
Content of Amino Acids in Maize and Yellow Lupine after Fluorine Application to Soil


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Design
2.2. Plants and Soil Analysis
2.3. Statistical Analysis
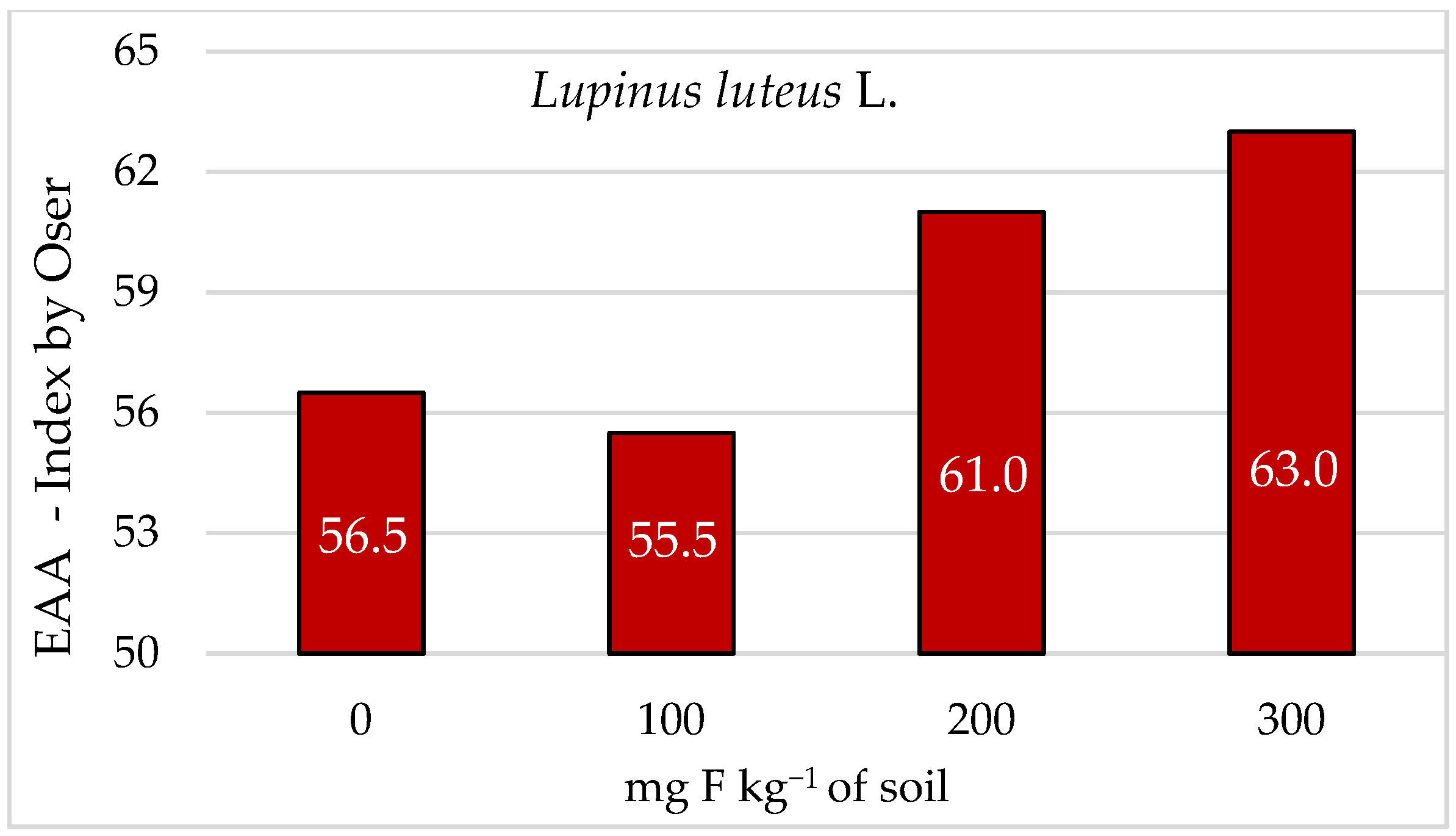
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Szostek, R.; Ciećko, Z. Content of fluoride in biomass of crops depending on soil contamination by this element. Fluoride 2014, 47, 294–306. [Google Scholar]
- Reddy, M.P.; Kaur, M. Sodium fluoride induced growth and metabolic changes in Salicornia Brachiata Roxb. Water Air Soil Pollut. 2008, 188, 171–179. [Google Scholar] [CrossRef]
- Smolik, B.; Pelc, J.; Śnioszek, M.; Smolik, M. Fluoride content of teas available on the polish market. Fluoride 2017, 50, 292–299. [Google Scholar]
- Gouider, M.; Feki, M.; Sayadi, S. Separative recovery with lime of phosphate and fluoride from an acidic effluent containing H3PO4, HF and/or H2SiF6. J. Hazard. Mater. 2009, 170, 962–968. [Google Scholar] [CrossRef]
- Zhu, S.; Zhang, J.; Dong, T. Removal of fluorine from contaminated field soil by anolyte enhanced electrokinetic remediation. Environ. Earth Sci. 2009, 59, 379–384. [Google Scholar] [CrossRef]
- Yu, L.; Zhang, J.; Du, C.; Yang, H.; Ye, B.-C. Distribution and pollution evaluation of fluoride in a soil-water-plant system in Shihezi, Xinjiang, China. Hum. Ecol. Risk Assess. 2018, 24, 445–455. [Google Scholar] [CrossRef]
