Linking Endophytic Fungi to Medicinal Plants Therapeutic Activity. A Case Study on Asteraceae




Abstract
:1. Introduction
2. Asteraceae Ecology and Biochemistry
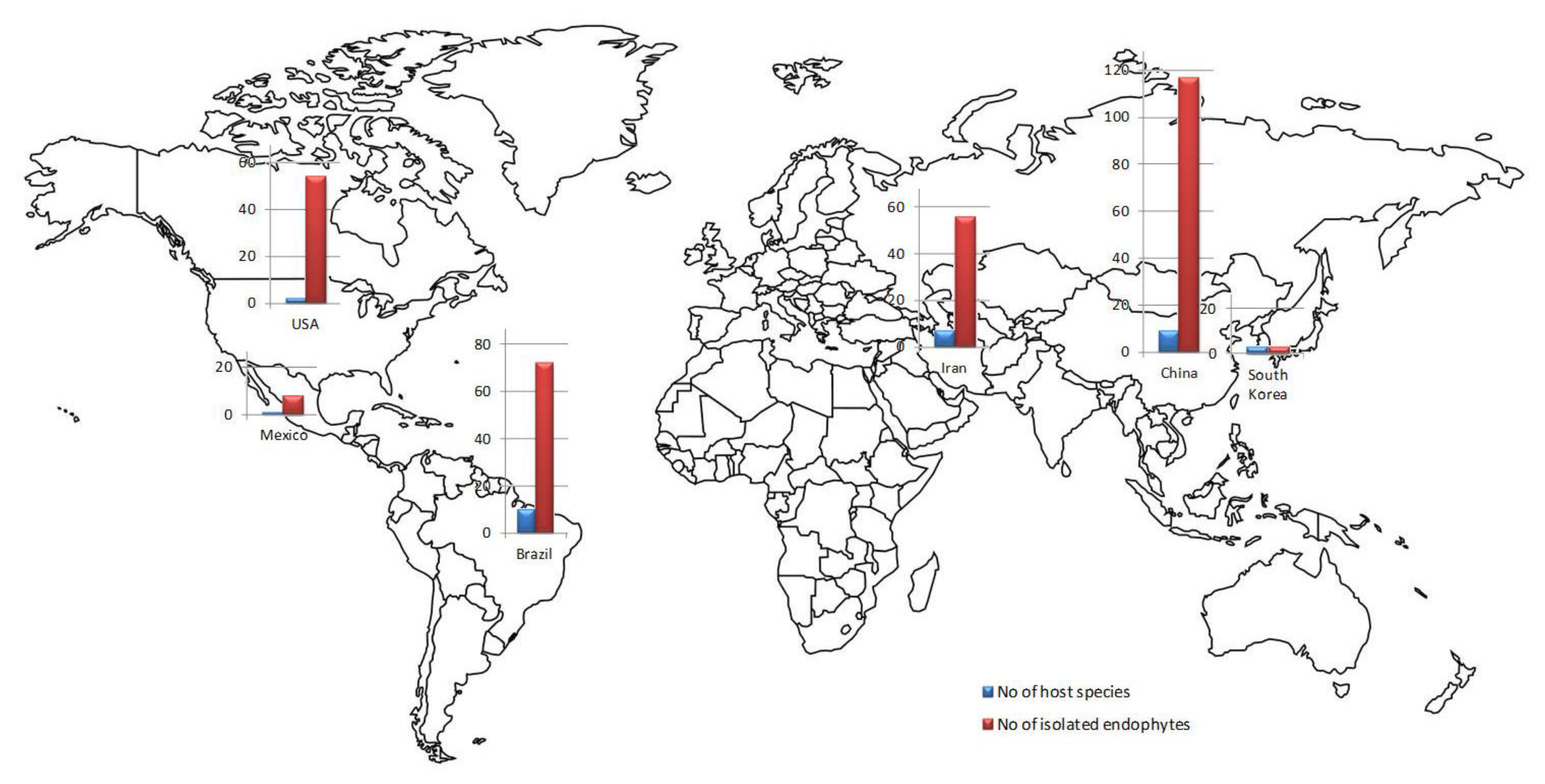
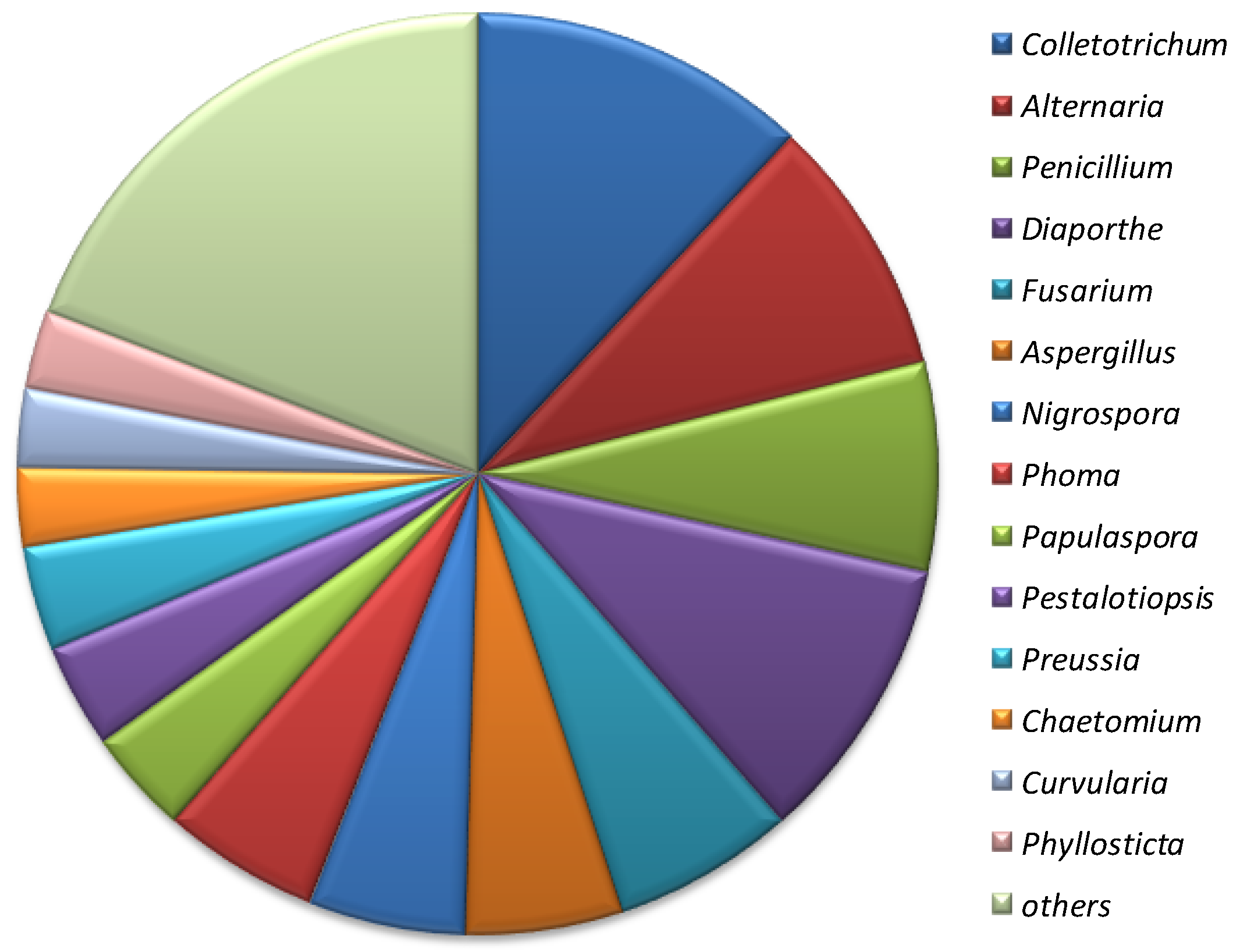
3. Fungal Endophytes Associated with Asteraceae—Biodiversity, and Ecology
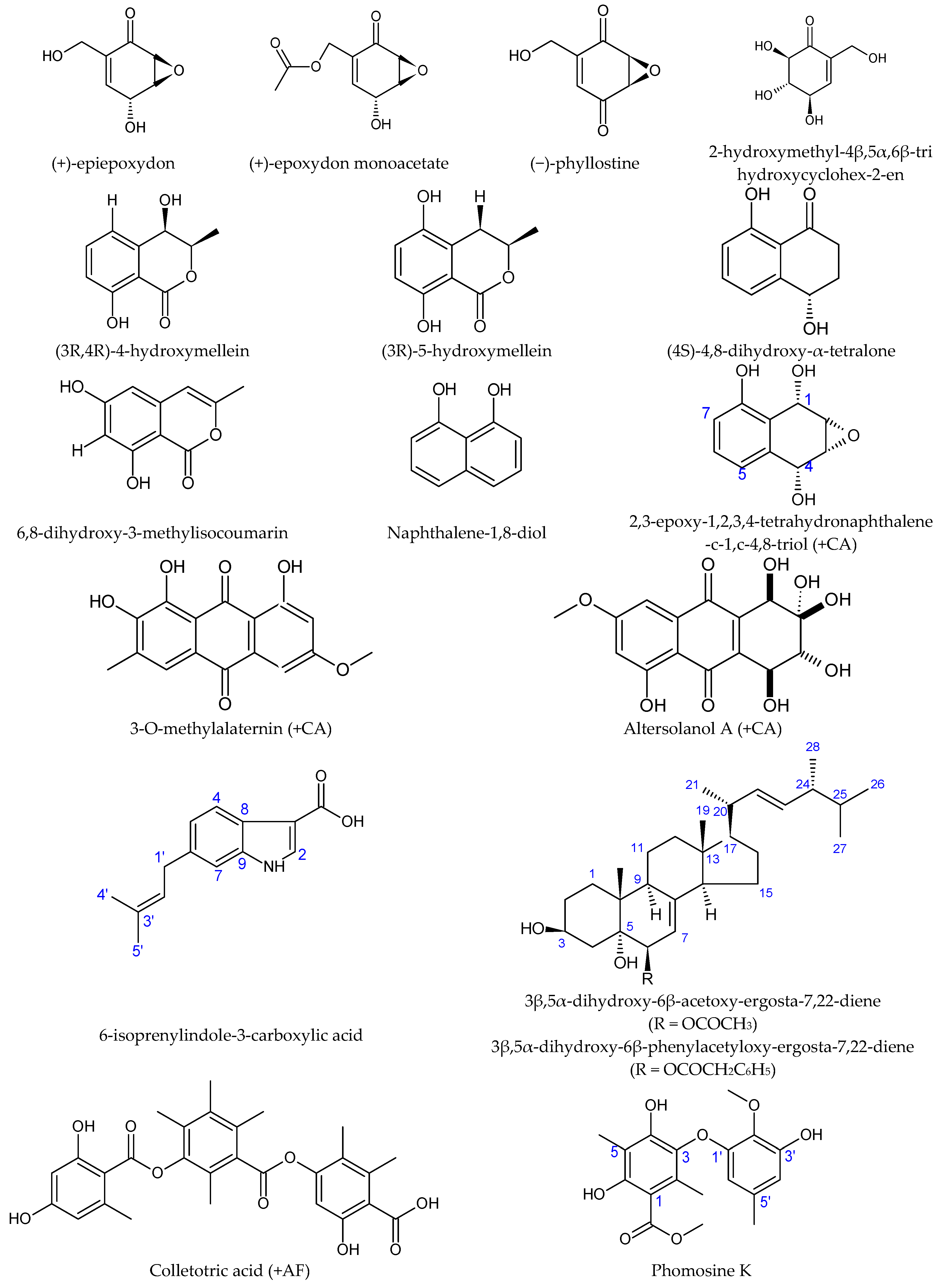
4. Fungal Endophytes Associated with Asteraceae—Biochemistry
4.1. Plant Growth Promoting Secondary and Anti-Stress Metabolites
4.2. Antibacterial Secondary Metabolites
4.3. Antifungal Secondary Metabolites
4.4. Antiparasitic Secondary Metabolites
4.5. Cytotoxic Secondary Metabolites
5. Review Methodology
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Aswani, R.; Vipina Vinod, T.; Ashitha, J. Benefits of plant–endophyte interaction for sustainable agriculture. In Microbial Endophytes: Functional Biology and Applications; Kumar, A., Radhakrishnan, E., Eds.; Elsevier INC.: Amsterdam, The Netherlands, 2020; pp. 35–55. [Google Scholar]
- Mani, V.M.; Soundari, A.P.G.; Karthiyaini, D.; Preethi, K. Bioprospecting endophytic fungi and their metabolites from medicinal tree Aegle marmelos in Western Ghats, India. Mycobiology 2015, 43, 303–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, D. Endophyte: The evolution of a term, and clarification of its use and definition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Golinska, P.; Wypij, M.; Agarkar, G.; Rathod, D.; Dahm, H.; Rai, M. Endophytic actinobacteria of medicinal plants: Diversity and bioactivity. Antonie Van Leeuwenhoek 2015, 108, 267–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez–Klimova, E.; Rodríguez–Peña, K.; Sánchez, S. Endophytes as sources of antibiotics. Biochem. Pharm. 