
Interactions between Biochar and Compost Treatment and Mycorrhizal Fungi to Improve the Qualitative Properties of a Calcareous Soil under Rhizobox Conditions



Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling and Preparation
2.2. Preparation of Biochar and Compost
2.3. Greenhouse Experiment
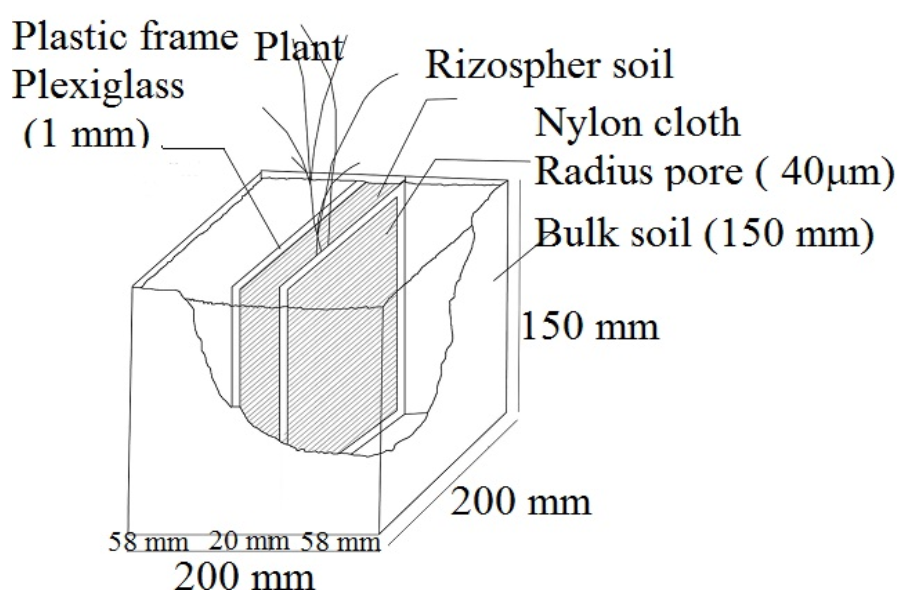
2.3.1. Rhizobox Experiment
2.3.2. Soil Analysis
2.3.3. Statistical Analysis
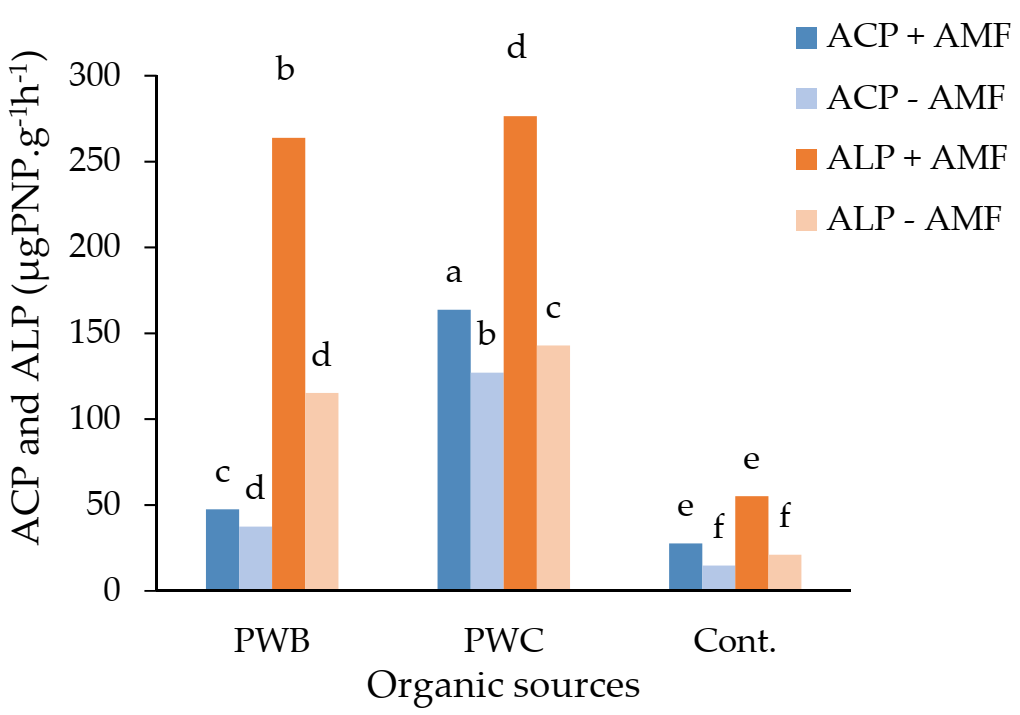
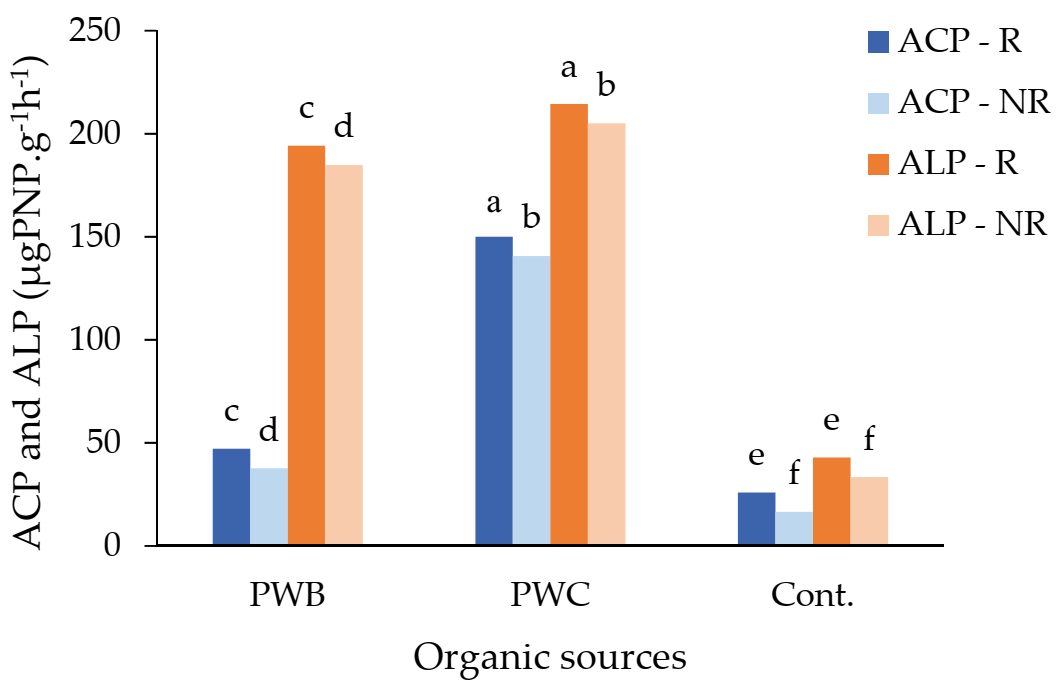
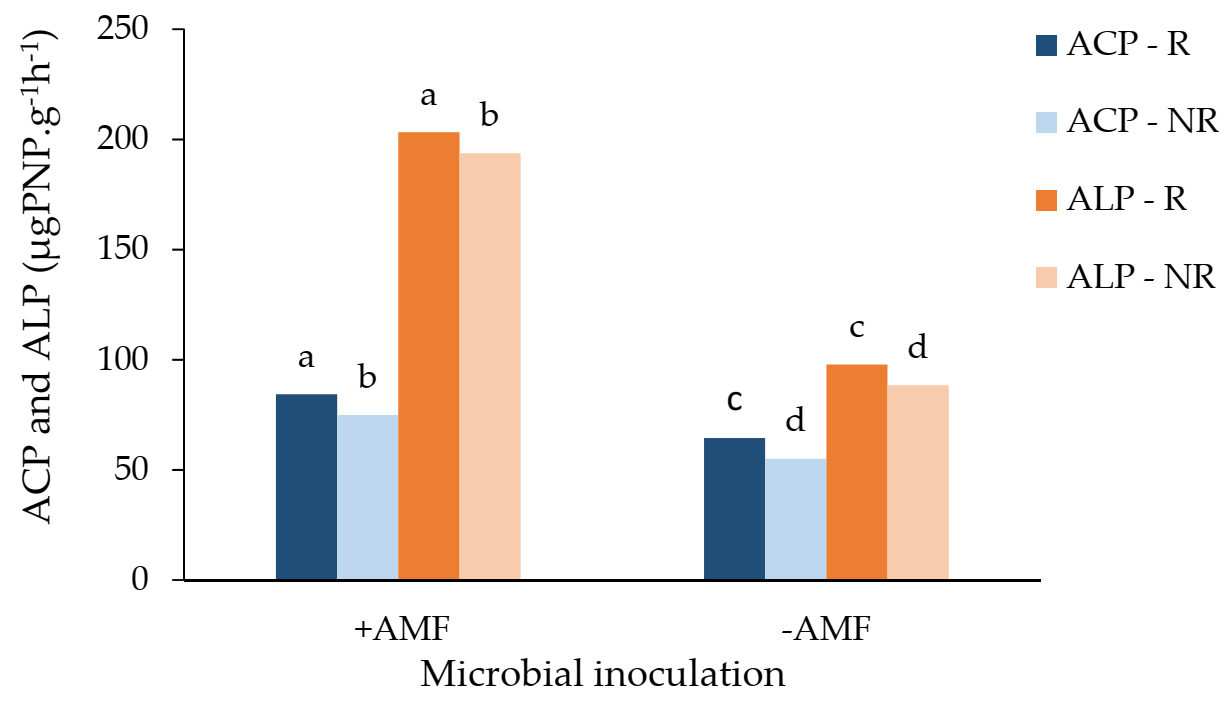
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moshiri, F.; Samavat, S.; Balali, M.R. Soil Organic Carbon: A Key Factor of Sustainable Agriculture in Iran; Global Symposium on Soil Organic Carbon: Rome, Italy, 2017; pp. 21–23. [Google Scholar]
- Rawat, J.; Saxena, J.; Sanwal, P. Biochar: A sustainable approach for improving plant growth and soil properties. In Biochar—An Imperative Amendment for Soil and the Environment; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Allelopathic interactions and allelochemicals: New possibilities or sustainable weed management. J. CRC Crit. Rev. Plant Sci. 2003, 22, 239–311. [Google Scholar] [CrossRef]
- Tomczyk, A.; Sokołowska, Z.; Boguta, P. Biochar physicochemical properties: Pyrolysis temperature and feedstock kind effects. J. Rev. Environ. Sci. Biotechnol. 2020, 19, 191–215. [Google Scholar] [CrossRef] [Green Version]
- Roberts, G.K.; Gloy, B.A.; Joseph, S.; Scott, N.R.; Lehmann, J. Life cycle assessment of biochar system: Estimating the enegetic, economic, and climate change potential. J. Environ. Sci. Technol. 2010, 44, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Ayilara, M.S.; Olanrewaju, O.S.; Babalola, O.O.; Odeyemi, O. Waste management through composting: Challenges and potentials. Sustainability 2020, 12, 4456. [Google Scholar] [CrossRef]
- Das, A.; Patel, D.P.; Munda, G.C.; Ghosh, P.K. Effect of organic and inorganic sources of nutrients on yield, nutrient uptake and soil fertility of maize (Zea mays)-mustard (Brassica campestris) cropping system. Indian J. Agric. Sci. 2010, 80, 85–88. [Google Scholar]
- Bünemann, E.K.; Bongiorno, G.; Bai, Z.; Creamer, R.E.; De Deyn, G.; de Goede, R.; Fleskens, L.; Geissen, V.; Kuyper, T.W.; Mäder, P.; et al. Soil quality–A critical review. J. Soil Biol. Biochem. 2018, 120, 105–125. [Google Scholar] [CrossRef]