- Szostek, R.; Ciećko, Z. Effect of soil contamination with fluorine on the yield and content of nitrogen forms in the biomass of crops. Environ. Sci. Pollut. Res. 2017, 24, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Szostek, R.; Ciećko, Z. The effect of soil contamination with fluorine on the contents of calcium and magnesium in the biomass of crop plants. Fluoride 2017, 50, 41–58. [Google Scholar]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2011; 505p. [Google Scholar]
- Telesiński, A.; Śnioszek, M. Bioindycators of environmental pollution with fluorine. Bromat. Chem. Toksykol. 2009, 62, 1148–1154. [Google Scholar]
- Gramowska, H.; Siepak, J. The effect of the fluorine level on the state of leaves and needles of trees in Poznań city and its vicinities. Annu. Set Environ. Protect. 2002, 4, 455–478. [Google Scholar]
- Fornasiero, B.R. Fluorides effects on Hypericum perforatum plants: First field observations. Plant Sci. 2003, 165, 507–513. [Google Scholar] [CrossRef]
- Rakowski, K. Physiological and molecular basis of trees’response to environmental stresses. Part II. The influence of hydrogen fluoride and low temperatures. For. Res. Pap. 2005, 2, 93–105. [Google Scholar]
- Pelc, J.; Smolik, B.; Krupa-Małkiewicz, M. Effect of sodium fluoride on some morphological and physiological parameters of 10-day-old seedlings of various plant species. Folia Pomeranae Univ. Technol. Stetin. Agric. Aliment. Piscaria Zootech. 2017, 3384, 151–158. [Google Scholar] [CrossRef]
- Pelc, J.; Śnioszek, M.; Wrobel, J.; Telesiński, A. Effect of Fluoride on Germination, Early Growth and Antioxidant Enzymes Activity of Three Winter Wheat (Triticum aestivum L.) Cultivars. Appl. Sci. 2020, 10, 6971. [Google Scholar] [CrossRef]
- Research Procedure No. 29. Research Procedure. Chemical-Agricultural Station Instruction, II ed.; Chemical-Agricultural Station: Warsaw, Poland, 2008.