2017, 134, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G.; Moreno–Hagelsieb, G.; del Carmen Orozco–Mosqueda, M.; Glick, B.R. Plant growth–promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Alvin, A.; Miller, K.I.; Neilan, B.A. Exploring the potential of endophytes from medicinal plants as sources of antimycobacterial compounds. Microbiol. Res. 2014, 169, 483–495. [Google Scholar] [CrossRef]
- Madhurama, G.; Nagma, S.; Preeti, S. Indole–3–acetic acid production by Streptomyces sp. isolated from rhizospheric soils of medicinal plants. J. Pure Appl. Microbiol. 2014, 8, 965–971. [Google Scholar]
- Aly, A.H.; Debbab, A.; Kjer, J.; Proksch, P. Fungal endophytes from higher plants: A prolific source of phytochemicals and other bioactive natural products. Fungal Diver. 2010, 41, 1–16. [Google Scholar] [CrossRef]
- Nicoletti, R.; Ferranti, P.; Caira, S.; Misso, G.; Castellano, M.; Di Lorenzo, G.; Caraglia, M. Myrtucommulone production by a strain of Neofusicoccum australe endophytic in myrtle (Myrtus communis). World J. Microbiol. Biotechnol. 2014, 30, 1047–1052. [Google Scholar] [CrossRef] [Green Version]
- Nicoletti, R.; Fiorentino, A. Plant bioactive metabolites and drugs produced by endophytic fungi of Spermatophyta. Agriculture 2015, 5, 918–970. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Shan, T.; Mou, Y.; Zhou, L. Plant–derived bioactive compounds produced by endophytic fungi. Mini-Rev. Med. Chem. 2011, 11, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Kusari, S.; Lamshöft, M.; Spiteller, M. Aspergillus fumigatus Fresenius, an endophytic fungus from Juniperus communis L. Horstmann as a novel source of the anticancer pro-drug deoxypodophyllotoxin. J. Appl. Microbiol. 2009, 107, 1019–1030. [Google Scholar] [CrossRef]
- Guimarães, D.O.; Lopes, N.P.; Pupo, M.T. Meroterpenes isolated from the endophytic fungus Guignardia mangiferae. Phytochem. Lett. 2012, 5, 519–523. [Google Scholar] [CrossRef]
- Nicoletti, R.; Ciavatta, M.L.; Buommino, E.; Tufano, M.A. Antitumor extrolites produced by Penicillium species. Int. J. Biomed. Pharm. Sci. 2008, 2, 1–23. [Google Scholar]
- Stammati, A.; Nicoletti, R.; De Stefano, S.; Zampaglioni, F.; Zucco, F. Cytostatic properties of a novel compound derived from Penicillium pinophilum: An in vitro study. ALTA Alter. Lab. Anim. 2002, 30, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Rosenblueth, M.; Martínez–Romero, E. Bacterial endophytes and their interactions with hosts. Mol. Plant–Microbe Interact. 2006, 19, 827–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsitsigiannis, D.I.; Keller, N.P. Oxylipins as developmental and host–fungal communication signals. Trends Microbiol. 2007, 15, 109–118. [Google Scholar] [CrossRef]
- Yang, Z.; Rogers, L.M.; Song, Y.; Guo, W.; Kolattukudy, P. Homoserine and asparagine are host signals that trigger in planta expression of a pathogenesis gene in Nectria haematococca. Proc. Natl. Acad. Sci. USA 2005, 102, 4197–4202. [Google Scholar] [CrossRef] [Green Version]
