
An Agronomic Approach to Pine Nut Production by Grafting Stone Pine on Two Rootstocks

Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Test and Plant Material
2.2. Growing and Production Measurements
2.3. Statistical Analysis
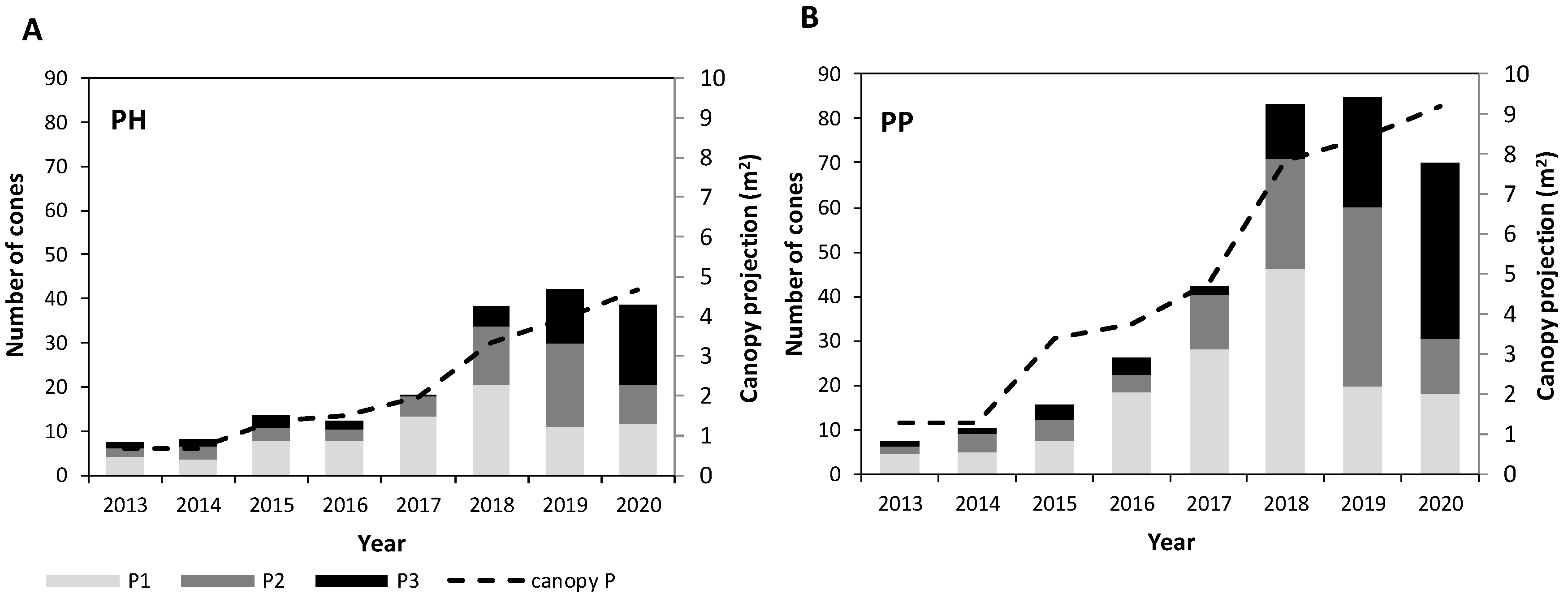
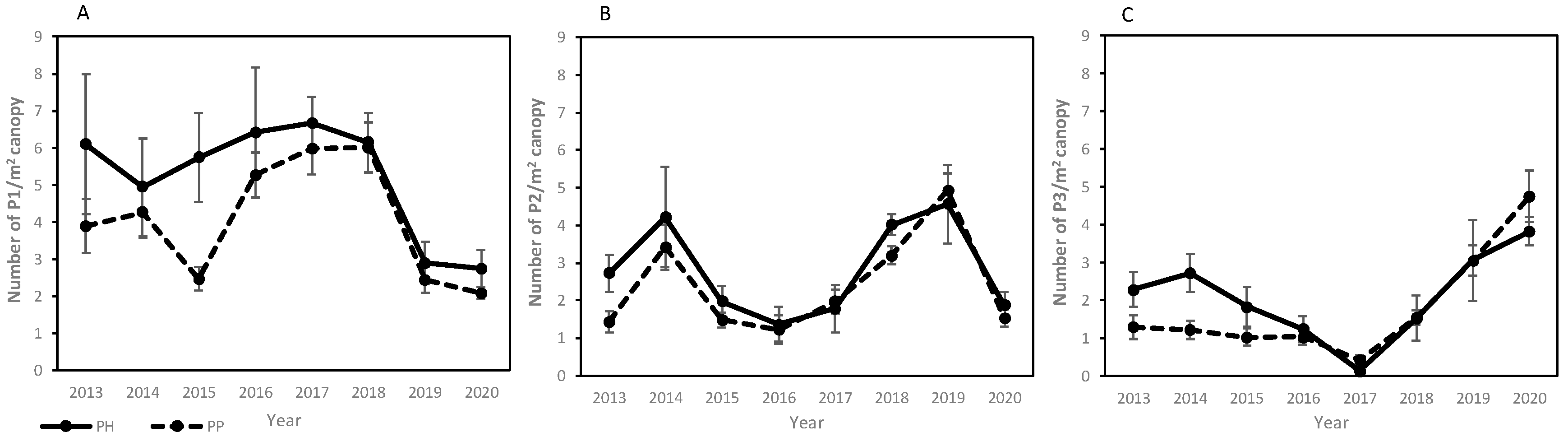
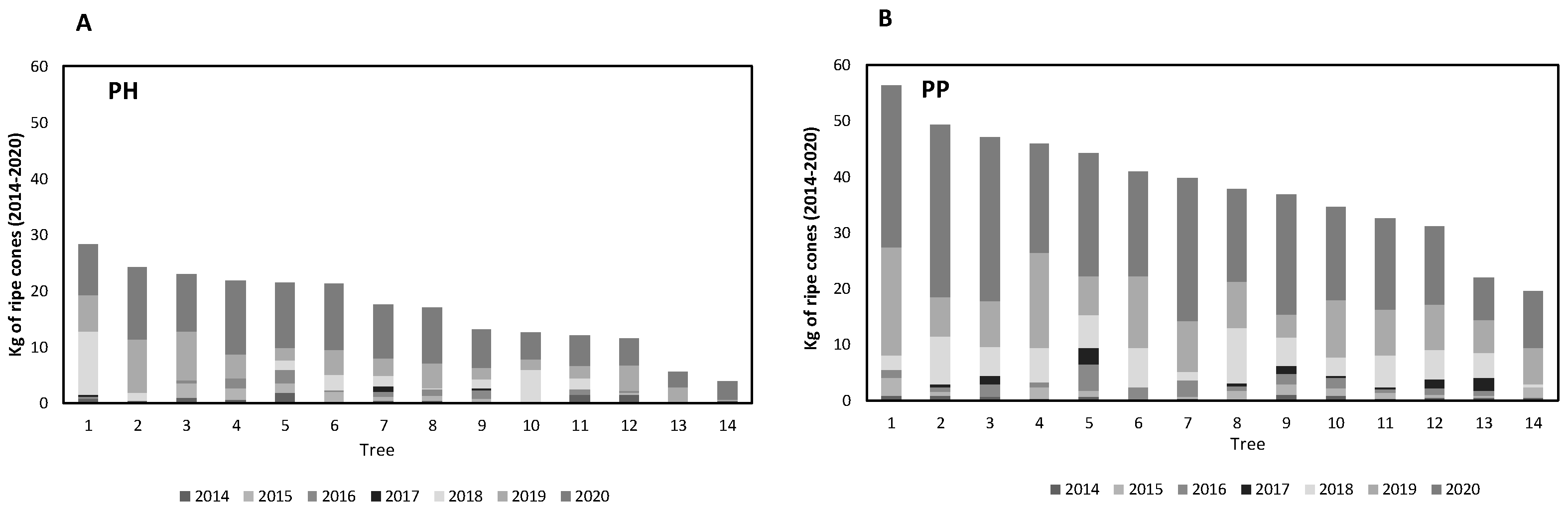
3. Results
3.1. Evaluation of Cone Production
3.2. Growth vs. Cone Production
3.3. Heterogeneity of the Harvest
3.4. Pine Nut Composition
4. Discussion
4.1. Productivity and Growth
4.2. Quality of the Harvest
4.2.1. Cone Production
4.2.2. Pine Nut Composition
4.3. New Stone Pine Orchards
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mutke, S.; Calama, R.; González-Martínez, S.; Montero, G.; Gordo, F.J.; Bono, D.; Gil, L. Mediterranean Stone Pine: Botany and Horticulture. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2012; Volume 39, pp. 153–201. [Google Scholar]
- Vendramin, G.G.; Fady, B.; González-Martínez, S.C.; Hu, F.S.; Scotti, I.; Sebastiani, F.; Soto, Á.; Petit, R.J. Genetically Depauperate but Widespread: The Case of an Emblematic Mediterranean Pine. Evolution 2008, 62, 680–688. [Google Scholar] [CrossRef]
- Mutke, S.; Vendramin, G.G.; Fady, B.; Bagnoli, F.; González-Martínez, S.C. Molecular and Quantitative Genetics of Stone Pine (Pinus pinea). In Genetic Diversity in Horticultural Plants; Springer: Cham, Switzerland, 2019; Volume 22. [Google Scholar]
