
Evolutionary Subdivision of Domestic Chickens: Implications for Local Breeds as Assessed by Phenotype and Genotype in Comparison to Commercial and Fancy Breeds



 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Phenotypic Characteristics of the Studied Chicken Populations
2.3. Genotyping of Chickens and SNP Data Processing
2.4. Mathematical and Statistical Analyzes
3. Results
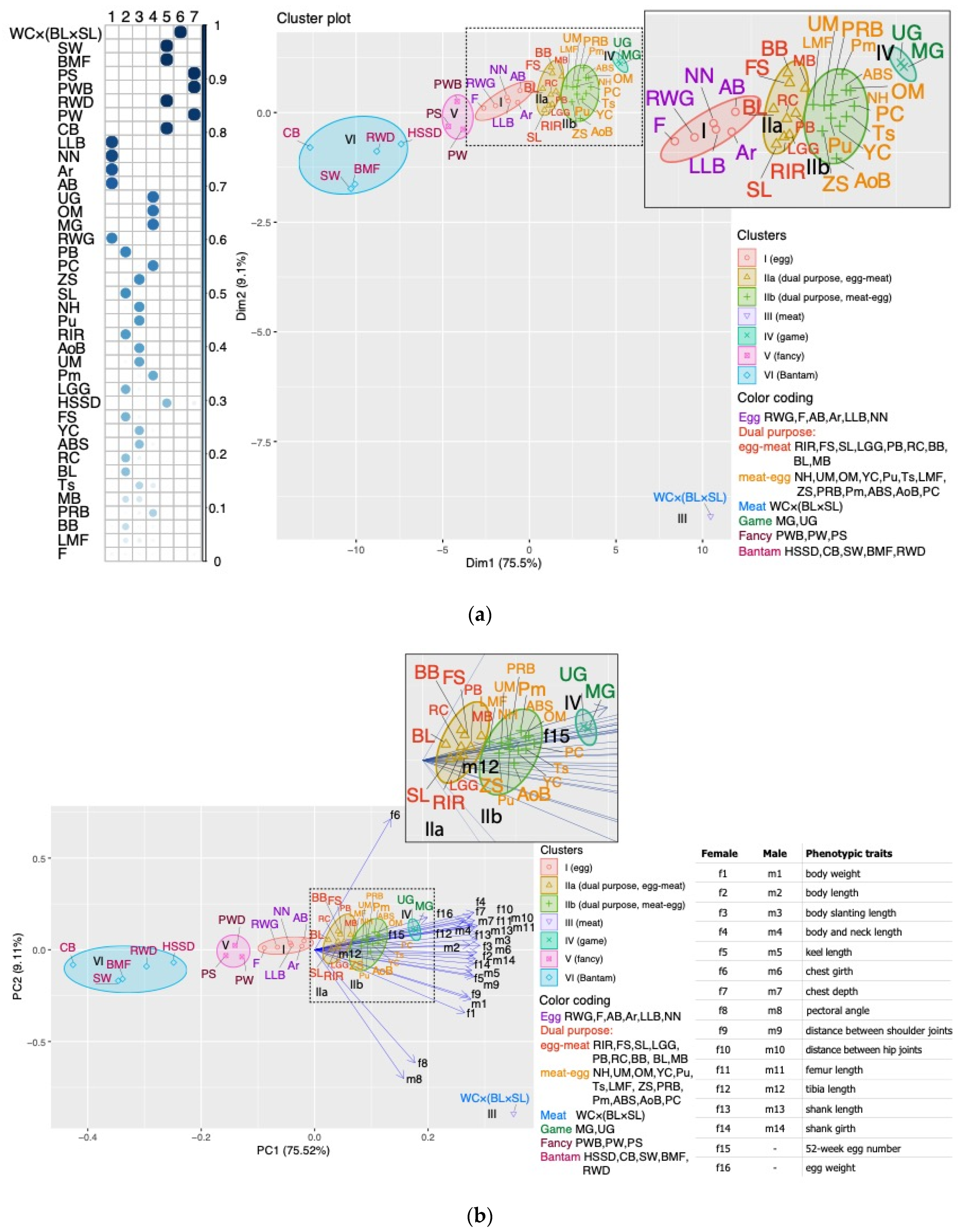
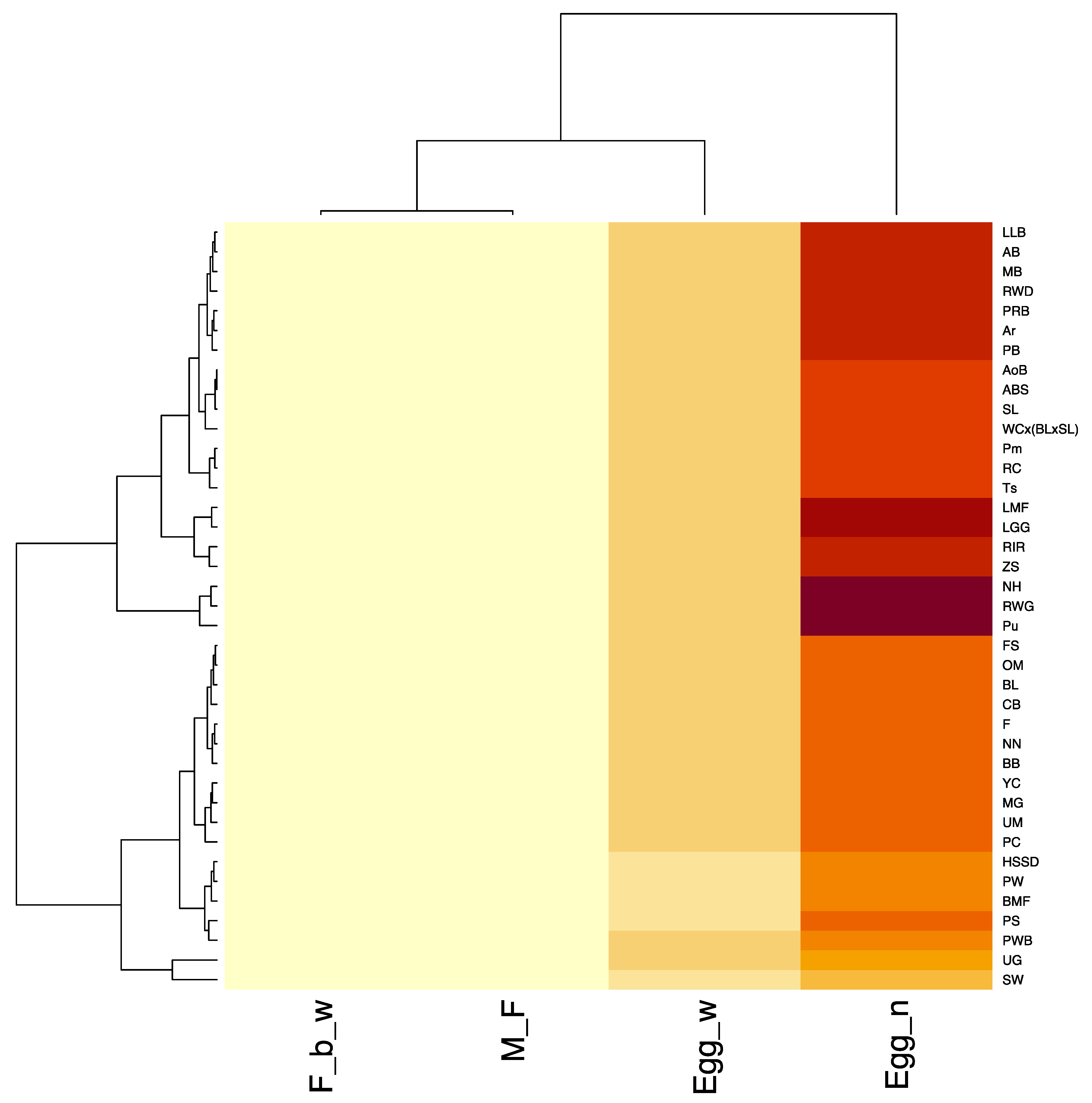
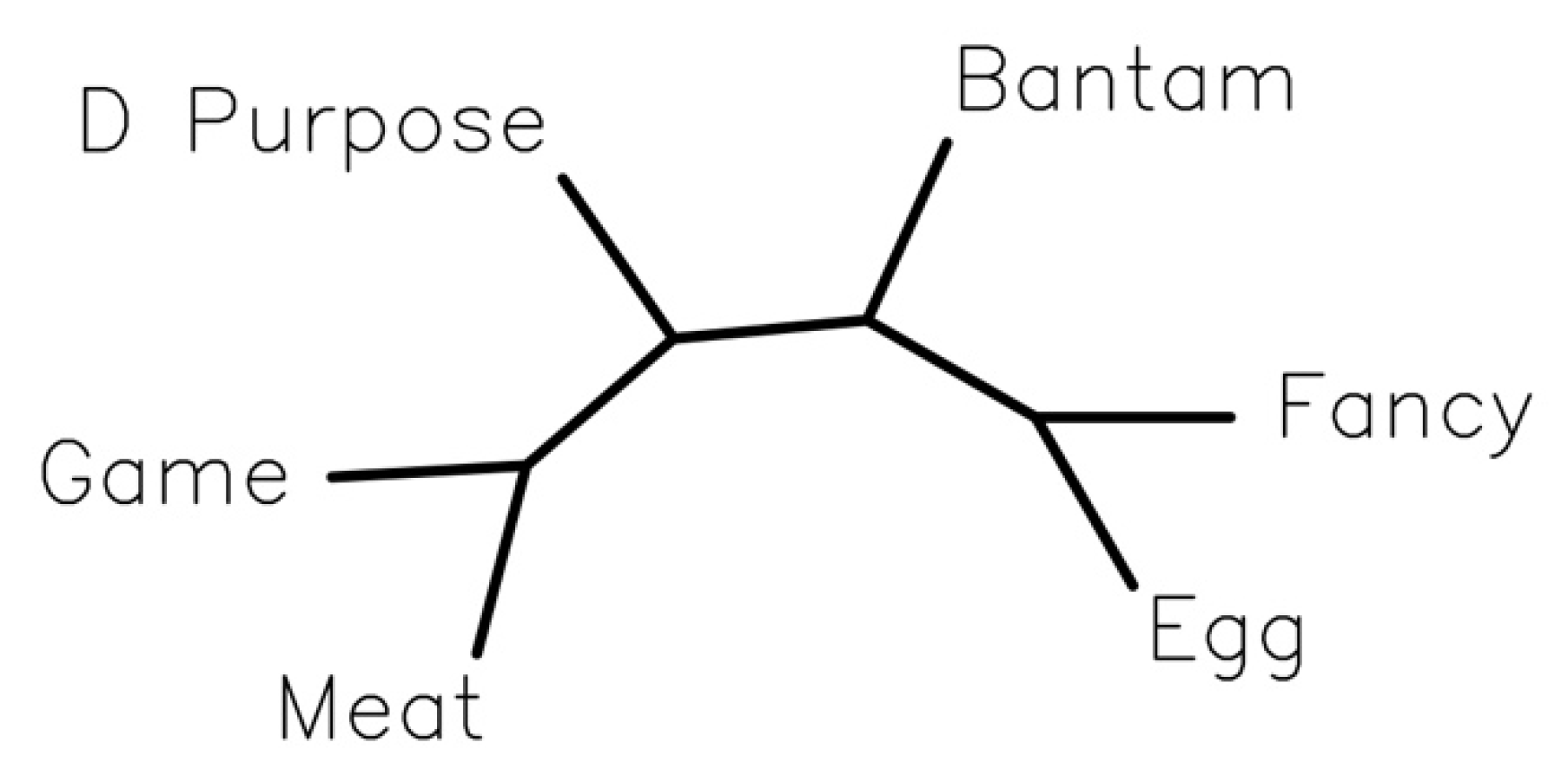
3.1. Analysis of Phenotypic Traits and Breed Clustering by Phenotype
3.2. Analysis of Genetic Variation at the Locus NCAPG-LCORL
3.2.1. Population Genetic Parameters
3.2.2. Clustering of the Analyzed Chicken Breeds by Genotypes
3.2.3. Assessment of Breed Admixture by Genotypes
3.2.4. Genetic Differentiation of Populations by Alleles
3.2.5. Overlapping of SNPs with QTLs
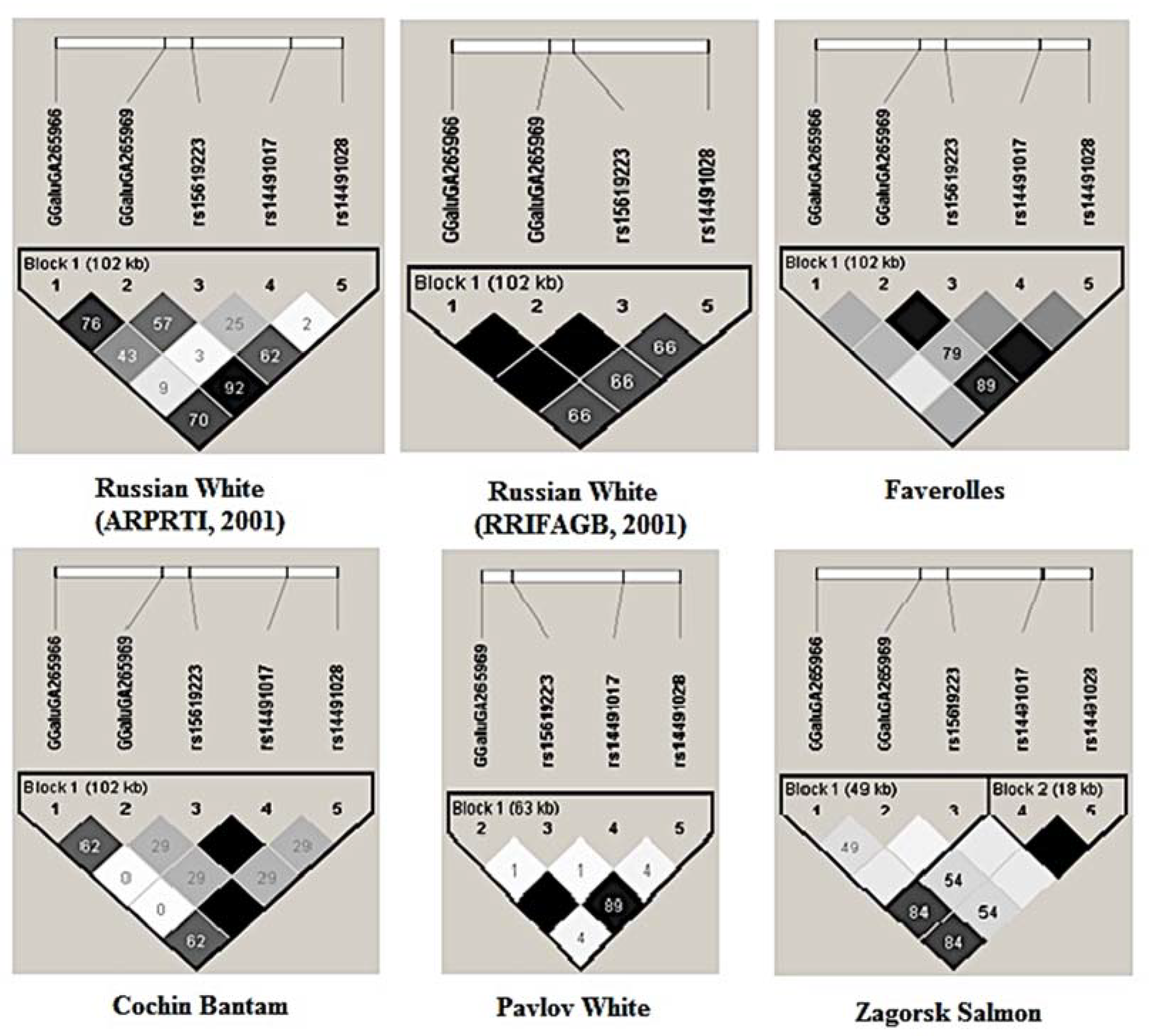
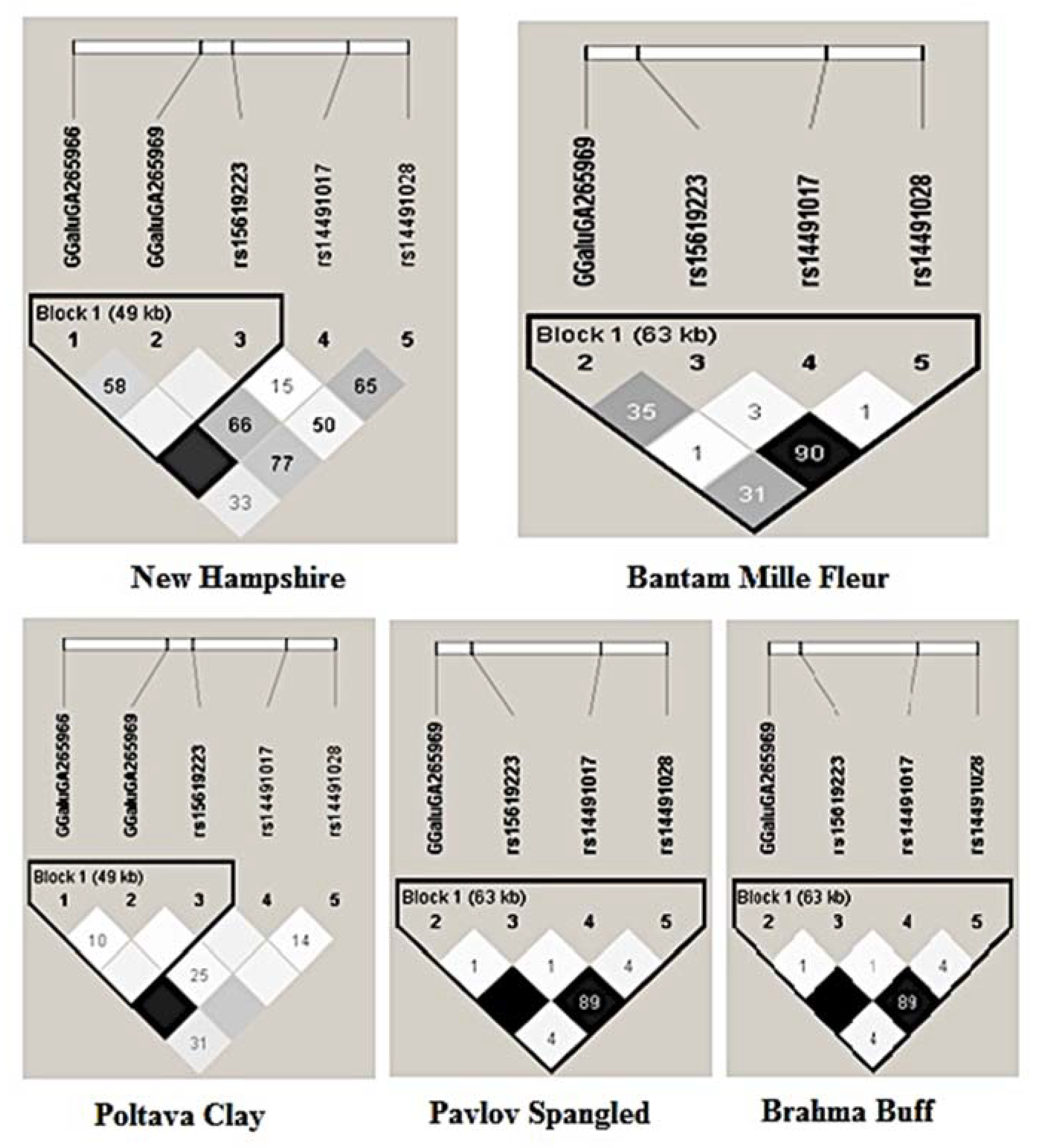
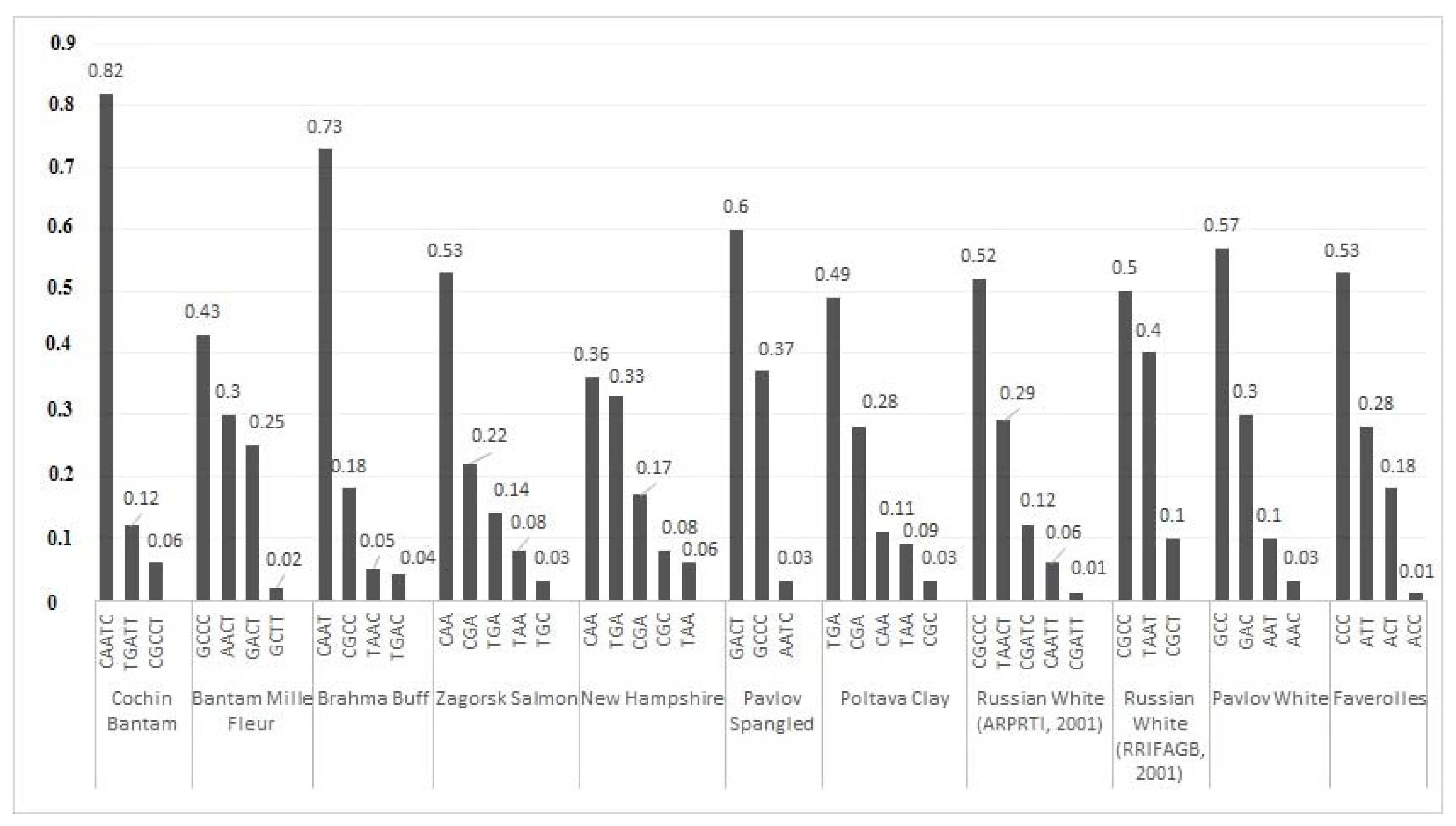
3.2.6. LD Structure of Gene Pool Breeds Based on Haplotypes
4. Discussion
4.1. Analysis of Phenotypic Traits and a Model for Clustering Breeds by Phenotypes (PCM)
4.2. Genotypic Models of Clustering and Admixture
4.3. Analysis of Genetic Variation at the Locus NCAPG-LCORL
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Romanov, M.N. Genetics of Broodiness in Poultry—A Review. Asian-Australas. J. Anim. Sci. 2001, 14, 1647–1654. [Google Scholar] [CrossRef]
- Dodgson, J.B.; Romanov, M.N. Use of Chicken Models for the Analysis of Human Disease. Curr. Protoc. Hum. Genet. 2004, 40, 15.5.1–15.5.15. [Google Scholar] [CrossRef] [PubMed]
- Moiseyeva, I.G.; Romanov, M.N.; Nikiforov, A.A.; Sevastyanova, A.A.; Semyenova, S.K. Evolutionary Relationships of Red Jungle Fowl and Chicken Breeds. Genet. Sel. Evol. 2003, 35, 403–423. [Google Scholar] [CrossRef] [Green Version]
- Kholofelo Malomane, D.; Simianer, H.; Reimer, C.; Weigend, A.; Weigend, S. Different Evolutionary Dynamics Revealed by Functional SNP Classes in Global Chicken Groups. Worlds Poult. Sci. J. 2018. Suppl.: The XVth European Poultry Conference. Conference Information and Proceedings, Dubrovnik, Croatia, 17–21 September 2018, p. 87, Abstract ID 207. [Google Scholar]