- Young, C.A.; Felitti, S.; Shields, K.; Spangenberg, G.; Johnson, R.D.; Bryan, G.T.; Saikia, S.; Scott, B. A complex gene cluster for indole–diterpene biosynthesis in the grass endophyte Neotyphodium lolii. Fungal Gen. Biol. 2006, 43, 679–693. [Google Scholar] [CrossRef]
- Schroeckh, V.; Scherlach, K.; Nützmann, H.-W.; Shelest, E.; Schmidt-Heck, W.; Schuemann, J.; Martin, K.; Hertweck, C.; Brakhage, A.A. Intimate bacterial–fungal interaction triggers biosynthesis of archetypal polyketides in Aspergillus nidulans. Proc. Natl. Acad. Sci. USA 2009, 106, 14558–14563. [Google Scholar] [CrossRef] [Green Version]
- Schulz, B.; Haas, S.; Junker, C.; Andrée, N.; Schobert, M. Fungal endophytes are involved in multiple balanced antagonisms. Curr. Sci. 2015, 109, 39–45. [Google Scholar]
- Mazur, S.; Nadziakiewicz, M.; Kurzawińska, H.; Nawrocki, J. Effectiveness of mycorrhizal fungi in the protection of juniper, rose, yew and highbush blueberry against Alternaria alternata. Folia Hort. 2019, 31, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Jiang, D.; Wei, X. Mutation breeding of Emericella foeniculicola TR21 for improved production of tanshinone IIA. Process Biochem. 2011, 46, 2059–2063. [Google Scholar] [CrossRef]
- Zhao, K.; Sun, L.; Ma, X.; Li, X.; Wang, X.; Ping, W.; Zhou, D. Improved taxol production in Nodulisporium sylviforme derived from inactivated protoplast fusion. Afr. J. Biotechnol. 2013, 10, 4175–4182. [Google Scholar]
- Karuppusamy, S. A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. J. Med. Plants Res. 2009, 3, 1222–1239. [Google Scholar]