- Luo, Y.; Zhou, X. Soil Respiration and the Environment, 1st ed.; Academic Press, An Imprint of Elsevier Science: Amsterdam, The Netherlands; London, UK, 2006; p. 328. [Google Scholar] [CrossRef]
- Samuel, A.D.; Brejea, R.; Domuta, C.; Bungau, S.; Cenusa, N.; Tit, D.M. Enzymatic indicators of soil quality. J. Environ. Prot. Ecol. 2017, 18, 871–878. [Google Scholar]
- Samuel, A.D.; Tit, D.M.; Melinte (Frunzulica), C.E.; Iovan, C.; Purza, L.; Gitea, M.; Bungau, S. Enzymological and Physicochemical Evaluation of the Effects of Soil Management Practices. Rev. Chim. 2017, 68, 2243–2247. [Google Scholar] [CrossRef]
- Gil-Sotres, F.; Trasar-Cepeda, C.; Leiros, M.C.; Seoane, S. Different approaches to evaluating soil quality using biochemical properties. J. Soil Biol. Biochem. 2005, 37, 877–887. [Google Scholar] [CrossRef]
- Liu, X.; Zheng, J.; Zhang, D.; Cheng, K.; Zhou, H.; Zhang, A.; Li, L.; Joseph, S.; Smith, P.; Crowley, D.; et al. Biochar has no effect on soil respiration across Chinese agricultural soils. J. Sci. Total Environ. 2016, 554, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Wei, M.; Liu, X.; He, Y.; Xu, X.; Wu, Z.; Yu, K.; Zheng, X. Biochar inoculated with Pseudomonas putida improves grape (Vitis vinifera L.) fruit quality and alters bacterial diversity. Rhizosphere 2020, 16. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota- a review. J. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Tejada, M.; Gonzalez, J.L. Crushed cotton gin compost on soil biological properties and rice yield. Eur. J. Agron. 2006, 25, 22–29. [Google Scholar] [CrossRef]
- Marinari, S.; Masciandaro, G.; Ceccanti, B.; Grego, S. Influence of organic and mineral fertilisers on soil biological and physical properties. J. Bioresour. Technol. 2000, 72, 9–17. [Google Scholar] [CrossRef]
- Guo, X.; Wan, Y.; Shakeel, M.; Wang, D.; Xiao, L. Effect of mycorrhizal fungi inoculation on bacterial diversity, community structure and fruit yield of blueberry. Rhizosphere 2021, 19. [Google Scholar] [CrossRef]
- Mickan, B.S.; Abbott, L.K.; Stefanova, K.; Solaiman, Z.M. Interactions between biochar and mycorrhizal fungi in a water-stressed agricultural soil. J. Mycorrhiza 2016, 26, 565–574. [Google Scholar] [CrossRef]
- Wathira, N.L.; Peter, W.; Sheila, O. Enhancement of colonisation of soybean roots by arbuscular mycorrhizal fungi using vermicompost and biochar. J. Int. Agric. For. Fish. 2016, 5, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Q.; Hu, J.; Wei, H.; Zhang, H.; Zhu, J. Relationship between plant roots, rhizosphere microorganisms, and nitrogen and its special focus on rice. J. Agric. 2021, 11, 234. [Google Scholar] [CrossRef]
