Wild Relatives of Wheat Respond Well to Water Deficit Stress: A Comparative Study of Antioxidant Enzyme Activities and Their Encoding Gene Expression


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Procedures
2.2. Determination of Enzyme Activities
2.3. RNA Extraction and RT-qPCR Analyses
2.4. Data Analysis
3. Results
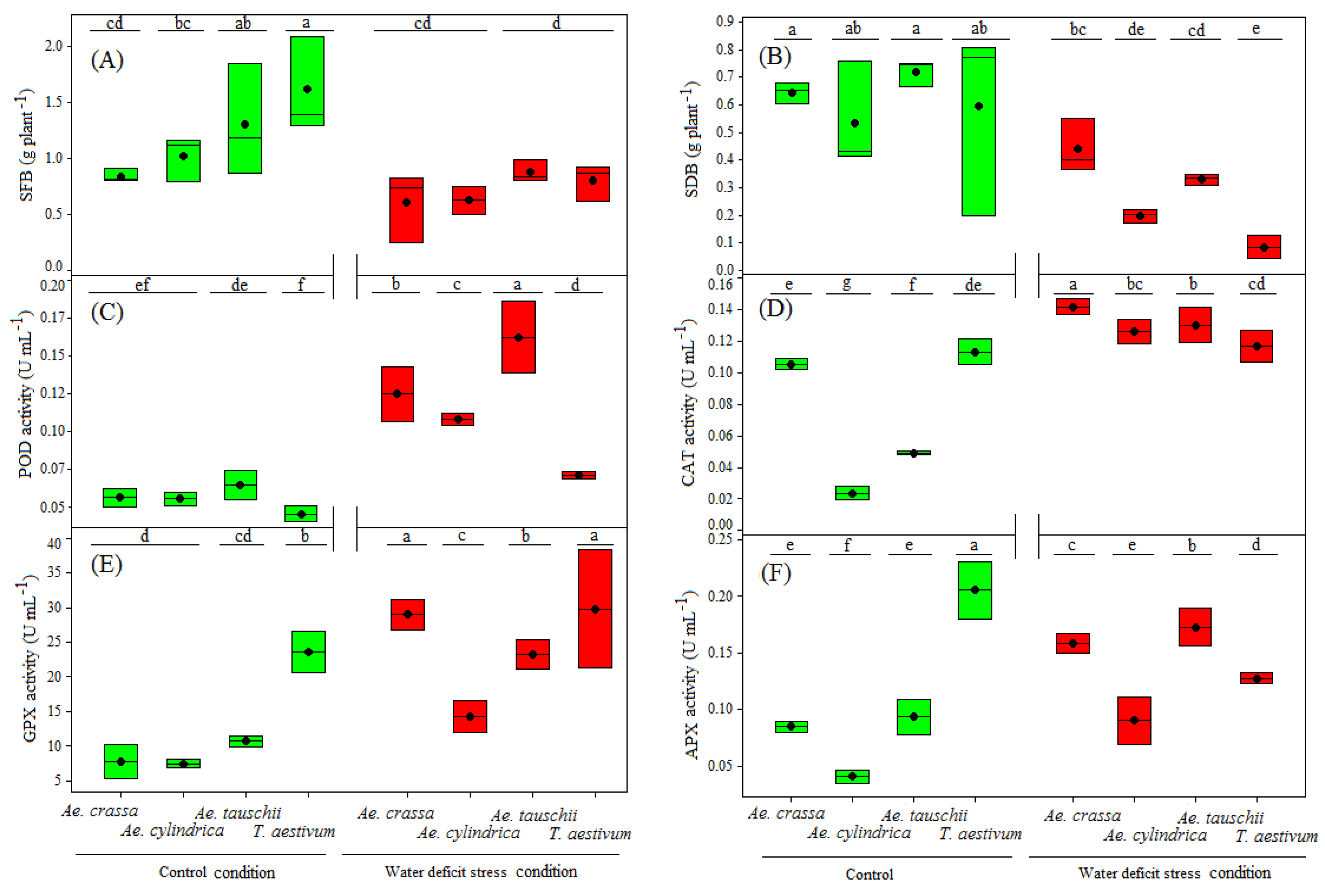
3.1. Effects of Water Deficit Treatments on Shoot Biomasses and Antioxidant Activities
3.2. Effects of Water Deficit Treatments on Biochemical Traits
3.3. Effects of Water Deficit Treatments on Gene Expression Profiles
3.4. Association between Shoot Biomasses with Biochemical Activities and Relative Gene Expressions
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shao, H.B.; Liang, Z.S.; Shao, M.A.; Sun, Q. Dynamic changes of antioxidative enzymes of ten wheat genotypes at soil water deficits. Colloids Surf. B Biointerf. 2005, 42, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H. Prospects for enhancement of the soluble antioxidants, ascorbate and glutathione. Biofactors 2001, 15, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.B.; Chu, L.Y.; Jaleel, C.A.; Zhao, C.X. Water-deficit stress-induced anatomical changes in higher plants. CR Biologies 2008, 331, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.S.; El Sayad, A.I.; Moore, M.; Dietz, K.J. Redox and reactive oxygen species network in acclimation for salinity tolerance in sugar beet. J. Exp. Bot. 2017, 68, 1283–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, Y.; Peng, Y.; Wang, X.; Peng, D.; Li, Y.; He, X.; Zhang, X.; Ma, X.; Huang, L.; et al. Clones of FeSOD, MDHAR, DHAR genes from white clover and gene expression analysis of ROS scavenging enzymes during abiotic stress and hormone treatments. Molecules 2015, 20, 20939–20954. [Google Scholar] [CrossRef]
- Hossain, M.S.; Dietz, K.J. Tuning of redox regulatory mechanisms, reactive oxygen species and redox homeostasis under salinity stress. Front. Plant Sci. 2016, 7, 548. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M. Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol. Adv. 2009, 27, 84–93. [Google Scholar] [CrossRef]
- Abedi, T.; Pakniyat, H. Antioxidant enzyme changes in response to drought stress in ten cultivars of oilseed rape (Brassica napus L.). Czech J. Plant Breed. 2010, 46, 27–34. [Google Scholar] [CrossRef]
- Akbari, M.; Katam, R.; Husain, R.; Farajpour, M.; Mazzuca, S.; Mahna, N. Sodium Chloride Induced Stress Responses of Antioxidative Activities in Leaves and Roots of Pistachio Rootstock. Biomolecules 2020, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, J.; Pour-Aboughadareh, A.; Fabriki-Ourang, S.; Khalili, P.; Poczai, P. Unravelling salinity stress responses in ancestral and neglected wheat species at early growth stage: A baseline for utilization in future wheat improvement programs. Physiol. Mol. Biol. Plants 2020, 26, 537–549. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Etminan, A.; Abdelrahman, M.; Siddique, K.; Tran, L.S.P. Assessment of biochemical and physiological parameters of durum wheat genotypes at the seedling stage duting polyethylene glycol-induced water stress. Plant Growth Regul. 2020. [Google Scholar] [CrossRef]
- Suneja, Y.; Gupta, A.K.; Bains, N.S. Bread wheat progenitors: Aegilops tauschii (DD genome) and Triticum dicoccoides (AABB genome) reveal differential antioxidative response under water stress. Physiol. Mol. Biol. Plants. 2017, 23, 99–114. [Google Scholar] [CrossRef]
- Suneja, Y.; Gupta, A.K.; Bains, N.S. Stress adaptive plasticity: Aegilops tauschii and Triticum dicoccoides as potential donors of drought associated morpho-physiological traits in Wheat. Front. Plant Sci. 2019, 10, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Bai, S.; Li, L.; Han, X.; Li, J.; Zhu, Y.; Fang, Y.; Zhang, D.; Li, S. Comparative transcriptome analysis of two Aegilops tauschii with contrasting drought tolerance by RNA-Seq. Int. J. Mol. Sci. 2020, 21, 3595. [Google Scholar] [CrossRef] [PubMed]
- Yousfi, S.; Marquez, A.J.; Betti, M. Gene expression and physiological responses to salinity and water stress of contrasting durum wheat genotypes. J. Integr. Plant Biol. 2016, 58, 48–66. [Google Scholar] [CrossRef]
- Blum, A. Drought resistance—Is it really a complex trait. Funct. Plant Biol. 2011, 38, 753–757. [Google Scholar] [CrossRef] [PubMed]
- Maxted, N. Crop Wild Relatives; Bioversity International: Rome, Italy, 2006. [Google Scholar]
- Maxted, N.; Ford-Lloyd, B.V.; Kell, S.P. Crop wild relatives: Establishing the context. In Crop Wild Relative Conservation and Use; Maxted, N., Ford-Lloyd, B.V., Kell, S.P., Iriondo, J., Dulloo, E., Turok, J., Eds.; CABI Publishing: Wallingford, UK, 2008; pp. 3–30. [Google Scholar]
- Maghsoudi, K.; Emam, Y.; Pessarakli, M. Effect of silicon on photosynthetic gas exchange, photosynthetic pigments, cell membrane stability and relative water content of different wheat cultivars under drought stress conditions. J. Plant Nutr. 2016, 39, 1001–1015. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. 2020. Available online: http://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 21 January 2020).
- Singh, P.; Mahajan, M.M.; Singh, N.K.; Kumar, D.; Kumar, K. Physiological and molecular response under salinity stress in bread wheat (Triticum aestivum L.). J. Plant Biochem. Biotehnol. 2020, 29, 125–133. [Google Scholar] [CrossRef]
- Kimber, G.; Feldman, M. Wild Wheat. An Introduction; Special Report No.353; College of Agriculture, University of Missouri-Columbia: Missouri, Columbia, 1987; p. 142. (In Columbia) [Google Scholar]