- Mutke, S.; Gordo, J.; Chambel, M.R.; Prada, M.A.; Álvarez, D.; Iglesias, S.; Gil, L. Phenotypic plasticity is tronger than adaptative differentitation among Mediterranean stone pine provenances. For. Syst. 2010, 19, 354–366. [Google Scholar]
- Loewe-Muñoz, V.; Del Río, R.; Delard, C.; Balzarini, M. Enhancing Pinus pinea cone production by grafting in a non-native habitat. New For. 2021. [Google Scholar] [CrossRef]
- Evaristo, I.; Batista, D.; Correia, I.; Correia, P.; Costa, R. Chemical profiling of Portuguese Pinus pinea L. nuts and comparative analysis with Pinus koraiensis Sieb. & Zucc. comercial kernels. Options Mediterr. Mediterr. Stone Pine Agrofor. 2013, 105, 99–104. [Google Scholar]
- Nutfruit. Pine Nuts-Statistical Review. Nutfruit Magazine, March 2021; p. 1.
- Calama, R.; Gordo, J.; Mutke, S.; Conde, M.; Madrigal, G.; Garriga, E.; Arias, M.J.; Piqué, M.; Gandía, R.; Montero, G.; et al. Decline in commercial pine nut and kernel yield in Mediterranean stone pine (Pinus pinea L.) in Spain. iForest-Biogeosciences For. 2020, 13, 251–260. [Google Scholar] [CrossRef]
- Pimpão, M.; Valdiviesso, T.; Trindade, C.S.; Naves, P.; Sousa, E. Leptoglossus occidentalis damages on stone pine female reproductive structures. Options Mediterr. Mediterr. Stone Pine Agrofor. 2017, 122, 5. [Google Scholar]