- Lityński, T.; Jurkowska, H.; Gorlach, E. Chemical and Agricultural Analysis; PWN Publishing House: Warsaw, Poland, 1976; pp. 129–132. [Google Scholar]
- Shimadzu. Shimadzu Analytical and Measuring Instruments. User’s Manual; Shimadzu Corporation: Kyoto, Japan, 2016. [Google Scholar]
- Bremner, J.M. Total nitrogen. In Methods of Soil Analysis, Part 2. Chemical and Microbiological Properties (Agronomy 9); Black, C.A., Evans, D.D., Ensminger, L.E., White, J.L., Clark, F.E., Eds.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 1149–1178. [Google Scholar]
- Panak, H. Methodical Guide to Agricultural Chemistry Exercises; Agricultural and Technical Academy, ART Olsztyn Publishing House: Olsztyn, Poland, 1997; p. 242. (In Polish) [Google Scholar]
- Amino Acid Analyser AAA400, User Manual; Ingos S.R.O.: Prague, Czech Republic, 2007.
- Oser, B.L. An Integrated Essential Amino Acid Index for Predicting Biological Value of Proteins; Academic Press: New York, NY, USA, 1959; pp. 295–311. [Google Scholar]
- StatSoft, Inc. STATISTICA Data Analysis Software System; Version 12.0; Statsoft Inc.: Tulsa, OK, USA, 2012; Available online: www.statsoft.com (accessed on 23 October 2020).
- Li, C.H.; Ni, D. Effect of fluoride on the amino acid composition of tea leaves. Fluoride 2016, 49, 274–278. [Google Scholar]
- Li, T.; Yu, L.; Li, M.; Li, W. Comparative studies on the qualities of green teas in Karst and non-Karst areas of Yichang, Hubei Province, PR China. Food Chem. 2007, 103, 71–74. [Google Scholar] [CrossRef]
- Cai, H.; Dong, Y.; Li, Y.; Li, D.; Peng, C.; Zhang, Z.; Wan, X. Physiological and cellular responses to fluoride stress in tea (Camellia sinensis) leaves. Acta Physiol. Plant. 2016, 38, 144–155. [Google Scholar] [CrossRef]
- Das, C.; Dey, U.; Chakraborty, D.; Datta, J.K.; Mondal, N.K. Fluoride toxicity effects in potato plant (Solanum tuberosum L.) grown in contaminated soils. Octa J. Environ. Res. 2015, 3, 136–143. [Google Scholar]
- Zouari, M.; Ahmed, C.B.; Elloumi, N.; Rouina, B.B.; Labrousse, P.; Abdallah, F.B. Effects of irrigation water fluoride on relative water content, photosynthetic activity, and proline accumulation in young olive trees (Olea europaea L. cv. chemlali) in Arid Zones. Fluoride 2016, 49, 303–372. [Google Scholar]
- Saleh, A.A.H.; Abdel-Kader, D.Z. Metabolic responses of two Helianthus annuus cultivars to different fluoride concentrations during germination and seedling growth stages. Egypt. J. Exp. Biol. 2003, 5, 43–54. [Google Scholar]
- Ahmed, S.; Khalid, K.; Jabeen, F.; Ahmad, M.N.; Zia, A.; Haider, A.; Mujahid, M.; Zia, D.; Khan, N.P. The effects of fluoride stress on okra (Abelmoschus esculentus L.). Fluoride 2019, 52, 354–361. [Google Scholar]
- Mezghani, I.; Zouari, M.; Rouina, B.B.; Abdallah, F.B. Mulberry leaves as a bioindicator of fluoride pollution in the vicinity of a phosphate fertilizer factory located in Sfax, Tunisia. Fluoride 2019, 52, 537–545. [Google Scholar]