- Makarova, A.V.; Mitrofanova, O.V.; Vakhrameev, A.B.; Dementeva, N.V. Molecular-Genetic Bases of Plumage Coloring in Chicken. Vavilovskii Zh. Genet. Sel. (Vavilov J. Genet. Breed.) 2019, 23, 343–354. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.-S.; Thakur, M.; Peng, M.-S.; Jiang, Y.; Frantz, L.A.F.; Li, M.; Zhang, J.-J.; Wang, S.; Peters, J.; Otecko, N.O.; et al. 863 Genomes Reveal the Origin and Domestication of Chicken. Cell Res. 2020, 30, 693–701. [Google Scholar] [CrossRef]
- Bennett, C.E.; Thomas, R.; Williams, M.; Zalasiewicz, J.; Edgeworth, M.; Miller, H.; Coles, B.; Foster, A.; Burton, E.J.; Marume, U. The Broiler Chicken as a Signal of a Human Reconfigured Biosphere. R. Soc. Open Sci. 2018, 5, 180325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.-S.; Huo, Y.-X.; Li, Y.; Otecko, N.O.; Su, L.-Y.; Xu, H.-B.; Wu, S.-F.; Peng, M.-S.; Liu, H.-Q.; Zeng, L.; et al. Comparative Population Genomics Reveals Genetic Basis Underlying Body Size of Domestic Chickens. J. Mol. Cell Biol. 2016, 8, 542–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.-J.; Zhang, L.; Ren, P.; Wang, Y.; Yin, L.-Q.; Ran, J.-S.; Zhang, X.-X.; Liu, Y.-P. Genotype Frequency Distributions of 28 SNP Markers in Two Commercial Lines and Five Chinese Native Chicken Populations. BMC Genet. 2020, 21, 12. [Google Scholar] [CrossRef] [PubMed]
- Wężyk, S.; Cywa-Benko, K.; Romanov, M.N. Ochrona Przed Zagładą Rodzimych Ras Drobiu w Krajach Wschodniej Europy/Protection against the Extinction of the Native Breeds of Poultry in the Countries of Eastern Europe. In Proceedings of the International Symposium Conservation Measures for Rare Farm Animal Breeds, Balice, Poland, 17–19 May 1994; National Research Institute of Animal Production: Balice, Poland; Polish Society of Animal Production: Balice, Poland, 1994; pp. 25–26. [Google Scholar]