- De la Mata, R.; Teixidó, A.; Aletà, N.; Torrell, A.; Ros, L.; Segarra, M. La producció de pinyons en plantacions empeltades. In XXXVI Jornades Tècniques Silvícoles Emili Garolera; Consorci Forestal de Catalunya: Girona, Spain, 2019; p. 86. [Google Scholar]
- Farinha, A.O.; Carvalho, C.; Correia, A.C.; Branco, M. Impact assessment of Leptoglossus occidentalis in Pinus pinea: Integrating population density and seed loss. For. Ecol. Manag. 2021, 496, 119422. [Google Scholar] [CrossRef]
- INC. Global statistical review. Crop progress report. Pine nuts. Nutfruit Magazine, March 2021; p. 4. [Google Scholar]
- Loewe-Muñoz, V.; Noel, D. Mediterranean Pinus pinea L. nuts from Southern Hemisphere provenances. Rend. Lincei Sci. Fis. Nat. 2021, 32, 181–189. [Google Scholar] [CrossRef]
- Gordo, F.J.; Calama, R.; Rojo, L.I.; Madrigal, G.; Álvarez, D.; Mutke, S.; Montero, G.; Finat, L. Experiencias de clareos en masas de Pinus pinea L. en la Mesta Norte. In Proceedings of the 5º Congreso Forestal Español, Ávila, Spain, 21–25 September 2009. [Google Scholar]
- Montero, G.; Calama, R.; Ruiz-Peinado, R. Selvicultura de Pinus pinea. In Compendio de Selvicultura Aplicada en España; Montero, G., Reque, J., Eds.; INIA-FUCOVASA: Serrada, Spain, 2008. [Google Scholar]
- Loewe, V.; Delard, C. Un Nuevo Cultivo Para Chile. El Pino Piñonero (Pinus pinea L.); Instituto Forestal-Chile: Santiago, Chile, 2012. [Google Scholar]
