitrogen. Forests 2019, 10, 615. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I.; Hengeler, C.; Marschner, H. Partitioning of shoot and root dry matter and carbohydrates in bean plants suffering from phosphorus, potassium and magnesium deficiency. J. Exp. Bot. 1994, 45, 1245–1250. [Google Scholar] [CrossRef]
- Geng, Y.; Cao, G.; Wang, L.; Wang, S. Effects of equal chemical fertilizer substitutions with organic manure on yield, dry matter, and nitrogen uptake of spring maize and soil nitrogen distribution. PLoS ONE 2019, 14, e0219512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hocking, P.; Randall, P.; DeMarco, D. The response of dryland canola to nitrogen fertilizer: Partitioning and mobilization of dry matter and nitrogen, and nitrogen effects on yield components. Field Crop. Res. 1997, 54, 201–220. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Treatment 1 | N (g·plant−1) 1 | P2O5 (g·plant−1) 1 | KO2 (g·plant−1) 1 |
|---|---|---|---|---|
| T1 | N0P0K0 | 0 (0) | 0 (0) | 0 (0) |
| T2 | N0P2K2 | 0 (0) | 2 (0.1332) | 2 (0.356) |
| T3 | N1P2K2 | 1 (0.266) | 2 (0.1332) | 2 (0.356) |
| T4 | N2P0K2 | 2 (0.532) | 0 (0) | 2 (0.356) |
| T5 | N2P1K2 | 2 (0.532) | 1 (0.0666) | 2 (0.356) |
| T6 | N2P2K2 | 2 (0.532) | 2 (0.1332) | 2 (0.356) |
| T7 | N2P3K2 | 2 (0.532) | 3 (0.1998) | 2 (0.356) |
| T8 | N2P2K0 | 2 (0.532) | 2 (0.1332) | 0 (0) |
| T9 | N2P2K1 | 2 (0.532) | 2 (0.1332) | 1 (0.178) |
| T10 | N2P2K3 | 2 (0.532) | 2 (0.1332) | 3 (0.534) |
| T11 | N3P2K2 | 3 (0.798) | 2 (0.1332) | 2 (0.356) |
| T12 | N1P1K2 | 1 (0.266) | 1 (0.0666) | 2 (0.356) |
| T13 | N1P2K1 | 1 (0.266) | 2 (0.1332) | 1 (0.178) |
| T14 | N2P1K1 | 2 (0.532) | 1 (0.0666) | 1 (0.178) |
| No. | Root/g·plant−1 | Stem/g·plant−1 | Leaf/g·plant−1 | Total Plant/g·plant−1 |
|---|---|---|---|---|
| T1 | 0.83 ± 0.05 f | 0.80 ± 0.09 g | 1.26 ± 0.09 g | 2.89 ± 0.23 e |
| T2 | 0.68 ± 0.06 g | 0.62 ± 0.04 h | 1.43 ± 0.12 g | 2.73 ± 0.27 e |
| T3 | 1.08 ± 0.09 e | 1.06 ± 0.09 de | 1.86 ± 0.13 de | 4.01 ± 0.32 c |
| T4 | 1.56 ± 0.06 b | 1.35 ± 0.04 b | 2.06 ± 0.04 bcd | 4.97 ± 0.14 b |
| T5 | 1.21 ± 0.02 d | 0.99 ± 0.03 ef | 1.90 ± 0.03 cde | 4.10 ± 0.07 c |
| T6 | 2.52 ± 0.04 a | 2.06 ± 0.14 a | 2.70 ± 0.20 a | 7.27 ± 0.41 a |
| T7 | 1.45 ± 0.05 c | 1.21 ± 0.03 c | 2.05 ± 0.16 bcd | 4.71 ± 0.34 b |