- Romanov, M.N.; Sakhatsky, N.I. Ochrona Zasobów Genetycznych Drobiu na Ukrainie/Conservation of Poultry Genetic Resources in Ukraine. In Summaries of the 9th International Symposium of Young Poultry Scientists, Siedlce, Poland, 8–10 June 1995; World’s Poultry Science Association, Polish Branch: Siedlce, Poland, 1995; pp. 87–88. [Google Scholar]
- Romanov, M.N.; Sakhatsky, N.I. Inventory of Poultry Genetic Resources in Ukraine. In Naučno-Tehničeskij Bûlleten-Institut Pticevodstva; Poultry Research Institute, Ukrainian Academy of Agrarian Sciences: Kharkiv, Ukraine, 1995; No. 34; pp. 3–14. [Google Scholar]
- Wezyk, S.; Cywa-Benko, K.; Romanov, M.N. Protection against the Extinction of the Native Breeds of Poultry in Eastern European Countries. In Proceedings of the International Symposium “Conservation Measures for Rare Farm Animal Breeds, Balice, Poland, 17–19 May 1994; pp. 89–101. [Google Scholar]
- Romanov, M.N.; Weigend, S.; Bondarenko, Y.V.; Podstreshny, A.P.; Kutnyuk, P.I.; Sakhatsky, N.I. Studies on poultry germplasm diversity and conservation in Ukraine. In Proceedings of the Poultry Genetics Symposium, Mariensee, Germany, 6–8 October 1999; Working Group 3 of WPSA: Mariensee, Germany, 1999; p. 140. [Google Scholar]
- Dementeva, N.V.; Vakhrameev, A.B.; Larkina, T.A.; Mitrofanova, O.V. Efficiency of Using SNP Markers in the MSTN Gene in the Selection of the Pushkin Breed Chickens. Vavilovskii Zh. Genet. Sel. (Vavilov J. Genet. Breed.) 2019, 23, 993–998. [Google Scholar] [CrossRef]
- Dou, T.; Shen, M.; Ma, M.; Qu, L.; Li, Y.; Hu, Y.; Lu, J.; Guo, J.; Wang, X.; Wang, K. Genetic Architecture and Candidate Genes Detected for Chicken Internal Organ Weight with a 600 K Single Nucleotide Polymorphism Array. Asian-Australas. J. Anim. Sci. 2019, 32, 341–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, G.; Shen, M.; Yuan, J.; Sun, C.; Duan, Z.; Qu, L.; Dou, T.; Ma, M.; Lu, J.; Guo, J.; et al. Genome-Wide Association Study Dissects Genetic Architecture Underlying Longitudinal Egg Weights in Chickens. BMC Genom. 