- Singh, M.; Singh, V.P.; Dubey, G.; Prasad, S.M. Exogenous proline application ameliorates toxic effects of arsenate in Solanum melongena L. seedlings. Ecotoxicol. Environ. Saf. 2015, 117, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Gadi, B.R.; Verma, P.; Amra, R. Influence of NaF on seed germination, membrane stability and some biochemical content in Vigna seedlings. J. Chem. Biol. Phys. Sci. 2012, 2, 1371–1378. [Google Scholar]
- Datta, J.K.; Maitra, A.; Mondal, N.K.; Banerjee, A. Studies on the impact of fluoride toxicity on germination and seedling growth of gram seed (Cicer arietinum L. cv. Anuradha). J. Stress Physiol. Biochem. 2012, 8, 194–202. [Google Scholar]
- Maitra, A.; Datta, J.K.; Mondal, N.K. Amelioration of fluoride toxicity with the use of indigenous inputs. J. Stress Physiol. Biochem. 2013, 9, 207–219. [Google Scholar]
- Eyini, M.; Sujanandini, K.; Pothiraj, C.; Jayakumar, M.; Kil, B.-S. Differental response of Azolla microphylla Kaulf. and Azolla filiculoides Lam. to sodium fluoride. J. Plant Biol. 1999, 42, 299–301. [Google Scholar] [CrossRef]
- Greenway, H.; Munns, R. Mechanism of salt tolerance in non-halophytes. Annu. Rev. Plant Physiol. 1980, 31, 149–190. [Google Scholar] [CrossRef]
- Hanson, A.D.; Nelson, C.E.; Everson, E.H. Evaluation of free proline accumulation as an index of drought resistance using, two contrasting barley cultivars. Crop Sci. 1977, 17, 720–726. [Google Scholar] [CrossRef]
- Singh, U.P.; Rana Yashu, B.; Kumar, S.; Striastava, J.P. Biochemical responses of elevated level of fluoride in nutrient medium on wheat and barley. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 3116–3123. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Patra, K.P. Biochemical and antioxidant responses of paddy (Oryza sativa L.) to fluoride stress. Fluoride 2015, 48, 56–61. [Google Scholar]
- Pal, C.K.; Mondal, K.N.; Bhaumik, R.; Banerjee, A.; Datta, K.J. Incorporation of fluoride in vegetation and associated biochemical changes due to fluoride contamination in water and soil: A comparative field study. Ann. Environ. Sci. 2012, 6, 123–139. [Google Scholar]
- Kabata-Pendias, A.; Pendias, H. Trace Elements in Soils and Plants, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2001; 403p. [Google Scholar]
- FAO. World Food and Agriculture—Statistical Yearbook; FAO: Rome, Italy, 2021; p. 368. [Google Scholar] [CrossRef]
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World. Transforming Food Systems for Affordable Healthy Diets; FAO: Rome, Italy, 2020; p. 320. [Google Scholar] [CrossRef]
- FAO. The State of the World’s Biodiversity for Food and Agriculture; Bélanger, J., Pilling, D., Eds.; FAO Commission on Genetic Resources for Food and Agriculture Assessments: Rome, Italy, 2019; 572p, Available online: http://www.fao.org/3/CA3129EN/CA3129EN.pdf (accessed on 7 November 2021).