- Bono, D.; Aletà, N. Cone yield evaluation of a grafted Pinus pinea L. trial. Options Mediterr. A Mediterr. Stone Pine Agrofor. 2013, 105, 35–41. [Google Scholar]
- Guadaño, C.; Mutke, S. Establecimiento de Plantaciones Clonales de Pinus Pinea Para la Produccción de Piñón Mediterráneo; INIA: Madrid, Spain, 2016; Volume 28. [Google Scholar]
- Mutke, S.; Iglesias, S.; Gil, L. Selección de clones de pino piñonero sobresalientes en la producción de piña. Investig. Agrar. Sist. Recur. For. 2007, 16, 13. [Google Scholar]
- Catalán Bachiller, G. Plantaciones de Pinus pinea en zonas calizas para producción precoz de piñón. Ecología 1990, 4, 105–120. [Google Scholar]
- Gordo, F.J.; Mutke, S.; Gil, L. La relevancia de la especie de patrón portainjerto para el desarrollo de la copa y la producción de piña del pino piñonero injertado. In Proceedings of the 6º Congreso Forestal Español, Vitoria-Gasteiz, Spain, 10–14 June 2013. [Google Scholar]
- Piqué, M.; Coello, J.; Ammari, Y.; Aletà, N.; Sghaier, T.; Mutke, S. Grafted stone pine plantations for cone production: Trials on Pinus pinea and Pinus halepensis rootstocks from Tunisia and Spain. Options Mediterr. Ser. A Mediterr. Semin. 2017, 122, 17–23. [Google Scholar]
- García del Barrio, J.M.; de Miguel, J.; Alía, R.; Iglesias, S. Regiones de Identificación y Utilización del Material Forestal de Reproducción; Ministerio de Medio Ambiente: Madrid, Spain, 2001.