2015, 16, 746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, M.; Qu, L.; Ma, M.; Dou, T.; Lu, J.; Guo, J.; Hu, Y.; Wang, X.; Li, Y.; Wang, K.; et al. A Genome-Wide Study to Identify Genes Responsible for Oviduct Development in Chickens. PLoS ONE 2017, 12, e0189955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, A.R.; Esko, T.; Yang, J.; Vedantam, S.; Pers, T.H.; Gustafsson, S.; Chu, A.Y.; Estrada, K.; Luan, J.; Kutalik, Z.; et al. Defining the Role of Common Variation in the Genomic and Biological Architecture of Adult Human Height. Nat. Genet. 2014, 46, 1173–1186. [Google Scholar] [CrossRef] [Green Version]
- Okada, Y.; Kamatani, Y.; Takahashi, A.; Matsuda, K.; Hosono, N.; Ohmiya, H.; Daigo, Y.; Yamamoto, K.; Kubo, M.; Nakamura, Y.; et al. A Genome-Wide Association Study in 19 633 Japanese Subjects Identified LHX3-QSOX2 and IGF1 as Adult Height Loci. Hum. Mol. Genet. 2010, 19, 2303–2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carty, C.L.; Johnson, N.A.; Hutter, C.M.; Reiner, A.P.; Peters, U.; Tang, H.; Kooperberg, C. Genome-Wide Association Study of Body Height in African Americans: The Women’s Health Initiative SNP Health Association Resource (SHARe). Hum. Mol. Genet. 2012, 21, 711–720. [Google Scholar] [CrossRef] [Green Version]
- Soranzo, N.; Rivadeneira, F.; Chinappen-Horsley, U.; Malkina, I.; Richards, J.B.; Hammond, N.; Stolk, L.; Nica, A.; Inouye, M.; Hofman, A.; et al. Meta-Analysis of Genome-Wide Scans for Human Adult Stature Identifies Novel Loci and Associations with Measures of Skeletal Frame Size. PLoS Genet. 2009, 5, e1000445. [Google Scholar] [CrossRef]
- Lindholm-Perry, A.K.; Sexten, A.K.; Kuehn, L.A.; Smith, T.P.; King, D.A.; Shackelford, S.D.; Wheeler, T.L.; Ferrell, C.L.; Jenkins, T.G.; Snelling, W.M.; et al. Association, Effects and Validation of Polymorphisms within the NCAPG-LCORL Locus Located on BTA6 with Feed Intake, Gain, Meat and Carcass Traits in Beef Cattle. BMC Genet. 2011, 12, 103. [Google Scholar] [CrossRef] [Green Version]
- Rubin, C.-J.; Megens, H.-J.; Barrio, A.M.; Maqbool, K.; Sayyab, S.; Schwochow, D.; Wang, C.; Carlborg, Ö.; Jern, P.; Jørgensen, C.; et al. Strong Signatures of Selection in the Domestic Pig Genome. Proc. Natl. Acad. Sci. USA 2012, 109, 19529–19536. [Google Scholar] [CrossRef] [Green Version]