- OECD/FAO. OECD-FAO Agricultural Outlook 2021–2030; OECD Publishing: Paris, France, 2021; 337p. [Google Scholar] [CrossRef]



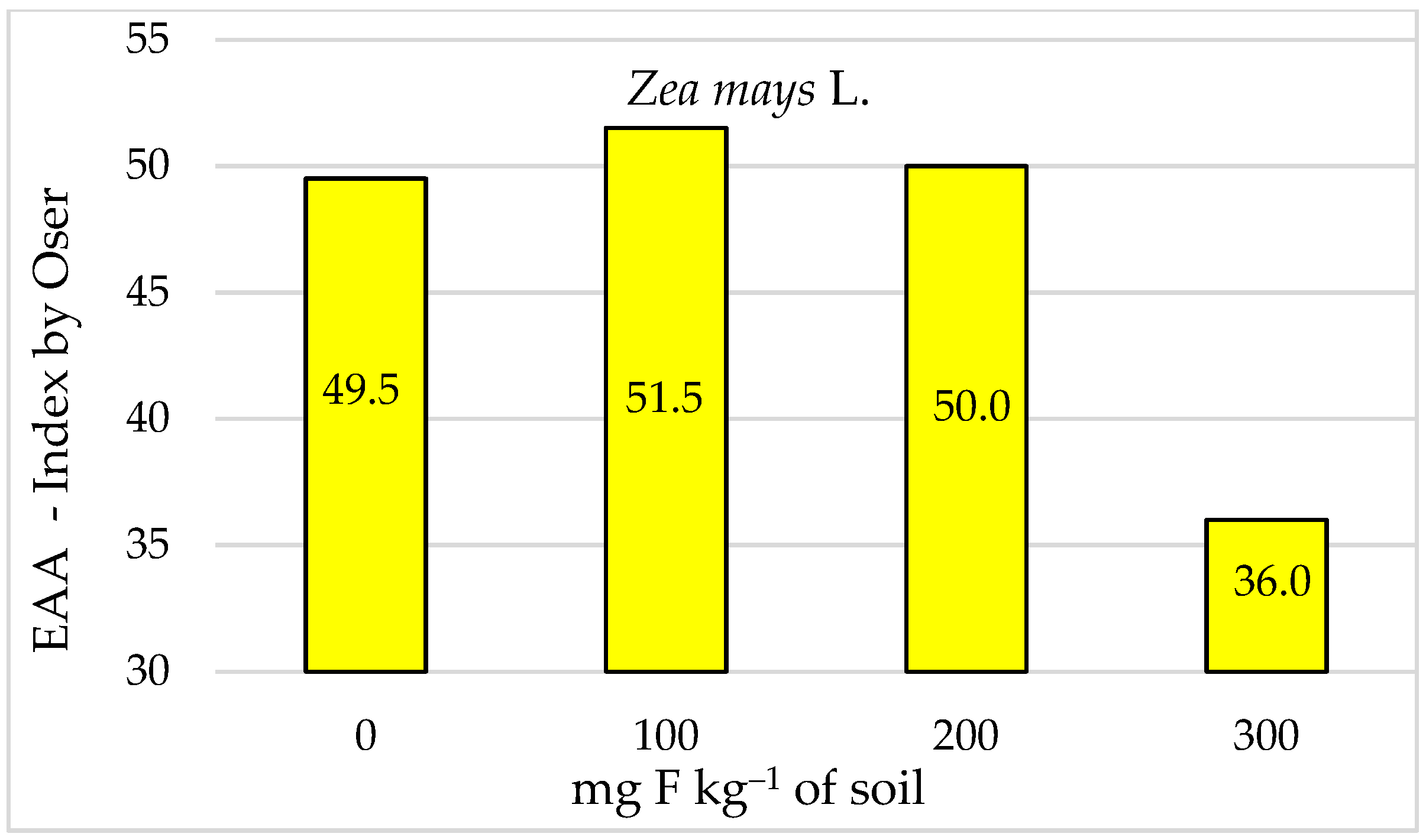{kind=link}
{kind=link}
| Amino Acids | Soil Contamination with Fluorine in mg F kg−1 of Soil | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 100 | 200 | 300 | r | 0 | 100 | 200 | 300 | r | |
| in g 100 g−1 Total Protein (16 g N) | in g kg−1 DM | |||||||||
| Exogenous | ||||||||||
| Arginine | 2.87 | 3.55 | 3.61 | 2.45 | −0.28 | 1.20 | 1.64 | 1.67 | 1.12 | −0.09 |
| Phenylalanine | 3.41 | 3.07 | 3.63 | 2.65 | −0.52 | 1.43 | 1.42 | 1.68 | 1.21 | −0.27 |
| Histidine | 1.65 | 1.64 | 1.90 | 1.29 | −0.42 | 0.69 | 0.76 | 0.88 | 0.59 | −0.19 |
| Isoleucine | 2.72 | 2.94 | 2.88 | 2.13 | −0.64 * | 1.14 | 1.36 | 1.33 | 0.97 | −0.38 |
| Leucine | 5.99 | 6.29 | 6.46 | 4.69 | −0.60 * | 2.51 | 2.91 | 2.99 | 2.14 | −0.34 |
| Lysine | 2.53 | 2.59 | 2.62 | 1.82 | −0.71 ** | 1.06 | 1.20 | 1.21 | 0.83 | −0.50 |
| Methionine | 0.91 | 0.99 | 0.61 | 0.39 | −0.90 ** | 0.38 | 0.46 | 0.28 | 0.18 | −0.83 ** |
| Threonine | 2.79 | 3.55 | 3.68 | 2.32 | −0.26 | 1.17 | 1.64 | 1.70 | 1.06 | −0.11 |
| Tyrosine | 1.96 | 1.51 | 1.47 | 1.32 | −0.92 ** | 0.82 | 0.70 | 0.68 | 0.60 | −0.97 ** |
| Valine | 3.96 | 4.26 | 4.32 | 3.07 | −0.58 * | 1.66 | 1.97 | 2.00 | 1.40 | −0.37 |
| Sum of exogenous amino acids | 28.80 | 30.40 | 31.18 | 22.14 | - | 12.06 | 14.06 | 14.42 | 10.10 | - |
| LSD0.05 | 2.67 | 1.20 | ||||||||
| Endogenous | ||||||||||
| Alanine | 4.94 | 5.23 | 5.17 | 3.75 | −0.68 * | 2.07 | 2.42 | 2.39 | 1.71 | −0.43 |
| Cysteine | 0.02 | 0.04 | 0.07 | 0.02 | 0.16 | 0.01 | 0.02 | 0.03 | 0.01 | 0.13 |
| Glycine | 3.92 | 4.15 | 4.28 | 3.16 | −0.55 | 1.64 | 1.92 | 1.98 | 1.44 | −0.28 |