- Pour-Aboughadareh, A.; Ahmadi, J.; Mehrabi, A.A.; Etminan, A.; Moghaddam, M.; Siddique, K.H.M. Physiological responses to drought stress in wild relatives of wheat: Implications for wheat improvement. Acta Physiol. Plant. 2017, 39, 106. [Google Scholar] [CrossRef]
- Ahmadi, J.; Pour-Aboughadareh, A.; Fabriki Ourang, S.; Mehrabi, A.A.; Siddique, K.H.M. Wild relatives of wheat: Aegilops–Triticum accessions disclose differential antioxidative and physiological responses to water stress. Acta Physiol. Plant 2018, 40, 90. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Omidi, M.; Naghavi, M.R.; Etminan, A.; Mehrabi, A.A.; Poczai, P.; Bayat, H. Efect of water deficit stress on seedling biomass and physio-chemical characteristics in diferent species of wheat possessing the D genome. Agronomy 2019, 9, 522. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, J.; Pour-Aboughadareh, A.; Fabriki Ourang, S.; Mehrabi, A.A.; Siddique, K.H.M. Screening wheat germplasm for seedling root architectural traits under contrasting water regimes: Potential sources of variability for drought adaptation. Arch. Agron. Soil Sci. 2018, 64, 1351–1365. [Google Scholar] [CrossRef]
- Souza, C.C.; Oliveira, F.A.; Silva, I.F.; Amorim Neto, M.S. Evaluation of methods of available water determination and irrigation management in ‘‘terra roxa’’ under cotton crop. Rev. Bras. Eng. Agric. Ambient. 2000, 4, 338–342. [Google Scholar] [CrossRef] [Green Version]
- Pagariya, M.C.; Devarumath, R.M.; Kawar, P.G. Biochemical characterization and identification of differentially expressed candidate genes in salt stressed sugarcane. Plant Sci. 2012, 184, 1–13. [Google Scholar] [CrossRef]
- Manoranjan, K.; Dinabandhu, M. Catalase, peroxidase, and polyphenoloxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar]
- Hadwan, M. Simple spectrophotometric assay for measuring catalase activity in biological tissues. BMC Biochem. 2018, 19, 1–7. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, S.K. Assay of catalase and peroxidase. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Nakano, Y.; Asada, N.K. Hydrogen peroxide is scavenged by ascorbate-specifc peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867. [Google Scholar]
- Baek, K.H.; Skinner, D.Z. Alteration of antioxidant enzyme gene expression during cold acclimation of near-isogenic wheat lines. Plant Sci. 2003, 165, 1221–1227. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucl. Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- MSTAT-C. A Software Program for the Design, Management and Analysis of Agronomic Research Experiments; Michigan State University: East Lansing, MI, USA, 1991. [Google Scholar]