- Vetterlein, D.; Carminati, A.; Kögel-Knabner, I.; Bienert, G.P.; Smalla, K.; Oburger, E.; Schnepf, A.; Banitz, T.; Tarkka, M.T.; Schlüter, S. Rhizosphere spatiotemporal organization–a key to rhizosphere functions. J. Front. Agron. 2020, 2, 8. [Google Scholar] [CrossRef]
- Liang, Y.; Nikolic, M.; Peng, Y.; Chen, W.; Jiang, Y. Organic manure stimulates biological activity and barley growth in soil subject to secondary salinization. J. Soil Biol. Biochem. 2005, 37, 1185–1195. [Google Scholar] [CrossRef]
- Hylander, L.D. Improvements of rhizoboxes used for studies of soil-root interactions. J. Commun. Soil Sci. Plant Anal. 2002, 33, 155–161. [Google Scholar] [CrossRef]
- Wenzel, W.W.; Wieshammer, G.; Fitz, W.J.; Puschenreiter, M. Novel rhizobox design to asses rhizosphere characteristics at high spatial resolution. J. Plant Soil. 2001, 237, 37–45. [Google Scholar] [CrossRef]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H.; Soltanpour, P.N.; Tabatabai, M.A.; Johnston, C.T.; Sumner, M.E. Methods of Soil Analysis: Part 3 Chemical Methods 5.3, 1st ed.; Soil Science Society of America Book Series 5; Soil Science Society of America: Madison, WI, USA, 1996; p. 1390. [Google Scholar] [CrossRef] [Green Version]
- Rajkovich, S.; Enders, A.; Hanley, K.; Hyland, C.; Zimmerman, A.R.; Lehmann, J. Corn growth and nitrogen nutrition after additions of biochars with varying properties to a temperate soil. J. Biol. Fertil. 2011, 48, 271–284. [Google Scholar] [CrossRef]
- Alfano, G.; Lustrato, G.; Lima, G.; Vitullo, D.; Ranalli, G. Characterization of composted olive mill wastes to predict potential plant disease suppressiveness. Biol. Control. 2011, 58, 199–207. [Google Scholar] [CrossRef]
- Hemashenpagam, N.; Selvaraj, T. Effect of arbuscular mycorrhizal (AM) fungus and plant growth promoting rhizomicroorganisms (PGPR’s) on medicinal plant Solanum viarum seedlings. J. Environ. Biol. 2011, 32, 579–583. [Google Scholar]
- Rorison, I.H. Ecological inferences from laboratory experiments on mineral nutrition. In Ecological Aspects of the Mineral Nutrition of Plants; Rorison, I.H., Ed.; Blackwell Sci. Publ.: Oxford, UK; Edinburgh, UK, 1969; pp. 155–176. [Google Scholar]