| Asparagine acid | 6.95 | 7.70 | 7.52 | 5.41 | −0.60 * | 2.91 | 3.56 | 3.48 | 2.47 | −0.35 |
| Glutamic acid | 7.57 | 7.81 | 8.43 | 4.60 | −0.63 * | 3.17 | 3.61 | 3.90 | 2.10 | −0.48 |
| Proline | 4.18 | 4.67 | 4.22 | 3.81 | −0.57 * | 1.75 | 2.16 | 1.95 | 1.74 | −0.16 |
| Serine | 3.32 | 3.74 | 3.81 | 2.48 | −0.52 | 1.39 | 1.73 | 1.76 | 1.13 | −0.32 |
| Sum of endogenous amino acids | 30.90 | 33.34 | 33.49 | 23.23 | - | 12.94 | 15.42 | 15.49 | 10.60 | - |
| LSD0.05 | 2.87 | 1.30 | ||||||||
| Sum of total amino acids | 59.70 | 63.74 | 64.67 | 45.37 | - | 25.00 | 29.48 | 29.91 | 20.70 | - |
| LSD0.05 | 5.54 | 2.49 | ||||||||
| Amino Acids | Soil Contamination with Fluorine in mg F kg−1 of Soil (Residual Effect) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 100 | 200 | 300 | r | 0 | 100 | 200 | 300 | r | |
| in g 100 g−1 Total Protein (16 g N) | in g kg−1 DM | |||||||||
| Exogenous | ||||||||||
| Arginine | 4.30 | 4.62 | 4.80 | 4.76 | 0.89 ** | 10.40 | 11.95 | 10.65 | 10.53 | −0.16 |
| Phenylalanine | 3.39 | 3.04 | 3.30 | 3.29 | −0.03 | 8.21 | 7.87 | 7.33 | 7.28 | −0.96 ** |
| Histidine | 2.31 | 2.15 | 2.47 | 2.39 | 0.53 | 5.58 | 5.56 | 5.48 | 5.29 | −0.93 ** |
| Isoleucine | 2.89 | 2.69 | 3.15 | 3.26 | 0.79 ** | 6.98 | 6.97 | 6.98 | 7.21 | 0.77 ** |
| Leucine | 5.75 | 5.38 | 6.32 | 6.86 | 0.85 ** | 13.90 | 13.93 | 14.03 | 15.18 | 0.83 ** |
| Lysine | 4.33 | 3.88 | 4.40 | 4.41 | 0.39 | 10.48 | 10.04 | 9.76 | 9.76 | −0.93 ** |
| Methionine | 0.46 | 0.48 | 0.65 | 0.85 | 0.95 ** | 1.11 | 1.24 | 1.44 | 1.87 | 0.96 ** |
| Threonine | 3.60 | 3.42 | 4.02 | 4.10 | 0.83 ** | 8.71 | 8.86 | 8.93 | 9.08 | 0.99 ** |
| Tyrosine | 3.27 | 2.80 | 3.02 | 2.89 | −0.58 * | 7.90 | 7.25 | 6.69 | 6.39 | −0.99 ** |
| Valine | 3.86 | 3.64 | 4.26 | 4.52 | 0.85 ** | 9.34 | 9.43 | 9.45 | 10.00 | 0.86 ** |