- Kusari, S.; Singh, S.; Jayabaskaran, C. Rethinking production of Taxol (paclitaxel) using endophyte biotechnology. Trends Biotechnol. 2014, 32, 304–311. [Google Scholar] [CrossRef]
- Yang, H.; Wang, Y.; Zhang, Z.; Yan, R.; Zhu, D. Whole-genome shotgun assembly and analysis of the genome of Shiraia sp. strain Slf14, a novel endophytic fungus producing huperzine A and hypocrellin A. Genome Announc. 2014, 2, e00011–e00014. [Google Scholar] [CrossRef] [Green Version]
- Venugopalan, A.; Srivastava, S. Endophytes as in vitro production platforms of high value plant secondary metabolites. Biotechnol. Adv. 2015, 33, 873–887. [Google Scholar] [CrossRef] [PubMed]
- Chutulo, E.C.; Chalannavar, R.K. Endophytic mycoflora and their bioactive compounds from Azadirachta indica: A comprehensive review. J. Fungi 2018, 4, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, D.; Pramanik, A.; Agrawal, P.K. Evaluation of bioactive secondary metabolites from endophytic fungus Pestalotiopsis neglecta BAB-5510 isolated from leaves of Cupressus torulosa D. Don. 3 Biotech 2016, 6, 210. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Zhang, L.; Li, L.; Zheng, C.; Guo, L.; Li, W.; Sun, P.; Qin, L. Recent developments and future prospects of antimicrobial metabolites produced by endophytes. Microbiol. Res. 2010, 165, 437–449. [Google Scholar] [CrossRef]
- Karsch-Völk, M.; Barrett, B.; Kiefer, D.; Bauer, R.; Ardjomand-Woelkart, K.; Linde, K. Echinacea for preventing and treating the common cold. Cochrane Database Syst. Rev. 2014, 2, CD000530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Plan List. Available online: www.theplantlist.org (accessed on 12 June 2020).