- Jenkinson, D.S.; Ladd, J.N. Microbial biomass in soil: Measurement and turnover. In Soil Biochemistry; Paul, E.A., Ladd, J.N., Eds.; New York Marcel Dekker, Inc.: New York, NY, USA, 1981; Volume 5, pp. 415–471. ISBN 0824711319. [Google Scholar]
- Brookes, P.C.; Powlson, D.S.; Jenkinson, D.S. Measurement of microbial biomass phosphorus in soil. J. Soil Biol. Biochem. 1982, 14, 319–329. [Google Scholar] [CrossRef]
- Anderson, J.P.E. Soil Respiration. In Methods of Soil Analysis, 2nd ed.; Part 2; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; ASA and SSSA: Madison, WI, USA, 1982; Volume 2, pp. 831–871. [Google Scholar]
- Alef, K.; Nannipieri, P. Methods in Applied Soil Microbiology and Biochemistry, 1st ed.; Academic Press: London, UK, 1995; p. 608. [Google Scholar]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. J. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Verheijen, F.; Jeffery, S.; Bastos, A.C.; van der Velde, M.; Diafas, I. Biochar Application to Soils—A Critical Scientific Review of Effects on Soil Properties, Processes and Functions; JRC Scientific and Technical Reports Luxembourg, European Commission: Luxembourg, 2010; p. 149. Available online: http://publications.jrc.ec.europa.eu/repository/bitstream/111111111/13558/1/jrc_biochar_soils.pdf (accessed on 5 November 2012).
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. J. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Babalola, O.A.; Adesodun, J.K.; Olasantan, F.O.; Adekunle, A.F. Responses of some soil biological, chemical and physical properties to short-term compost amendment. Int. J. Soil Sci. 2012, 7, 28–38. [Google Scholar] [CrossRef] [Green Version]
- Hammer, E.C.; Balogh-Brunstad, Z.; Jakobsen, I.; Olsson, P.A.; Stipp, S.L.S.; Rillig, M.C. A mycorrhizal fungus grows on biochar and captures phosphorus from its surfaces. J. Soil Biol. Biochem. 2014, 77, 252–260. [Google Scholar] [CrossRef]
- Jin, H.Y. Characterization of Microbial Life Colonizing Biochar and Biochar-Amended Soils. Ph.D. Thesis, Cornell University, Ithaca, NY, USA, 2010. [Google Scholar]
- Steiner, C.; Melear, N.; Harris, K.; Das, K.C. Biochar as bulking agent for poultry litter composting. J. Carbon Manag. 2011, 2, 227–230. [Google Scholar] [CrossRef]
- Steiner, C.; Teixeira, W.G.; Lehmann, J.; Zech, W. Microbial response to charcoal amendments of highly weathered soils and Amazonian Dark Earths in Central Amazonia-preliminary results. In Amazonian Dark Earths; Glaser, B., Woods, W.I., Eds.; Explorations in Space and Time Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2004; pp. 195–212. [Google Scholar]