| T8 | 0.85 ± 0.01 f | 1.15 ± 0.01 cd | 1.95 ± 0.02 cde | 3.95 ± 0.14 c |
| T9 | 1.22 ± 0.07 d | 0.98 ± 0.06 ef | 1.78 ± 0.06 ef | 3.97 ± 0.20 c |
| T10 | 0.89 ± 0.05 f | 0.79 ± 0.03 g | 1.62 ± 0.15 f | 3.30 ± 0.12 d |
| T11 | 0.62 ± 0.04 g | 0.68 ± 0.05 h | 1.44 ± 0.12 g | 2.73 ± 0.16 e |
| T12 | 1.52 ± 0.11 bc | 1.10 ± 0.02 cd | 2.03 ± 0.01 bcd | 4.65 ± 0.32 b |
| T13 | 1.09 ± 0.09 e | 1.40 ± 0.04 b | 2.20 ± 0.07 b | 4.69 ± 0.12 b |
| T14 | 1.19 ± 0.03 de | 0.95 ± 0.03 f | 2.06 ± 0.06 bc | 4.20 ± 0.12 c |
| No. | Dry Biomass Allocation Rate (%) | N Allocation Rate (%) | ||||
|---|---|---|---|---|---|---|
| Root | Stem | Leaves | Root | Stem | Leaves | |
| T1 | 28.67 ± 0.65 ef 1 | 27.66 ± 0.97 cd | 43.67 ± 0.32 de | 4.54 ± 0.25 g | 4.69 ± 0.23 bc | 90.77 ± 0.41 bc |
| T2 | 24.92 ± 0.26 h | 22.76 ± 0.36 i | 52.32 ± 0.11 a | 3.90 ± 0.10 h | 3.08 ± 0.24 e | 93.02 ± 0.19 a |
| T3 | 27.03 ± 0.19 g | 26.53 ± 0.23 e | 46.44 ± 0.42 c | 5.21 ± 0.16 f | 3.71 ± 0.08 de | 91.08 ± 0.15 b |
| T4 | 29.59 ± 0.05 de | 24.14 ± 0.21 gh | 46.26 ± 0.16 c | 5.15 ± 0.21 fg | 4.98 ± 0.62 bc | 89.87 ± 0.71 bcd |
| T5 | 31.47 ± 0.32 c | 27.11 ± 0.12 de | 41.42 ± 0.44 f | 5.63 ± 0.03 ef | 5.05 ± 0.10 bc | 89.32 ± 0.10 cde |
| T6 | 34.64 ± 1.24 a | 28.26 ± 0.41 bc | 37.09 ± 0.83 g | 10.28 ± 0.46 a | 6.27 ± 0.89 a | 83.45 ± 1.10 h |
| T7 | 30.89 ± 0.46 c | 25.66 ± 0.69 f | 43.44 ± 1.14 e | 7.45 ± 0.42 c | 5.02 ± 0.41 bc | 87.52 ± 0.83 fg |
| T8 | 21.58 ± 0.06 j | 29.01 ± 0.02 b | 49.41 ± 0.04 b | 6.01 ± 0.07 de | 4.39 ± 0.08 bcd | 89.61 ± 0.14 cde |
| T9 | 30.61 ± 0.29 cd | 24.65 ± 0.39 g | 44.74 ± 0.68 d | 8.40 ± 0.86 b | 5.30 ± 0.10 b | 86.31 ± 0.84 g |
| T10 | 26.98 ± 0.26 g | 24.08 ± 0.82 gh | 48.94 ± 1.08 b | 6.18 ± 0.47 de | 4.43 ± 0.25 bcd | 89.39 ± 0.68 cde |
| T11 | 22.70 ± 0.39 i | 24.76 ± 0.06 g | 52.54 ± 0.44 a | 5.10 ± 0.12 fg | 4.23 ± 0.18 cd | 90.67 ± 0.18 bc |
| T12 | 32.64 ± 1.28 b | 23.64 ± 0.25 h | 43.72 ± 1.04 de | 6.02 ± 0.35 de | 4.95 ± 0.11 bc | 89.04 ± 0.35 de |
| T13 | 23.16 ± 0.96 i | 29.88 ± 0.35 a | 46.96 ± 0.61 c | 5.19 ± 0.37 fg | 5.27 ± 1.03 b | 89.54 ± 1.24 cde |
| T14 | 28.26 ± 0.07 f | 22.62 ± 0.13 i | 49.13 ± 0.05 b | 6.63 ± 0.25 d | 5.08 ± 0.84 bc | 88.29 ± 2.25 ef |
| No. | N Content/g·kg −1 | N Accumulation/mg·plant−1 | |||||
|---|---|---|---|---|---|---|---|