- Metzger, J.; Schrimpf, R.; Philipp, U.; Distl, O. Expression Levels of LCORL are Associated with Body Size in Horses. PLoS ONE 2013, 8, e56497. [Google Scholar] [CrossRef] [Green Version]
- Al-Mamun, H.A.; Kwan, P.S.; Clark, A.; Ferdosi, M.H.; Tellam, R.; Gondro, C. Genome-Wide Association Study of Body Weight in Australian Merino Sheep Reveals an Orthologous Region on OAR6 to Human and Bovine Genomic Regions Affecting Height and Weight. Genet. Sel. Evol. 2015, 47, 66. [Google Scholar] [CrossRef] [Green Version]
- Takasuga, A. PLAG1 and NCAPG-LCORL in Livestock. Anim. Sci. J. 2016, 87, 159–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyu, S.; Arends, D.; Nassar, M.K.; Brockmann, G.A. Fine Mapping of a Distal Chromosome 4 QTL Affecting Growth and Muscle Mass in a Chicken Advanced Intercross Line. Anim. Genet. 2017, 48, 295–302. [Google Scholar] [CrossRef]
- Dementieva, N.V.; Kudinov, A.A.; Larkina, T.A.; Mitrofanova, O.V.; Dysin, A.P.; Terletsky, V.P.; Tyshchenko, V.I.; Griffin, D.K.; Romanov, M.N. Genetic Variability in Local and Imported Germplasm Chicken Populations as Revealed by Analyzing Runs of Homozygosity. Animals 2020, 10, 1887. [Google Scholar] [CrossRef] [PubMed]
- Dementieva, N.V.; Mitrofanova, O.V.; Dysin, A.P.; Kudinov, A.A.; Stanishevskaya, O.I.; Larkina, T.A.; Plemyashov, K.V.; Griffin, D.K.; Romanov, M.N.; Smaragdov, M.G. Assessing the Effects of Rare Alleles and Linkage Disequilibrium on Estimates of Genetic Diversity in the Chicken Populations. Animal 2021, 15, 100171. [Google Scholar] [CrossRef] [PubMed]
- Paronyan, I.A.; Plemyashov, K.V.; Segal, E.L.; Yurchenko, O.P.; Shabanova, S.A.; Vakhrameev, A.B.; Karpukhina, I.V.; Makarova, A.V.; Pervushina, A.T. Breeds and Populations of Chickens Bred at the Germplasm Farm of the State Scientific Institution VNIIGRZh of the Russian Agricultural Academy: Album; GNU VNIIGRZh: St. Petersburg, Russia, 2014; Available online: http://vniigen.ru/wp-content/uploads/2017/04/Katalog-Kur-1.pdf (accessed on 9 July 2021).