- Smith, J.L.; Collins, H.P.; Bailey, V.L. The effect of young biochar on soil respiration. J. Soil Biol. Biochem. 2010, 42, 2345–2347. [Google Scholar] [CrossRef]
- Zaman, M.; Matsushima, M.; Chang, S.; Inubushi, K.; Nguyen, L.; Goto, S.; Kanek, O.F.; Yoneyama, T. Nitrogen mineralization, N2O production and soil microbiological prosperities as affected by long-term application of sewage sludge composts. J. Biol. Fertil. 2004, 40, 101–109. [Google Scholar] [CrossRef]
- De Neergaard, A.; Magid, J. Influence of the rhizosphere on microbial biomass and recently formed organic matter. Eur. J. Soil Sci. 2001, 52, 377–384. [Google Scholar] [CrossRef]
- Watzinger, A.; Feichtmair, S.; Kitzler, B.; Zehetner, F.; Kloss, S.; Wimmer, B.; Boltenstern, S.Z.; Soja, G. Soil microbial communities responded to biochar application in temperate soils and slowly metabolized 13C-labelled biochar as revealed by 13C PLFA analysis: Results from a short term incubation and pot experiment. Eur. J. Soil Sci. 2014, 65, 40–51. [Google Scholar] [CrossRef]
- Redel, Y.; Escudey, M.; Alvear, M.; Conrad, J.; Borie, F. Effects of tillage and crop rotation on chemical phosphorus forms and some related biological activities in a Chilean Ultisol. Soil Use Manag. 2011, 27, 221–228. [Google Scholar] [CrossRef]
- Brewer, C.E.; Schmidt-Rohr, K.; Satrio, J.A.; Brown, R.C. Characterization of biochar from fast pyrolysis and gasification systems. J. Environ. Prog. Sustain. 2009, 28, 386–396. [Google Scholar] [CrossRef]
- Urbankova, O.; Elbl, J.; Zahora, J. The effects of biochar on soil respiration in rhizosphere and non-rhizosphere soil. J. Mendel Net. 2014, 326–329. Available online: https://mnet.mendelu.cz/mendelnet2014/articles/52_urbankova_1077.pdf (accessed on 18 August 2021).
- Huang, Q.Y.; Chen, W.L.; Guo, X.J. Sequential fractionation of Cu, Zn and Cd in soils in the absence and presence of rhizobia. In Proceedings of the 17th WCSS, Bangkok, Thailand, 14–21 August 2002; p. 1453. [Google Scholar]
- Li, H.; Shao, H.; Li, W.; Bi, R.; Bai, Z. Improving soil enzyme activities and related quality properties of reclaimed soil by applying weathered coal in opencast-mining areas of the Chinese Loess Plateau. J. Clean 2012, 40, 233–238. [Google Scholar] [CrossRef]