| Sum of exogenous amino acids | 34.15 | 32.12 | 36.39 | 37.33 | - | 82.61 | 83.10 | 80.74 | 82.59 | - |
| LSD0.05 | 3.30 | n.s. | ||||||||
| Endogenous | ||||||||||
| Alanine | 3.73 | 3.51 | 4.44 | 4.58 | 0.86 ** | 9.02 | 9.08 | 9.86 | 10.13 | 0.95 ** |
| Cysteine | <0.01 | <0.01 | <0.01 | <0.01 | 0.00 | 0.01 | 0.01 | 0.01 | 0.01 | 0.00 |
| Glycine | 3.71 | 3.49 | 4.05 | 4.00 | 0.70 * | 8.97 | 9.02 | 8.98 | 8.84 | −0.71 ** |
| Asparagine acid | 17.93 | 16.87 | 14.96 | 13.55 | −0.99 ** | 43.38 | 43.65 | 33.19 | 29.98 | −0.93 ** |
| Glutamic acid | 8.05 | 7.52 | 8.83 | 8.52 | 0.61 * | 19.48 | 19.45 | 19.6 | 18.85 | −0.67 * |
| Proline | 4.57 | 4.46 | 5.33 | 5.15 | 0.79 ** | 11.05 | 11.54 | 11.83 | 11.40 | 0.53 |
| Serine | 3.93 | 3.26 | 3.79 | 3.79 | 0.05 | 9.50 | 8.43 | 8.42 | 8.39 | −0.79 ** |
| Sum of endogenous amino acids | 41.93 | 39.10 | 41.42 | 39.59 | - | 101.40 | 101.10 | 91.89 | 87.60 | - |
| LSD0.05 | n.s. | 9.01 | ||||||||
| Sum of total amino aids | 76.08 | 71.22 | 77.81 | 76.92 | - | 184.00 | 184.20 | 172.60 | 170.10 | - |
| LSD0.05 | n.s. | n.s. | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szostek, R.; Ciećko, Z.; Rolka, E.; Wyszkowski, M. Content of Amino Acids in Maize and Yellow Lupine after Fluorine Application to Soil. Agriculture 2021, 11, 1120. https://doi.org/10.3390/agriculture11111120
Szostek R, Ciećko Z, Rolka E, Wyszkowski M. Content of Amino Acids in Maize and Yellow Lupine after Fluorine Application to Soil. Agriculture. 2021; 11(11):1120. https://doi.org/10.3390/agriculture11111120
Chicago/Turabian StyleSzostek, Radosław, Zdzisław Ciećko, Elżbieta Rolka, and Mirosław Wyszkowski. 2021. "Content of Amino Acids in Maize and Yellow Lupine after Fluorine Application to Soil" Agriculture 11, no. 11: 1120. https://doi.org/10.3390/agriculture11111120