- Bogolyubsky, S.I. Poultry Breeding; Agropromizdat: Moscow, Russia, 1991. [Google Scholar]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-Generation PLINK: Rising to the Challenge of Larger and Richer Datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- RDocumentation. Heatmap: Draw a Heat Map. Available online: https://www.rdocumentation.org/packages/stats/versions/3.6.2/topics/heatmap (accessed on 9 July 2021).
- Shitikov, V.K.; Mastitsky, S.E. 10.4.2 Fuzzy k-Means Method (Fuzzy Analysis Clustering). In Classification, Regression, Data Mining Algorithms Using R. Electronic Book. 2017. Available online: https://ranalytics.github.io/data-mining/104-Other-Clustering-Methods.html#sec_10_4_2 (accessed on 9 July 2021).
- Fanny: Fuzzy Analysis Clustering. Documentation for Package ‘Cluster’ Version 2.1.0. “Finding Groups in Data”: Cluster Analysis Extended Rousseeuw et al. 2019. Available online: https://rdrr.io/cran/cluster/man/fanny.html (accessed on 9 July 2021).
- Suzuki, R.; Shimodaira, H. Pvclust: An R Package for Assessing the Uncertainty in Hierarchical Clustering. Bioinformatics 2006, 22, 1540–1542. [Google Scholar] [CrossRef]
- Zhao, Q.; Hautamaki, V.; Fränti, P. Knee Point Detection in BIC for Detecting the Number of Clusters. In Lecture Notes in Computer Science; Blanc-Talon, J., Bourennane, S., Philips, W., Popescu, D., Scheunders, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; Volume 5259, pp. 664–673. [Google Scholar]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. Version 1.0.5; 22 August 2017. Available online: https://cran.r-project.org/web/packages/factoextra/index.html (accessed on 9 July 2021).
- Alexander, D.H.; Novembre, J.; Lange, K. Fast Model-Based Estimation of Ancestry in Unrelated Individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, A.L.; Patterson, N.J.; Plenge, R.M.; Weinblatt, M.E.; Shadick, N.A.; Reich, D. Principal Components Analysis Corrects for Stratification in Genome-Wide Association Studies. Nat. Genet. 2006, 38, 904–909. [Google Scholar] [CrossRef]
- Ayala, F.J.; Kiger, J.A., Jr. Modern Genetics, 2nd ed.; Benjamin/Cummings: Menlo Park, CA, USA; London, UK, 1984. [Google Scholar]
- Wright, S. The Genetical Structure of Populations. Ann. Eugen. 1951, 15, 323–354. [Google Scholar] [CrossRef] [PubMed]
- Pilot, M.; Greco, C.; vonHoldt, B.M.; Jędrzejewska, B.; Randi, E.; Jędrzejewski, W.; Sidorovich, V.E.; Ostrander, E.A.; Wayne, R.K. Genome-Wide Signatures of Population Bottlenecks and Diversifying Selection in European Wolves. Heredity 2014, 112, 428–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boc, A.; Diallo, A.B.; Makarenkov, V. T-REX: A Web Server for Inferring, Validating and Visualizing Phylogenetic Trees and Networks. Nucleic Acids Res. 2012, 40, 573–579. [Google Scholar] [CrossRef] [Green Version]
- The Chicken Quantitative Trait Locus (QTL) Database (Chicken QTLdb). Available online: https://www.animalgenome.org/cgi-bin/QTLdb/GG/index (accessed on 4 September 2021).
- Wellek, S.; Ziegler, A. A Genotype-Based Approach to Assessing the Association between Single Nucleotide Polymorphisms. Hum. Hered. 2009, 67, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and Visualization of LD and Haplotype Maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]
- Genome Reference Consortium Chicken Build 6a. Available online: https://www.ncbi.nlm.nih.gov/assembly/GCF_000002315.6 (accessed on 4 September 2021).