- Zhao, Q.; Zeng, D.; Fan, Z. Nitrogen and phosphorus transformations in the rhizosphere of three tree species in a nutrient-poor sandy soil. J. Appl. Soil Ecol. 2010, 46, 341–346. [Google Scholar] [CrossRef]
- Benizri, E.; Nguyen, C.; Piutti, S.; Slezack-Deschaumes, S.; Philippot, L. Additions of maize root mucilage to soil changed the structure of the bacterial community. J. Soil Biol. Biochem. 2007, 39, 1230–1233. [Google Scholar] [CrossRef]
- Duineveld, B.M.; Kowalchuk, G.A.; Keijzer, A.; van Elsas, J.D.; van Veen, J.A. Analysis of bacterial communities in the rhizosphere of chrysanthemum via denaturating gradient gel electrophoresis of PCR-amplified 16S rRNA as well as DNA fragments coding for 16S rRNA. Appl. Environ. Microbiol. 2001, 67, 172–178. [Google Scholar] [CrossRef] [Green Version]
- Kourtev, P.S.; Ehrenfeld, J.G.; Häggblom, M. Experimental analysis of the effect of exotic and native plant species on the structure and function of soil microbial communities. Soil Biol. Biochem. 2003, 35, 895–905. [Google Scholar] [CrossRef]
- Chen, J.; van Groenigen, K.J.; Hungate, B.A.; Terrer, C.; van Groenigen, J.W.; Maestre, F.T.; Ying, S.C.; Luo, Y.Q.; Jørgensen, U.; Sinsabaugh, R.L.; et al. Long-term nitrogen loading alleviates phosphorus limitation in terrestrial ecosystems. Glob. Change Biol. 2020, 26, 5077–5086. [Google Scholar] [CrossRef]
- Qaswar, M.; Jing, H.; Ahmed, W.; Li, D.C.; Liu, S.J.; Lu, Z.; Cai, A.D.; Liu, L.S.; Xu, Y.M.; Gao, J.S.; et al. Yield sustainability, soil organic carbon sequestration and nutrients balance under long-term combined application of manure and inorganic fertilizers in acidic paddy soil. Soil Tillage Res. 2020, 198, 104569. [Google Scholar] [CrossRef]
- Samuel, A.D.; Bungau, S.; Tit, D.M.; Melinte, C.E.; Purza, L.; Badea, G.E. Effects of long term application of organic and mineral fertilizers on soil enzymes. Rev. Chim. 2018, 69, 2608–2612. [Google Scholar] [CrossRef]
- Igalavithana, A.D.; Lee, S.S.; Niazi, N.K.; Lee, Y.H.; Kim, K.H.; Park, J.H.; Moon, D.H.; Ok, Y.S. Assessment of soil health in urban sgriculture: Soil enzymes and microbial properties. Sustainability 2017, 9, 310. [Google Scholar] [CrossRef] [Green Version]
- Bungau, S.; Behl, T.; Aleya, L.; Bourgeade, P.; Aloui-Sossé, B.; Purza, A.L.; Abid, A.; Samuel, A.D. Expatiating the impact of anthropogenic aspects and climatic factors on long term soil monitoring and management. Environ. Sci. Pollut. Res. 2021, 30528–30550. [Google Scholar] [CrossRef]