| Root | Stem | Leaves | Root | Stem | Leaves | Total | |
| T1 | 1.72 ± 0.08 j 1 | 1.85 ± 0.01 e | 22.65 ± 0.48 g | 1.43 ± 0.13 i | 1.48 ± 0.16 ef | 28.52 ± 1.80 h | 31.43 ± 2.05 g |
| T2 | 2.31 ± 0.08 hi | 1.93 ± 0.01 e | 26.25 ± 0.40 d | 1.57 ± 0.18 i | 1.24 ± 0.04 f | 37.47 ± 3.53 g | 39.87 ± 4.31 f |
| T3 | 2.67 ± 0.07 g | 1.94 ± 0.03 e | 27.17 ± 0.24 c | 2.89 ± 0.22 g | 2.06 ± 0.20 def | 50.55 ± 3.84 de | 55.50 ± 4.24 d |
| T4 | 2.07 ± 0.06 i | 2.32 ± 0.31 de | 27.40 ± 0.04 c | 3.23 ± 0.20 f | 3.12 ± 0.41 bc | 56.36 ± 0.94 bc | 62.71 ± 1.22 bc |
| T5 | 3.01 ± 0.02 f | 3.31 ± 0.04 abc | 30.59 ± 0.05 b | 3.66 ± 0.05 de | 3.28 ± 0.11 bc | 58.02 ± 0.69 b | 64.96 ± 0.84 b |
| T6 | 3.36 ± 0.01 de | 2.52 ± 0.41 bcde | 25.50 ± 0.31 e | 8.46 ± 0.14 a | 5.16 ± 0.72 a | 68.83 ± 4.73 a | 82.45 ± 4.85 a |
| T7 | 3.01 ± 0.06 f | 2.44 ± 0.10 cde | 25.16 ± 0.33 ef | 4.37 ± 0.14 c | 2.94 ± 0.11 bcd | 51.53 ± 4.71 cde | 58.85 ± 4.84 cd |
| T8 | 3.83 ± 0.04 bc | 2.08 ± 0.04 de | 24.95 ± 0.01 ef | 3.27 ± 0.07 f | 2.39 ± 0.07 cde | 48.74 ± 0.57 e | 54.39 ± 0.70 d |
| T9 | 4.49 ± 0.60 a | 3.51 ± 0.15 a | 31.49 ± 0.33 a | 5.43 ± 0.40 b | 3.44 ± 0.15 b | 55.96 ± 2.28 bcd | 64.83 ± 2.05 bc |
| T10 | 3.09 ± 0.06 ef | 2.49 ± 0.09 cde | 24.71 ± 1.14 f | 2.75 ± 0.15 gh | 1.97 ± 0.12 def | 40.00 ± 4.95 fg | 44.73 ± 5.19 ef |
| T11 | 3.92 ± 0.05 b | 2.98 ± 0.11 abcd | 30.13 ± 0.37 b | 2.43 ± 0.14 h | 2.02 ± 0.20 def | 43.28 ± 3.49 f | 47.73 ± 3.81 e |
| T12 | 2.47 ± 0.02 gh | 2.81 ± 0.09 abcde | 27.35 ± 0.24 c | 3.76 ± 0.28 d | 3.09 ± 0.10 bc | 55.61 ± 0.73 bcd | 62.46 ± 1.05 bc |
| T13 | 3.15 ± 0.25 ef | 2.48 ± 0.51 cde | 26.75 ± 0.35 cd | 3.41 ± 0.27 ef | 3.49 ± 0.81 b | 58.87 ± 2.25 b | 65.76 ± 3.05 b |
| T14 | 3.58 ± 0.11 cd | 3.44 ± 1.73 ab | 27.40 ± 0.38 c | 4.24 ± 0.12 c | 3.26 ± 1.61 bc | 56.54 ± 1.37 bc | 64.04 ± 1.24 bc |
| No. | P Content/g·kg−1 | P Accumulation/mg·plant−1 | |||||
|---|---|---|---|---|---|---|---|
| Root | Stem | Leaves | Root | Stem | Leaves | Total | |
| T1 | 0.35 ± 0.03 g 1 | 0.19 ± 0.00 i | 0.37 ± 0.02 b | 2.87 ± 0.07 h | 1.54 ± 0.19 e | 4.72 ± 0.44 de | 9.14 ± 0.56 g |
| T2 | 0.49 ± 0.01 c | 0.45 ± 0.01 a | 0.39 ± 0.04 ab | 3.35 ± 0.28 gh | 2.79 ± 0.16 cd | 5.66 ± 1.02 cd | 11.80 ± 1.39 ef |