- Mammadov, J.; Buyyarapu, R.; Guttikonda, S.K.; Parliament, K.; Abdurakhmonov, I.Y.; Kumpatla, S.P. Wild relatives of maize, rice, cotton, and soybean: Treasure troves for tolerance to biotic and abiotic stresses. Front. Plant Sci. 2018, 9, 886. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Steffenson, B.J.; Sela, H.; Stinebaugh, K. Resistance of Aegilops longissima to the rusts of wheat. Plant Dis. 2018, 102, 1124–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivera, P.D.; Rouse, M.N.; Jin, Y. Identification of new sources of resistance to wheat stem rust in Aegilops spp. in the tertiary genepool of wheat. Front. Plant Sci. 2018, 9, 1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.M.; Doherty, S.J.; Croy, R.R.D. Biphasic superoxide generation in potato tubers. A self-amplifying response to stress. Plant Physiol. 2003, 13, 1440–1449. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.A.; Alqurainy, F. Activities of antioxidants in plants under environmental stress. In The Lutein-Prevention and Treatment for Diseases; Motohashi, N., Ed.; Transworld Research Network: Trivandrum, India, 2006; pp. 187–256. [Google Scholar]
- Feng, X.; Lai, Z.; Lin, Y.; Lai, G.; Lian, C. Genome-wide identification and characterization of the superoxide dismutase gene family in Musa acuminata cv. Tianbaojiao (AAA group). BMC Genom. 2015, 16, 823. [Google Scholar]
- Bowler, C.; Van Montagu, M.; Inze, D. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant. Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wu, C.; Xie, B.; Liu, Y.; Cui, J.; Chen, G.; Zhang, Y. Model analyzing the antioxidant responses of leaves and roots of switch grass to NaCl-salinity stress. Plant. Physiol. Biochem. 2012, 58, 288–296. [Google Scholar] [CrossRef]
- Xiao, X.; Xu, X.; Yang, F. Adaptive responses to progressive drought stress in two Populus cathayana populations. Silva Fennica 2008, 42, 705–719. [Google Scholar] [CrossRef] [Green Version]
- El-Esawi, M.; Al-Ghamdi, A.A.; Ali, H.M.; Ahmad, M. Overexpression of AtWRKY30 transcription factor enhances heat and drought stress tolerance in wheat (Triticum aestivum L.). Genes 2020, 10, 6968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Luo, X.; Nawaz, G.; Yin, J.; Yang, J. Physiological and biochemical responses of four cassava cultivars to drought stress. Sci. Rep. 2020, 10, 6968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Fartyal, D.; Agarwal, A.; Shukla, T.; James, D.; Kaul, T.; Reddy, M.K. Abiotic stress tolerance in plants: Myriad roles of ascorbate peroxidase. Front. Plant Sci. 2017, 8, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadi, J.; Pour-Aboughadareh, A.; Fabriki-Ourang, S.; Mehrabi, A.A.; Siddique, K.H.M. Screening wild progenitors of wheat for salinity stress at early stages of plant growth: Insight into potential sources of variability for salinity adaptation in wheat. Crop. Pasture Sci. 2018, 69, 649–658. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Khaishany, M.Y.; Al-Qutami, M.A.; Al-Whaibi, M.H.; Grover, A.; Ali, H.M.; Al-Wahibi, M.S. Morphological and physiological characterization of different genotypes of faba bean under heat stress. Saudi J. Biol. Sci. 2015, 22, 656–663. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′-3′) | |