- Kudinov, A.A.; Dementieva, N.V.; Mitrofanova, O.V.; Stanishevskaya, O.I.; Fedorova, E.S.; Larkina, T.A.; Mishina, A.I.; Plemyashov, K.V.; Griffin, D.K.; Romanov, M.N. Genome-Wide Association Studies Targeting the Yield of Extraembryonic Fluid and Production Traits in Russian White Chickens. BMC Genom. 2019, 20, 270. [Google Scholar] [CrossRef] [PubMed]
- Dementeva, N.V.; Larkina, T.A.; Mitrofanova, O.V.; Fedorova, E.S.; Pozdnyakova, T.E. Association of Single Nucleotide Polymorphism in LCORL Gene with the Productive Traits in Russian White Chicken Breed. Ptitsevodstvo 2019, 5, 14–17. [Google Scholar] [CrossRef]
- FAO. Phenotypic Characterization of Animal Genetic Resources; FAO Animal Production and Health Guidelines No. 11; FAO: Rome, Italy, 2012. [Google Scholar]
- Dorji, N.; Sunar, S.K. Short Communication: Morphometric Variations among Five Bhutanese Indigenous Chickens (Gallus domesticus). J. Anim. Poult. Sci. 2014, 3, 76–85. [Google Scholar]
- Mel’nyk, Y.F.; Mykytyuk, D.M.; Bilous, O.V.; Kudryavs’ka, N.V.; Burkat, V.P.; Huzyev, I.V.; Podoba, B.Y.; Sharan, P.I.; Kovtun, S.I.; Platonova, N.P.; et al. Program of Preservation of the Gene Pool of Main Types of Farm Animals in Ukraine for the Period Till 2015; Aristey: Kyiv, Ukraine, 2009. [Google Scholar]
- Negassa, D.; Melesse, A.; Banerjee, S. Phenotypic Characterization of Indigenous Chicken Populations in Southeastern Oromia Regional State of Ethiopia. Anim. Genet. Resour. Inf. 2014, 55, 101–113. [Google Scholar] [CrossRef]
- Assefa, S.; Melesse, A.; Banerjee, S. Egg Production and Linear Body Measurement Traits of Local and Three Exotic Chicken Genotypes Reared under Two Agroecological Zones. Int. J. Ecol. Ecosolut. 2018, 5, 18–23. [Google Scholar]
- Romanov, M.N. Study of Feathering Phenotypes in Hisex Brown Parent Lines and Their Hybrids—A Population-Genetic Analysis. In Proceedings of the 9th European Poultry Conference, Glasgow, UK, 7–12 August 1994; World’s Poultry Science Association, UK Branch: Glasgow, UK; Volume 1, pp. 352–354. [Google Scholar]
- Weigend, S.; Romanov, M.N. Current Strategies for the Assessment and Evaluation of Genetic Diversity in Chicken Resources. In Abstracts and Proceedings of the XXI World’s Poultry Congress, Montreal, QC, Canada, 20–24 August 2000; CD-ROM; World’s Poultry Science Association: Montreal, QC, Canada, 2000. [Google Scholar]
- Otecko, N.O.; Ogali, I.; Ng’ang’a, S.I.; Mauki, D.H.; Ogada, S.; Moraa, G.K.; Lichoti, J.; Agwanda, B.; Peng, M.S.; Ommeh, S.C.; et al. Phenotypic and Morphometric Differentiation of Indigenous Chickens from Kenya and Other Tropical Countries Augments Perspectives for Genetic Resource Improvement and Conservation. Poult. Sci. 2019, 98, 2747–2755. [Google Scholar] [CrossRef]
- Tyasi, T.L.; Mashiloane, K.; Mokoena, K. Comparison of Some Linear Body Measurement Traits of Local and Commercial Chicken Breeds of South Africa. Sib. J. Life Sci. Agric. 2021, 13, 134–143. [Google Scholar] [CrossRef]
- Liu, R.; Sun, Y.; Zhao, G.; Wang, H.; Zheng, M.; Li, P.; Liu, L.; Wen, J. Identification of Loci and Genes for Growth Related Traits from a Genome-Wide Association Study in a Slow- × Fast-Growing Broiler Chicken Cross. Genes Genom. 2015, 37, 829–836. [Google Scholar] [CrossRef]
- Han, Y.J.; Chen, Y.; Liu, Y.; Liu, X.L. Sequence Variants of the LCORL Gene and Its Association with Growth and Carcass Traits in Qinchuan Cattle in China. J. Genet. 2017, 96, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Tiley, G.P.; Pandey, A.; Kimball, R.T.; Braun, E.L.; Burleigh, J.G. Whole Genome Phylogeny of Gallus: Introgression and Data-Type Effects. Avian Res. 2020, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Dementeva, N.V.; Romanov, M.N.; Kudinov, A.A.; Mitrofanova, O.V.; Stanishevskaya, O.I.; Terletsky, V.P.; Fedorova, E.S.; Nikitkina, E.V.; Plemyashov, K.V. The Study of the Structure of the Gene Pool of the Russian White Breed of Chickens by the Method of Genomic SNP-Scanning. Selskokhoziaĭstvennaia Biol. (Agric. Biol.) 2017, 52, 1166–1174. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, O.; Odawara, S.; Takahashi, H.; Nirasawa, K.; Oyamada, Y.; Yamamoto, R.; Ishii, K.; Nagamine, Y.; Takeda, H.; Kobayashi, E.; et al. Genetic Mapping of Quantitative Trait Loci Affecting Body Weight, Egg Character and Egg Production in F2 Intercross Chickens. Anim. Genet. 2004, 35, 188–194. [Google Scholar] [CrossRef]
- Schreiweis, M.A.; Hester, P.Y.; Settar, P.; Moody, D.E. Identification of Quantitative Trait Loci Associated with Egg Quality, Egg Production, and Body Weight in an F2 Resource Population of Chickens. Anim. Genet. 2006, 37, 106–112. [Google Scholar] [CrossRef]
- Guo, J.; Qu, L.; Dou, T.-C.; Shen, M.-M.; Hu, Y.-P.; Ma, M.; Wang, K.-H. Genome-Wide Association Study Provides Insights into the Genetic Architecture of Bone Size and Mass in Chickens. Genome 2020, 63, 133–143. [Google Scholar] [CrossRef]
- Wolc, A.; Arango, J.; Settar, P.; Fulton, J.E.; O’Sullivan, N.P.; Preisinger, R.; Habier, D.; Fernando, R.; Garrick, D.J.; Hill, W.G.; et al. Genome-Wide Association Analysis and Genetic Architecture of Egg Weight and Egg Uniformity in Layer Chickens. Anim. Genet. 2012, 43, 87–96. [Google Scholar] [CrossRef]
- Barkova, O.Y.; Smaragdov, M.G. Association of a Nonsynonymous Substitution in the Condensin NCAPG Gene with Traits of Eggs in Laying Hens. Russ. J. Genet. Appl. Res. 2016, 6, 804–808. [Google Scholar] [CrossRef]
- Sun, C.; Qu, L.; Yi, G.; Yuan, J.; Duan, Z.; Shen, M.; Qu, L.; Xu, G.; Wang, K.; Yang, N. Genome-Wide Association Study Revealed a Promising Region and Candidate Genes for Eggshell Quality in an F2 Resource Population. BMC Genom. 2015, 16, 565. [Google Scholar] [CrossRef] [Green Version]
- Nangsuay, A.; Ruangpanit, Y.; Meijerhof, R.; Attamangkune, S. Yolk Absorption and Embryo Development of Small and Large Eggs Originating from Young and Old Breeder Hens. Poult. Sci. 2011, 90, 2648–2655. [Google Scholar] [CrossRef]
- Lourens, A.; van den Brand, H.; Meijerhof, R.; Kemp, B. Effect of Eggshell Temperature during Incubation on Embryo Development, Hatchability, and Posthatch Development. Poult. Sci. 2005, 84, 914–920. [Google Scholar] [CrossRef]
- Gaevsky, N.A. Acquaintance with Evolutionary Genetics; Krasnoyarsk State University: Krasnoyarsk, Russia, 2002. [Google Scholar]
- Dekkers, J. Commercial Application of Marker- and Gene-Assisted Selection in Livestock: Strategies and Lessons. J. Anim. Sci. 2004, 82, 313–328. [Google Scholar] [CrossRef]
- Boichard, D.; Chung, H.; Dassonneville, R.; David, X.; Eggen, A.; Fritz, S.; Gietzen, K.; Hayes, B.; Lawley, C.; Sonstegard, T.; et al. Design of a Bovine Low-Density SNP Array Optimized for Imputation. PLoS ONE 2012, 7, e34130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dementiva, N.V.; Kudinov, A.A.; Mitrofanova, O.V.; Mishina, A.I.; Smaragdov, M.G.; Yakovlev, A.F. WPSI-6 Chicken Resource Population as the Source of Study Genetic Improvement of Indigenous Breeds. J. Anim. Sci. 2018, 96, 513. [Google Scholar] [CrossRef]
- Moiseyeva, I.G.; Sevastyanova, A.A.; Aleksandrov, A.V.; Vakhrameev, A.B.; Romanov, M.N.; Dmitriev, Y.I.; Semenova, S.K.; Sulimova, G.E. Orloff Chicken Breed: History, Current Status and Studies. Izv. Timiryazev. S-Kh. Akad. (Proc. Timiryazev Agric. Acad.) 2016, (Issue 1), 78–96. [Google Scholar]
- Moiseyeva, I.G.; Nikiforov, A.A.; Romanov, M.N.; Aleksandrov, A.V.; Moysyak, Y.V.; Semyenova, S.K. Origin, History, Genetics and Economic Traits of the Yurlov Crower Chicken Breed. In Elektronnyi zhurnal (Electronic Journal); Laboratory of Animal Comparative Genetics, N.I. Vavilov Institute of General Genetics: Moscow, Russia, 2007; Available online: https://web.archive.org/web/20120210170800/http://www.lab-cga.ru/articles/Yurlovskaya/Yurlovskaya.htm (accessed on 9 July 2021).