| T3 | 0.54 ± 0.00 b | 0.39 ± 0.00 b | 0.27 ± 0.03 cd | 5.87 ± 0.54 d | 4.18 ± 0.39 b | 4.93 ± 0.42 de | 14.98 ± 0.32 d |
| T4 | 0.39 ± 0.03 f | 0.27 ± 0.02 ef | 0.21 ± 0.00 d | 4.68 ± 0.25 ef | 3.69 ± 0.16 bc | 4.35 ± 0.14 e | 12.72 ± 0.22 e |
| T5 | 0.41 ± 0.03 def | 0.43 ± 0.02 a | 0.30 ± 0.06 c | 6.37 ± 0.54 cd | 4.23 ± 2.13 b | 5.66 ± 1.15 cd | 16.27 ± 3.25 cd |
| T6 | 0.43 ± 0.00 d | 0.32 ± 0.01 d | 0.27 ± 0.01 cd | 10.86 ± 0.11 a | 6.60 ± 0.62 a | 7.22 ± 0.47 b | 24.68 ± 0.97 a |
| T7 | 0.52 ± 0.01 bc | 0.23 ± 0.01 h | 0.26 ± 0.01 cd | 7.50 ± 0.38 b | 2.79 ± 0.07 cd | 5.39 ± 0.40 cd | 15.69 ± 0.75 cd |
| T8 | 0.39 ± 0.03 ef | 0.24 ± 0.02 gh | 0.26 ± 0.00 cd | 3.21 ± 0.08 h | 2.74 ± 0.24 cd | 5.13 ± 0.09 cde | 11.08 ± 0.38 ef |
| T9 | 0.54 ± 0.03 b | 0.40 ± 0.01 b | 0.32 ± 0.03 c | 6.56 ± 0.17 c | 3.90 ± 0.29 b | 5.63 ± 0.41 cd | 16.09 ± 0.18 cd |
| T10 | 0.42 ± 0.01 de | 0.29 ± 0.02 e | 0.27 ± 0.04 c | 3.76 ± 0.31 g | 2.26 ± 0.09 de | 4.38 ± 0.31 e | 10.40 ± 0.38 fg |
| T11 | 0.71 ± 0.03 a | 0.35 ± 0.02 c | 0.42 ± 0.02 ab | 4.40 ± 0.08 f | 2.40 ± 0.32 de | 6.00 ± 0.50 c | 12.79 ± 0.79 e |
| T12 | 0.42 ± 0.00 de | 0.25 ± 0.00 fgh | 0.39 ± 0.03 ab | 6.45 ± 0.45 c | 2.78 ± 0.08 cd | 7.84 ± 0.57 b | 17.07 ± 0.48 bc |
| T13 | 0.49 ± 0.01 c | 0.29 ± 0.02 e | 0.44 ± 0.02 a | 5.12 ± 0.29 e | 4.06 ± 0.38 b | 9.60 ± 0.44 a | 18.78 ± 0.99 b |
| T14 | 0.38 ± 0.02 fg | 0.27 ± 0.03 efg | 0.27 ± 0.04 cd | 4.52 ± 0.18 f | 2.52 ± 0.33 de | 5.47 ± 0.86 cd | 12.52 ± 1.13 e |
| No. | P Allocation Rate (%) | K Allocation Rate (%) | ||||
|---|---|---|---|---|---|---|
| Root | Stem | Leaves | Root | Stem | Leaves | |
| T1 | 31.54 ± 2.41 ef 1 | 16.87 ± 1.24 f | 51.59 ± 1.61 a | 21.87 ± 2.35 d | 17.89 ± 1.25 e | 60.24 ± 3.57 b |
| T2 | 28.49 ± 1.45 f | 23.78 ± 2.69 bcd | 47.73 ± 2.11 b | 41.25 ± 4.98 a | 21.66 ± 2.49 d | 37.08 ± 5.42 f |
| T3 | 39.17 ± 2.81 c | 27.94 ± 3.08 ab | 32.89 ± 3.72 e | 40.43 ± 2.66 ab | 16.04 ± 3.67 f | 43.53 ± 4.21 e |
| T4 | 36.78 ± 1.77 cd | 28.98 ± 4.87 a | 34.24 ± 4.42 e | 30.77 ± 0.97 c | 19.34 ± 4.33 e | 49.89 ± 0.65 c |
| T5 | 40.07 ± 7.52 bc | 25.12 ± 5.47 abc | 34.80 ± 5.21 e | 28.17 ± 3.59 c | 13.49 ± 5.77 g | 58.34 ± 2.94 b |
| T6 | 44.05 ± 1.93 ab | 26.71 ± 6.53 ab | 29.24 ± 6.10 f | 27.60 ± 1.12 c | 27.87 ± 6.04 b | 44.54 ± 0.81 de |
| T7 | 47.81 ± 0.70 a | 17.84 ± 7.79 ef | 34.35 ± 7.28 e | 37.23 ± 0.73 b | 18.33 ± 7.53 e | 44.44 ± 1.25 de |