|---|---|---|
| APX | Forward | GCAGCTGCTGAAGGAGAAGT |
| Reverse | CACTGGGGCCACTCACTAAT | |
| CAT | Forward | CCATGAGATCAAGGCCATCT |
| Reverse | ATCTTACATGCTCGGCTTGG | |
| MnSOD | Forward | CAGAGGGTGCTGCTTTACAA |
| Reverse | GGTCACAAGAGGGTCCTGAT | |
| GPX | Forward | CCCCCTGTACAAGTTCCTGA |
| Reverse | GTCAACAACGTGACCCTCCT | |
| 18S rRNA | Forward | GGCCGCTCCTAGCCCTAATTG |
| Reverse | TGAGCACTCTAATTTCTTCAAAGTACG |
| Source of Variation | Biomass | Antioxidant Activity | Relative Expression | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SFB | SDB | APX | POD | GPX | CAT | APXg | MnSODg | GPXg | CATg | |
| R | 0.076 ns | 0.021 ns | 0.001 ns | 0.0001 ns | 9.32 ns | 0.0001 ns | 8.294 ns | 1.144 ns | 0.994 ns | 0.214 ns |
| WT | 1.302 *** | 0.784 *** | 0.006 *** | 0.022 *** | 830.18 *** | 0.019 *** | 2671.142 *** | 2259.096 *** | 756.119 *** | 18.727 *** |
| A | 0.304 * | 0.067 * | 0.011 *** | 0.003 *** | 254.21 *** | 0.003 *** | 187.535 *** | 136.500 *** | 20.201 *** | 1.665 *** |
| Int. | 0.090 ns | 0.022ns | 0.008 *** | 0.001 *** | 72.55 * | 0.003 *** | 203.474 *** | 113.804 *** | 20.034 *** | 1.548 *** |
| MCC | 1.19 | 0.62 | 0.11 | 0.06 | 12.32 | 0.07 | 2.34 | 2.36 | 2.28 | 1.54 |
| MSC | 0.73 | 0.26 | 0.14 | 0.12 | 24.08 | 0.13 | 20.62 | 19.16 | 12.01 | 3.07 |
| †RC | 38.65 | 58.06 | −27.27 | −100 | −95.45 | −85.71 | 8.81-fold | 8.12-fold | 5.27-fold | 1.99-fold |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pour-Aboughadareh, A.; Omidi, M.; Naghavi, M.R.; Etminan, A.; Mehrabi, A.A.; Poczai, P. Wild Relatives of Wheat Respond Well to Water Deficit Stress: A Comparative Study of Antioxidant Enzyme Activities and Their Encoding Gene Expression. Agriculture 2020, 10, 415. https://doi.org/10.3390/agriculture10090415
Pour-Aboughadareh A, Omidi M, Naghavi MR, Etminan A, Mehrabi AA, Poczai P. Wild Relatives of Wheat Respond Well to Water Deficit Stress: A Comparative Study of Antioxidant Enzyme Activities and Their Encoding Gene Expression. Agriculture. 2020; 10(9):415. https://doi.org/10.3390/agriculture10090415
Chicago/Turabian StylePour-Aboughadareh, Alireza, Mansoor Omidi, Mohammad Reza Naghavi, Alireza Etminan, Ali Ashraf Mehrabi, and Peter Poczai. 2020. "Wild Relatives of Wheat Respond Well to Water Deficit Stress: A Comparative Study of Antioxidant Enzyme Activities and Their Encoding Gene Expression" Agriculture 10, no. 9: 415. https://doi.org/10.3390/agriculture10090415