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Loewe-Muñoz, V.; Balzarini, M.; Álvarez-Contreras, A.; Delard-Rodríguez, C.; Navarro-Cerrillo, R.M. Fruit productivity of Stone pine (Pinus pinea L.) along a climatic gradient in Chile. Agric. For. Meteorol. 2016, 223, 203–216. [Google Scholar] [CrossRef]
- Del Rio, M.; Montero, G.; Calama, R. Selvicultura de Pinus halepensis. In Compendio de Selvicultura Aplicada en España; Montero, G., Reque, J., Eds.; INIA-FUCOVASA: Serrada, Spain, 2008. [Google Scholar]
- Cuesta, B.; Vega, J.; Villar-Salvador, P.; Rey-Benayas, J.M. Root growth dynamics of Aleppo pine (Pinus halepensis Mill.) seedlings in relation to shoot elongation, plant size and tissue nitrogen concentration. Trees 2010, 24, 899–908. [Google Scholar] [CrossRef]
- Shestakova, T.A.; Mutke, S.; Gordo, J.; Camarero, J.J.; Sin, E.; Pemán, J.; Voltas, J. Weather as main driver for masting and stem growth variation in stone pine supports compatible timber and nut co-production. Agric. For. Meteorol. 2021, 298-299. [Google Scholar] [CrossRef]
- Mutke, S.; Gordo, J.; Climent, J.; Gil, L. Shoot growth and phenology modelling of grafted stone pine (Pinus pinea L.) in Inner Spain. Ann. For. Sci. 2003, 60, 527–537. [Google Scholar] [CrossRef] [Green Version]
- Calama, R.; Mutke, S.; Tomé, J.; Gordo, J.; Montero, G.; Tomé, M. Modelling spatial and temporal variability in a zero-inflated variable: The case of stone pine (Pinus pinea L.) cone production. Ecol. Model. 2011, 222, 606–618. [Google Scholar] [CrossRef]
- Loewe-Muñoz, V.; Balzarini, M.; Delard, C.; Del Río, R.; Álvarez, A. Inter-annual variability of Pinus pinea L. cone productivity in a non-native habitat. New For. 2020, 51, 1055–1068. [Google Scholar] [CrossRef]
- Calama, R.; Mutke, S.; Pardos, M.; Morales, L.; Gordo, F.J.; Montero, G.; Finat, L. La producción de piña y piñón de Pinus pinea L. en la Meseta Norte. In La regeneración Natural de los Pinares en los Arenales de la Meseta Castellana; Gordo, F.J., Calama, R., Pardos, M., Bravo, F., Montero, G., Eds.; Instituto Universitario de Investigación en Gestión Forestal Sostenible (Universidad de Valladolid-INIA): Valladolid, Spain, 2012; p. 254. [Google Scholar]
- Climent, J.; Prada, M.A.; Gil, L.; Pardos, M. Increase of flowering in Pinus nigra Arn subsp salzamannii (Dunal) Franco by means of heteroplastic grafts. Ann. For. Sci. 1997, 54, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Afonso, A.; Gonçalves, A.C.; Pereira, D.G. Pinus pinea (L.) nut and kernel productivity in relation to cone, tree and stand characteristics. Agrofor. Syst. 2020, 94, 2065–2079. [Google Scholar] [CrossRef]
- Mutke, S.; Gordo, J.; Gil, L. Variability of Mediterranean Stone pine cone production: Yield loss as response to climate change. Agric. For. Meteorol. 2005, 132, 263–272. [Google Scholar] [CrossRef]