| T8 | 29.00 ± 0.27 f | 24.73 ± 8.31 bc | 46.27 ± 8.08 bc | 19.82 ± 0.93 d | 24.24 ± 8.83 c | 55.94 ± 1.48 b |
| T9 | 40.77 ± 1.43 bc | 24.25 ± 9.68 bcd | 34.98 ± 9.41 e | 27.63 ± 0.57 c | 23.14 ± 9.77 c | 49.23 ± 0.36 c |
| T10 | 36.18 ± 2.79 cde | 21.73 ± 10.58 cde | 42.09 ± 10.28 d | 20.51 ± 0.31 d | 13.97 ± 10.53 g | 65.53 ± 0.74 a |
| T11 | 34.46 ± 2.22 de | 18.69 ± 11.52 ef | 46.85 ± 11.08 bc | 15.37 ± 0.36 e | 18.86 ± 11.94 e | 65.77 ± 1.21 a |
| T12 | 37.77 ± 2.51 cd | 16.31 ± 12.39 f | 45.93 ± 12.88 bc | 30.86 ± 1.65 c | 19.04 ± 12.14 e | 50.10 ± 1.76 c |
| T13 | 27.27 ± 1.27 f | 21.60 ± 13.97 cde | 51.13 ± 13.83 a | 20.40 ± 1.43 d | 30.99 ± 13.90 a | 48.61 ± 1.72 cd |
| T14 | 36.26 ± 2.30 cd | 20.20 ± 14.38 def | 43.54 ± 14.07 cd | 28.72 ± 2.31 c | 13.61 ± 14.59 g | 57.67 ± 1.74 b |
| No. | K Content/g·kg−1 | K Accumulation/mg·plant−1 | |||||
|---|---|---|---|---|---|---|---|
| Root | Stem | Leaves | Root | Stem | Leaves | Total | |
| T1 | 2.06 ± 0.20 i 1 | 1.75 ± 0.16 f | 3.77 ± 0.72 d | 17.08 ± 2.25 g | 13.97 ± 1.46 i | 47.32 ± 8.22 i | 78.36 ± 10.72 i |
| T2 | 7.98 ± 0.06 a | 4.62 ± 0.42 a | 3.49 ± 0.87 de | 54.25 ± 4.84 d | 28.60 ± 2.35 e | 49.22 ± 10.23 hi | 132.07 ± 11.24 fg |
| T3 | 5.93 ± 0.39 b | 2.39 ± 0.09 e | 3.74 ± 0.61 d | 64.09 ± 4.99 b | 25.42 ± 3.69 efg | 68.95 ± 6.29 f | 158.46 ± 3.14 de |
| T4 | 2.92 ± 0.13 fg | 2.25 ± 0.01 e | 2.79 ± 0.02 f | 35.41 ± 1.19 f | 22.26 ± 4.48 fgh | 57.41 ± 1.22 gh | 115.07 ± 1.71 h |
| T5 | 4.00 ± 0.72 c | 2.21 ± 0.01 e | 6.78 ± 0.10 a | 62.42 ± 10.72 bc | 29.72 ± 5.03 de | 128.50 ± 0.10 b | 220.64 ± 10.78 b |
| T6 | 3.55 ± 0.08 de | 4.39 ± 0.04 a | 5.35 ± 0.24 b | 89.28 ± 1.79 a | 90.31 ± 6.74 a | 144.32 ± 9.28 a | 323.91 ± 16.26 a |
| T7 | 3.09 ± 0.01 fg | 1.83 ± 0.01 f | 2.62 ± 0.14 f | 44.83 ± 1.55 e | 22.06 ± 7.62 fgh | 53.57 ± 3.70 ghi | 120.47 ± 5.48 gh |
| T8 | 2.47 ± 0.17 hi | 2.24 ± 0.11 e | 3.06 ± 0.05 ef | 21.07 ± 1.70 g | 25.75 ± 8.59 ef | 59.36 ± 0.60 g | 91.79 ± 11.29 i |
| T9 | 3.32 ± 0.25 ef | 3.45 ± 0.19 c | 3.81 ± 0.28 cd | 40.26 ± 0.86 ef | 33.71 ± 9.06 cd | 71.74 ± 0.89 f | 145.80 ± 0.75 ef |
| T10 | 3.86 ± 0.04 cd | 2.95 ± 0.12 d | 6.80 ± 0.34 a | 34.33 ± 2.09 f | 23.37 ± 10.38 fg | 109.64 ± 4.97 c | 167.34 ± 7.97 cd |