- Moiseyeva, I.G.; Romanov, M.N.; Aleksandrov, A.V.; Nikiforov, A.A.; Sevastyanova, A.A. Evolution and Genetic Diversity of Old Domestic Hen’s Breed—Yurlovskaya Golosistaya: System Analysis of Variability Forms. Izv. Timiryazev. S-Kh. Akad. (Proc. Timiryazev Agric. Acad.) 2009, (Issue 3), 132–147. [Google Scholar]
- Romanov, M.N.; Bondarenko, Y.V. Introducing the Ukrainian Indigenous Poultry—The Poltava Chickens. Fancy Fowl 1994, 14(2), 8–9. [Google Scholar]
- Moiseyeva, I.G.; Kovalenko, A.T.; Mosyakina, T.V.; Romanov, M.N.; Bondarenko, Y.V.; Kutnyuk, P.I.; Podstreshny, A.P.; Nikiforov, A.A.; Tkachik, T.E. Origin, History, Genetics and Economic Traits of the Poltava Chicken Breed. In Elektronnyi zhurnal (Electronic Journal); Laboratory of Animal Comparative Genetics, N.I. Vavilov Institute of General Genetics: Moscow, Russia, 2006; Issue 4, Available online: https://web.archive.org/web/20120205195904/http://www.lab-cga.ru/articles/Jornal04/Statia2.htm (accessed on 9 July 2021).
- Moiseyeva, I.G.; Romanov, M.N.; Kovalenko, A.T.; Mosyakina, T.V.; Bondarenko, Y.V.; Kutnyuk, P.I.; Podstreshny, A.P.; Nikiforov, A.A. The Poltava Chicken Breed of Ukraine: Its History, Characterization and Conservation. Anim. Genet. Resour. Inf. 2007, 40, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Kulibaba, R.; Tereshchenko, A. Transforming Growth Factor β1, Pituitary-Specific Transcriptional Factor 1 and Insulin-Like Growth Factor I Gene Polymorphisms in the Population of the Poltava Clay Chicken Breed: Association with Productive Traits. Agric. Sci. Pract. 2015, 2, 67–72. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP | Position in GRCg6a Build [49] | Nearest Gene | Nucleotide Change | Location |
|---|---|---|---|---|
| GGaluGA265966 | 75,796,627 | NCAPG | A/G | Intergenic region |
| GGaluGA265969 | 75,827,200 | NCAPG LCORL | T/C | Intron |
| rs15619223 | 75,850,294 | NCAPG LCORL | A/C | Intron |
| rs14491017 | 75,885,777 | NCAPG | C/T | Intron |
| rs14491028 | 75,903,919 | NCAPG | C/T | Intron |
| No. of Animals | Chicken Groups | Egg-Type and Related | Dual Purpose and Related | Meat-Type and Related | Game | Fancy | Bantam and Related |
|---|---|---|---|---|---|---|---|
| 132 | Egg-type and related | (1.20) | 1.22 | 1.30 | 1.21 | 1.21 | 1.27 |
| 470 | Dual purpose and related | 0.0290 ± 0040 | (1.11) | 1.08 | 1.01 | 1.46 | 1.27 |
| 65 | Meat-type and related | 0.1490 ± 0.0522 | 0.0690 ± 0.0322 | (0.79) | 0.86 | 1.68 | 1.26 |
| 48 | Game | 0.0970 ± 0.0328 | 0.0290 ± 0.0130 | 0.0420 ± 0.0242 | (0.79) | 1.56 | 1.27 |
| 53 | Fancy | 0.1000 ± 0.0125 | 0.2190 ± 0.0258 | 0.3930 ± 0.0548 | 0.3210 ± 0.0325 | (0.84) | 1.30 |
| 92 | Bantam and related | 0.0160 ± 0.0124 | 0.0310 ± 0.0279 | 0.1200 ± 0.0526 | 0.1020 ± 0.0457 | 0.1420 ± 0.0493 | (1.27) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larkina, T.A.; Barkova, O.Y.; Peglivanyan, G.K.; Mitrofanova, O.V.; Dementieva, N.V.; Stanishevskaya, O.I.; Vakhrameev, A.B.; Makarova, A.V.; Shcherbakov, Y.S.; Pozovnikova, M.V.; et al. Evolutionary Subdivision of Domestic Chickens: Implications for Local Breeds as Assessed by Phenotype and Genotype in Comparison to Commercial and Fancy Breeds. Agriculture 2021, 11, 914. https://doi.org/10.3390/agriculture11100914
Larkina TA, Barkova OY, Peglivanyan GK, Mitrofanova OV, Dementieva NV, Stanishevskaya OI, Vakhrameev AB, Makarova AV, Shcherbakov YS, Pozovnikova MV, et al. Evolutionary Subdivision of Domestic Chickens: Implications for Local Breeds as Assessed by Phenotype and Genotype in Comparison to Commercial and Fancy Breeds. Agriculture. 2021; 11(10):914. https://doi.org/10.3390/agriculture11100914
Chicago/Turabian StyleLarkina, Tatyana A., Olga Y. Barkova, Grigoriy K. Peglivanyan, Olga V. Mitrofanova, Natalia V. Dementieva, Olga I. Stanishevskaya, Anatoly B. Vakhrameev, Alexandra V. Makarova, Yuri S. Shcherbakov, Marina V. Pozovnikova, and et al. 2021. "Evolutionary Subdivision of Domestic Chickens: Implications for Local Breeds as Assessed by Phenotype and Genotype in Comparison to Commercial and Fancy Breeds" Agriculture 11, no. 10: 914. https://doi.org/10.3390/agriculture11100914