- Bracalini, M.; Benedettelli, S.; Croci, F.; Terreni, P.; Tiberi, R.; Panzavolta, T. Cone and Seed Pests of Pinus pinea: Assessment and Characterization of Damage. J. Econ. Entomol. 2013, 106, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Nasri, N.; Khaldi, A.; Fady, B.; Triki, S. Fatty acids from seeds of Pinus pinea L.: Composition and population profiling. Phytochemistry 2005, 66, 1729–1735. [Google Scholar] [CrossRef] [PubMed]
- González, M.; Loewe, V.; Delard, C. El piñón de Pinus pinea producido en Chile: Características nutricionales y organolépticas. Vitae 2012, 19, S317–S319. [Google Scholar]
- Tukan, S.K.; Al-Ismail, K.; Ajo, R.Y.; Al-Dabbas, M.M. Seeds and seed oil compositions of Aleppo Pine (Pinus halepensis Mill.) grown in Jordan. Riv. Ital. Sostanze Grasse 2013, 90, 87–93. [Google Scholar]
- Piqué, M. Modelos de Producción Para las Masas de Pinus pinea L. en Catalunya; Universitat de Lleida: Lleida, Spain, 2003. [Google Scholar]
- Mutke, S.; Guadaño, C.; Iglesias, S.; León, D.; Arribas, S.; Gordo, F.J.; Gil, L.; Montero, G. Mediterranean pine nuts from forests and plantations. Selection and identification of Spanish elite clones for Mediterranean pine nut as orchard crop. Options Mediterr. 2017, 122, 71–75. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Tree Age (Year) | Rootstock | Total Number Per Tree | Weight Per Tree (g) | Weight Per Cone (g) | Ratios | ||||
|---|---|---|---|---|---|---|---|---|---|
| P1 | P2 | P3 | FW-P | W-EPN | FW-C | Sef | Yef | ||
| 5 (2014) | PP | 5.1 ± 2.9 | 3.9 ± 2 | 1.4 ± 0.8 | 577 ± 245 | 107 ± 59.6 | 377 ± 70.1 | 82 ± 14.7 | 17.6 ± 3.9 |
| PH | 3.5 ± 0.9 | 3.0 ± 1.0 | 1.7 ± 0.3 | 790 ± 138 | 135 ± 31.8 | 423 ± 22.7 | 76.6 ± 4.7 | 15.2 ± 1.5 | |
| 6 (2015) | PP | 7.6 ± 2.7 | 4.6 ± 2.3 | 3.3 ± 2.4 | 1343 ± 849 | 272 ± 165 | 350 ± 102 | 92.7 ± 7.5 | 20.5 ± 2.4 A |
| PH | 7.9 ± 1.9 | 2.9 ± 0.6 | 2.9 ± 0.9 | 1209 ± 223 | 217.5 ± 48.9 | 277 ± 20.9 | 87.1 ± 2.9 | 16.8 ± 1 B | |
| 7 (2016) | PP | 18.4 ± 7 A | 4.1 ± 4 | 3.6 ± 2.6 | 1728 ± 1128 | 286 ± 209 A | 435 ± 77.4 | 85.5 ± 8.8 | 16.1 ± 2.1 |
| PH | 7.9 ± 1.8 B | 2.5 ± 0.9 | 2.1 ± 0.6 | 1020 ± 181 | 120.1 ± 31.6 B | 375 ± 22.2 | 71 ± 3.7 | 11.4 ± 1 | |
| 8 (2017) 1 | PP | 28 ± 12.1 A | 12.4 ± 4.8A | 1.9 ± 2.2 A | 1311 ± 984 | 224 ± 199 A | 415 ± 111 | 89 ± 6.9 | 15.2 ± 3.3 |
| PH | 13.4 ± 1.9 B | 4.5 ± 1.0 B | 0.3 ± 0.2 B | 500 ± 97.2 | 66.9 ± 13.7 B | 346 ± 26.4 | 84.5 ± 1.8 | 13.3 ± 0.8 | |
| 9 (2018) | PP | 46.1 ± 17.6 A | 25.2 ± 6.4 A | 12.3 ± 6.2 A | 5077 ± 2517 | 1024 ± 566 A | 428 ± 67.4 | 93.1 ± 7.2 | 19.4 ± 2.6 A |
| PH | 20.4 ± 2.9 B | 13.5 ± 1.2 B | 4.5 ± 1.7 B | 2861 ± 884 | 337 ± 103 B | 465 ± 28.3 | 80.2 ± 7.8 | 14.6 ± 1.4 B | |
| 10 (2019) 2 | PP | 19.8 ± 5.4 A | 40.3 ± 11.6 A | 24.6 ± 12.9 A | 9384 ± 4264 A | 1377 ± 489 A | 426 ± 59.1 | 74.9 ± 11.3 | 15.2 ± 2.3 |