| T11 | 2.76 ± 0.08 gh | 3.10 ± 0.20 d | 5.10 ± 0.25 b | 17.10 ± 1.38 g | 20.99 ± 11.14 gh | 73.02 ± 2.96 f | 111.10 ± 6.32 h |
| T12 | 3.66 ± 0.03 cde | 3.12 ± 0.02 d | 4.44 ± 0.13 c | 55.63 ± 3.61 cd | 34.32 ± 12.59 c | 90.25 ± 2.14 d | 180.19 ± 2.11 c |
| T13 | 3.30 ± 0.15 ef | 3.89 ± 0.21 b | 3.88 ± 0.09 cd | 35.92 ± 3.82 f | 54.48 ± 13.31 b | 85.36 ± 0.62 de | 175.76 ± 7.18 c |
| T14 | 3.24 ± 0.38 ef | 1.91 ± 0.03 f | 3.74 ± 0.09 d | 38.47 ± 4.55 ef | 18.19 ± 14.84 h | 77.10 ± 3.86 ef | 133.77 ± 7.29 fg |
| Number | Treatment | N/P | N/K | P/K |
|---|---|---|---|---|
| T1 | N0P0K0 | 3.44 ± 0.18 c 1 | 0.40 ± 0.05 ef | 0.12 ± 0.02 ab |
| T2 | N0P2K2 | 3.38 ± 0.04 c | 0.30 ± 0.04 h | 0.09 ± 0.01 cde |
| T3 | N1P2K2 | 4.22 ± 0.99 abc | 0.35 ± 0.03 g | 0.09 ± 0.02 cde |
| T4 | N2P0K2 | 4.93 ± 0.16 ab | 0.55 ± 0.01 a | 0.11 ± 0.00 abc |
| T5 | N2P1K2 | 4.10 ± 0.80 abc | 0.29 ± 0.02 h | 0.07 ± 0.01 def |
| T6 | N2P2K2 | 4.22 ± 1.58 abc | 0.25 ± 0.00 i | 0.07 ± 0.02 ef |
| T7 | N2P3K2 | 3.75 ± 0.13 bc | 0.49 ± 0.02 b | 0.13 ± 0.00 a |
| T8 | N2P2K0 | 4.91 ± 0.20 ab | 0.51 ± 0.01 ab | 0.10 ± 0.01 bc |
| T9 | N2P2K1 | 4.03 ± 0.14 abc | 0.44 ± 0.01 cd | 0.11 ± 0.00 abc |
| T10 | N2P2K3 | 5.16 ± 1.52 a | 0.27 ± 0.02 hi | 0.05 ± 0.02 f |
| T11 | N3P2K2 | 4.33 ± 0.97 abc | 0.43 ± 0.02 de | 0.10 ± 0.02 bc |
| T12 | N1P1K2 | 3.66 ± 0.07 bc | 0.35 ± 0.00 g | 0.09 ± 0.00 bcd |
| T13 | N1P2K1 | 3.96 ± 0.92 abc | 0.37 ± 0.01 fg | 0.10 ± 0.02 bcd |
| T14 | N2P1K1 | 5.14 ± 0.48 a | 0.48 ± 0.03 bc | 0.09 ± 0.00 bcd |
| Main Index | Correlation Coefficient | Direct Path Coefficient |
|---|---|---|
| Total N accumulation | 0.836 ** | 0.24 |
| Root N accumulation | 0.880 ** | 0.437 |
| Leaves N content | 0.028 | −0.233 |
| Total K accumulation | 0.837 ** | 0.806 |
| Root P accumulation | 0.875 ** | 0.282 |
| Stem K accumulation | 0.811 ** | −0.432 |
| Stem K content | 0.195 | −0.252 |
| Root P content | −0.239 | −0.061 |
| Stem P content | −0.045 | 0.068 |
| N/P | 0.047 | −0.077 |
| P/K | −0.252 | −0.097 |
| Main Index | Root | Stem | Leaves |
|---|---|---|---|
| Total dry biomass | 0.62 ** | 0.39 ** | −0.79 ** |
| Total N accumulation | 0.46 ** | 0.23 | −0.55 ** |
| Root N accumulation | 0.61 ** | 0.19 | −0.68 ** |
| Leaves N content | 0.09 | −0.26 * | 0.07 |
| Total K accumulation | 0.54 ** | 0.25 * | −0.64 ** |