- Chen, J.; Wei, X. Controlled-release fertilizers as a means to reduce nitrogen leaching and runoff in container-grown plant production. In Nitrogen in Agriculture—Updates; IntechOpen: London, UK, 2018; pp. 33–52. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Liang, X.; He, M.; Liu, Y.; Tian, G.; Shi, J. Manure biochar influence upon soil properties, phosphorus distribution and phosphatase activities: A microcosm incubation study. Chemosphere 2016, 142, 128–135. [Google Scholar] [CrossRef]
- Caldwell, B.A. Enzyme activities as a component of soil biodiversity: A review. Pedobiologia 2005, 49, 637–644. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Paz-Ferreiro, J.; Fu, S.; Mendez, A.; Gasco, G. Interactive effects of biochar and the earthworm (Pontoscolex corethrurus) on plant productivity and soil enzyme activities. J. Soils Sediments 2014, 14, 483–494. [Google Scholar] [CrossRef]
- Balík, J.; Pavlíková, D.; Vanĕk, V. The influence of long-term sewage sludge application on the activity of phosphatases in the rhizosphere of plants. Plant Soil Environ. 2007, 53, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Eivazi, F.; Tabatabai, M.A. Phosphatase in soils. Soil Biol. Biochem. 1977, 9, 167–172. [Google Scholar] [CrossRef]
- Anwar, M.; Patra, D.D.; Chand, S.; Alpesh, K.; Naqvi, A.A.; Khanuja, S.P.S. Effect of organic manures and inorganic fertilizer on growth, herb and oil yield, nutrient accumulation, and oil quality of French basil. Commun. Soil Sci. Plant Anal. 2005, 36, 1737–1746. [Google Scholar] [CrossRef]
- Mäder, P.; Edenhofer, S.; Boller, T.; Wiemken, A.; Niggli, U. Arbuscular mycorrhizae in a long-term field trial comparing low-input (organic biological) and high-input (conventional) farming systems in a crop rotation. J. Biol. Fertil. 2000, 31, 150–156. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Unit | Soil |
|---|---|---|
| pH | - | 7.53 |
| EC | dS m−1 | 0.47 |
| O.C | % | 0.25 |
| CaCO3 | % | 14.25 |
| N | % | 0.08 |
| P | mg kg−1 | 7.64 |
| K | mg kg −1 | 98 |
| Characteristics | Unit | PWB | PWC |
|---|---|---|---|
| pH | - | 7.29 | 7.05 |
| EC | dS m−1 | 0.08 | 17.87 |
| N | % | 0.54 | 3.72 |
| C | % | 67.53 | 30.02 |
| P | % | 0.27 | 7.54 |
| Microbial Inoculation | Organic Sources | OC (%) | MBC (mg kg−1) | MBP (mg kg−1) | BR (mg CO2 day−1 kg−1) | SIR (mg CO2 day−1 kg−1) |
|---|---|---|---|---|---|---|