| PH | 11.1 ± 1.3 B | 18.7 ± 2.3 B | 12.4 ± 2 B | 4041 ± 717 B | 354.1 ± 42 B | 418 ± 21.7 | |||
| 11 (2020) | PP | 18.1 ± 7.4 A | 12.2 ± 5.2 A | 39.7 ± 12 A | 19840 ± 7015 A | 3669 ± 1335 A | 505 ± 84.6 | 89.2 ± 8.5 | 18.5 ± 2.1 |
| PH | 11.8 ± 1.7 B | 8.6 ± 1.7 A | 18.4 ± 2.4 B | 8292. 8 ± 956 B | 1531 ± 186 B | 532. ± 39.5 | 92.92 ± 0.9 | 18.4 ± 0.6 | |
| Year × Rootstock | * | * | * | * | * | * | * | * | |
| Cohort of | Rootstock | Ratios (%) | Age of Trees | Ripe Cones Production | ||
|---|---|---|---|---|---|---|
| P1/P2 | P2/P3 | Years | Min to Max Number/Tree | Non-Productive Trees (%) | ||
| 2013 | PP | 84.6 | 83.6 | 4 | 0–3 | 14.3 |
| PH | 68.8 | 95.2 | 0–4 | 21.4 | ||
| 2014 | PP | 91.5 | 78.4 | 5 | 0–3 | 7.1 |
| PH | 81.6 | 72.5 | 0–4 | 7.1 | ||
| 2015 | PP | 53.2 | 47.3 | 6 | 0–7 | 14.3 |
| PH | 31.5 | 11.4 | 0–9 | 35.8 | ||
| 2016 | PP | 67.7 | 98.8 | 7 | 0–11 | 7.1 |
| PH | 57.2 | 100 | 0–7 | 28.6 | ||
| 2017 | PP | 90.0 | 97.7 | 8 | 0–6 | 35.7 |
| PH | 100 | 91.0 | 0–2 | 78.6 | ||
| 2018 | PP | 87.44 | 98.5 | 9 | 2–24 | 0 |
| PH | 91.2 | 98.0 | 0–24 | 21.4 | ||
| 2019 | PP | 61.7 | 10 | 12–55 | 0 | |
| PH | 77.5 | 4–29 | 0 | |||
| 2020 | PP | 11 | 14–58 | 0 | ||
| PH | 6–31 | 0 | ||||
| Percentage of: | PH | PP | Significance |
|---|---|---|---|
| Protein | 36.1 ± 0.37 | 36.3 ± 0.37 | ns |
| Total Sugars | 2.63 ± 0.11 | 2.43 ± 0.11 | ns |
| Crude Fibre | 4.30 ± 0.14 | 4.47 ± 0.14 | ns |
| Starch | 1.67 ± 0.05 | 1.73 ± 0.05 | ns |
| Fat | 44.33 ± 0.33 | 44.2 ± 0.33 | ns |
| Saturated fatty acids | 10.6 ± 0.11 | 10.4 ± 0.11 | ns |
| Palmitic (C16:0) | 6.0 ± 0.04 | 5.9 ± 0.04 | ns |
| Stearic (C18:0 | 3.4 ± 0.06 | 3.2 ± 0.06 | ns |
| Arachidic (C20:0) | 0.59 ± 0 | 0.54 ± 0 | * |
| Monounsaturated fatty acids | 41.3 ± 0.43 | 43.4 ± 0.43 | * |
| Palmitoleic (C16:1) | 0.23 ± 0.01 | 0.23 ± 0.01 | ns |
| Oleic (C18:2 n-9) | 38.3 ± 0.42 | 40.2 ± 0.42 | * |
| Gadoleic (C20:1 n = 11) | 0.70 ± 0.01 | 0.77 ± 0.01 | * |
| Polyunsaturated fatty acids | 47.9 ± 0.4 | 46.1 ± 0.4 | * |
| Linoleic (C18:2 n = 6) | 47.2 ± 0.4 | 45.3 ± 0.4 | * |
| Linolenic (C18:3) | 0.97 ± 0.02 | 0.97 ± 0.02 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guàrdia, M.; Teixidó, A.; Sanchez-Bragado, R.; Aletà, N. An Agronomic Approach to Pine Nut Production by Grafting Stone Pine on Two Rootstocks. Agriculture 2021, 11, 1034. https://doi.org/10.3390/agriculture11111034
Guàrdia M, Teixidó A, Sanchez-Bragado R, Aletà N. An Agronomic Approach to Pine Nut Production by Grafting Stone Pine on Two Rootstocks. Agriculture. 2021; 11(11):1034. https://doi.org/10.3390/agriculture11111034
Chicago/Turabian StyleGuàrdia, Mercè, Anna Teixidó, Rut Sanchez-Bragado, and Neus Aletà. 2021. "An Agronomic Approach to Pine Nut Production by Grafting Stone Pine on Two Rootstocks" Agriculture 11, no. 11: 1034. https://doi.org/10.3390/agriculture11111034