| Stem K accumulation | 0.34 * | 0.44 ** | −0.56 ** |
| Stem K content | −0.04 | 0.00 | 0.04 |
| Root P accumulation | 0.72 ** | 0.19 | −0.77 ** |
| Root P content | −0.32 * | −0.12 | 0.36 * |
| Stem P content | −0.02 | −0.19 | 0.12 |
| N/P | −0.13 | −0.09 | 0.17 |
| N/K | −0.25 * | −0.08 | 0.27 * |
| P/K | −0.12 | 0.02 | 0.10 |
| Main Index | Rang Value | Fertilizer Effect Ordination | ||
|---|---|---|---|---|
| N | P | K | ||
| The total dry biomass | 4.54 | 3.17 | 3.97 | N > K > P |
| Root N accumulation | 6.89 | 5.23 | 6.03 | N > K > P |
| The total N accumulation | 42.58 | 28.06 | 37.72 | N > K > P |
| Root N allocation rate | 6.39 | 5.13 | 4.27 | N > P > K |
| Leaves N content | 4.63 | 5.43 | 6.54 | K > P > N |
| Root P accumulation | 7.51 | 6.18 | 7.65 | K > N > P |
| Stem P allocation rate | 9.25 | 11.14 | 4.98 | P > N > K |
| Root P content | 0.28 | 0.13 | 0.15 | N > K > P |
| Stem P content | 0.13 | 0.2 | 0.16 | P > K > N |
| Stem K accumulation | 69.32 | 68.05 | 66.94 | N > P > K |
| The total K accumulation | 212.81 | 208.84 | 232.12 | K > N > P |
| Stem K allocation rate | 11.83 | 14.38 | 13.9 | P > K > N |
| Stem K content | 2.23 | 2.56 | 2.15 | P > N > K |
| N/P | 0.95 | 1.18 | 1.13 | P > K > N |
| N/K | 0.18 | 0.3 | 0.26 | P > K > N |
| P/K | 0.03 | 0.06 | 0.06 | P = K > N |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.-J.; Wu, X.-H.; Chen, L.-H.; Huang, L.-M.; Chen, Y.; Wu, J.; El-Kassaby, Y.A.; Grossnickle, S.C.; Feng, J.-L. Fertilization Regulates Accumulation and Allocation of Biomass and Nutrients in Phoebe bournei Seedlings. Agriculture 2021, 11, 1187. https://doi.org/10.3390/agriculture11121187
Yang Z-J, Wu X-H, Chen L-H, Huang L-M, Chen Y, Wu J, El-Kassaby YA, Grossnickle SC, Feng J-L. Fertilization Regulates Accumulation and Allocation of Biomass and Nutrients in Phoebe bournei Seedlings. Agriculture. 2021; 11(12):1187. https://doi.org/10.3390/agriculture11121187
Chicago/Turabian StyleYang, Zhi-Jian, Xiao-Hui Wu, Lai-He Chen, Lan-Ming Huang, Yu Chen, Juan Wu, Yousry A. El-Kassaby, Steven C. Grossnickle, and Jin-Ling Feng. 2021. "Fertilization Regulates Accumulation and Allocation of Biomass and Nutrients in Phoebe bournei Seedlings" Agriculture 11, no. 12: 1187. https://doi.org/10.3390/agriculture11121187