| +AMF | PWB | 1.64 b | 789.3 b | 20.20 b | 53.15 b | 100.5 b |
| PWC | 2.0 a | 957.1 a | 85.72 a | 56.17 a | 112.3 a | |
| Cont. | 0.44 d | 429.0 d | 7.43 d | 25.97 e | 51.99 e | |
| −AMF | PWB | 0.87 c | 574.0 c | 15.01 c | 34.86 d | 78.21 d |
| PWC | 0.97 c | 573.7 c | 16.33 c | 43.63 c | 87.48 c | |
| Cont. | 0.32 d | 264.5 e | 3.52 e | 8.61 f | 15.46 f | |
| LSD0.05 | 0.18 | 7.31 | 2.41 | 2.88 | 4.34 |
| Soil | Organic Sources | OC (%) | MBC (mg kg−1) | MBP (mg kg−1) | BR (mg CO2 day−1 kg−1) | SIR (mg CO2 day−1 kg−1) |
|---|---|---|---|---|---|---|
| Rhizosphere | PWB | 1.29 bc | 687 c | 18.98 c | 46.99 b | 92.61 b |
| PWC | 1.52 a | 770.3 a | 52.41 a | 52.63 a | 103.3 a | |
| Cont. | 0.42 d | 352.7 e | 6.71 e | 20.27 d | 37.13 d | |
| Non-rhizosphere | PWB | 1.22 c | 676.3 d | 16.23 d | 41.03 c | 86.06 c |
| PWC | 1.45 ab | 760.4 b | 49.65 b | 47.16 b | 96.51 b | |
| Cont. | 0.35 d | 340.8 f | 4.24 f | 14.31 e | 30.33 e | |
| LSD0.05 | 0.18 | 7.31 | 2.41 | 2.88 | 4.34 |
| Microbial Inoculation | Soil | OC (%) | MBC (mg kg−1) | MBP (mg kg−1) | BR (mg CO2 day−1 kg−1) | SIR (mg CO2 day−1 kg−1) |
|---|---|---|---|---|---|---|
| +AMF | Rhizosphere | 1.39 a | 730.9 a | 39.08 a | 47.86 a | 91.64 a |
| Non-rhizosphere | 1.33 a | 719.4 b | 36.49 b | 42.33 b | 84.89 b | |
| −AMF | Rhizosphere | 0.75 b | 475.7 c | 12.99 c | 32.06 c | 63.73 c |
| Non-rhizosphere | 0.69 b | 465.6 d | 10.26 d | 26.01 d | 57.04 d | |
| LSD0.05 | 0.18 | 7.31 | 2.41 | 2.88 | 4.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vahedi, R.; Rasouli-Sadaghiani, M.; Barin, M.; Vetukuri, R.R. Interactions between Biochar and Compost Treatment and Mycorrhizal Fungi to Improve the Qualitative Properties of a Calcareous Soil under Rhizobox Conditions. Agriculture 2021, 11, 993. https://doi.org/10.3390/agriculture11100993
Vahedi R, Rasouli-Sadaghiani M, Barin M, Vetukuri RR. Interactions between Biochar and Compost Treatment and Mycorrhizal Fungi to Improve the Qualitative Properties of a Calcareous Soil under Rhizobox Conditions. Agriculture. 2021; 11(10):993. https://doi.org/10.3390/agriculture11100993
Chicago/Turabian StyleVahedi, Roghayeh, MirHassan Rasouli-Sadaghiani, Mohsen Barin, and Ramesh Raju Vetukuri. 2021. "Interactions between Biochar and Compost Treatment and Mycorrhizal Fungi to Improve the Qualitative Properties of a Calcareous Soil under Rhizobox Conditions" Agriculture 11, no. 10: 993. https://doi.org/10.3390/agriculture11100993