- Bessada, S.M.; Barreira, J.C.; Oliveira, M.B.P. Asteraceae species with most prominent bioactivity and their potential applications: A review. Ind. Crop. Prod. 2015, 76, 604–615. [Google Scholar] [CrossRef]
- Funk, V.A.; Bayer, R.J.; Keeley, S.; Chan, R.; Watson, L.; Gemeinholzer, B.; Schilling, E.; Panero, J.L.; Baldwin, B.G.; Garcia-Jacas, N. Everywhere but Antarctica: Using a supertree to understand the diversity and distribution of the Compositae. Biol. Skr. 2005, 55, 343–374. [Google Scholar]
- Petkova, N.T.; Ivanov, I.; Raeva, M.; Topuzova, M.; Todorova, M.; Denev, P. Fructans and antioxidants in leaves of culinary herbs from Asteraceae and Amaryllidaceae families. Food Res. 2019, 3, 407–415. [Google Scholar] [CrossRef]
- Martucci, M.E.P.; De Vos, R.C.; Carollo, C.A.; Gobbo-Neto, L. Metabolomics as a potential chemotaxonomical tool: Application in the genus Vernonia Schreb. PLoS ONE 2014, 9, e93149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lusa, M.G.; Martucci, M.E.; Loeuille, B.F.; Gobbo-Neto, L.; Appezzato-da-Glória, B.; Da Costa, F.B. Characterization and evolution of secondary metabolites in Brazilian Vernonieae (Asteraceae) assessed by LC–MS fingerprinting. Bot. J. Linn. Soc. 2016, 182, 594–611. [Google Scholar] [CrossRef]
- Gallon, M.E.; Jaiyesimi, O.A.; Gobbo-Neto, L. LC–UV–HRMS dereplication of secondary metabolites from Brazilian Vernonieae (Asteraceae) species supported through in–house database. Biochem. Syst. Ecol. 2018, 78, 5–16. [Google Scholar] [CrossRef]
- Mandel, J.R.; Barker, M.S.; Bayer, R.J.; Dikow, R.B.; Gao, T.G.; Jones, K.E.; Keeley, S.; Kilian, N.; Ma, H.; Siniscalchi, C.M. The Compositae tree of life in the age of phylogenomics. J. Syst. Evol. 2017, 55, 405–410. [Google Scholar] [CrossRef]
- Katinas, L.; Funk, V.A. An updated classification of the basal grade of Asteraceae (=Compositae): From Cabrera’s 1977 tribe Mutisieae to the present. N. Z. J. Bot. 2020, 58, 67–93. [Google Scholar] [CrossRef]
- Morales, P.; Ferreira, I.C.; Carvalho, A.M.; Sánchez-Mata, M.C.; Cámara, M.; Fernández-Ruiz, V.; Pardo-de-Santayana, M.; Tardío, J. Mediterranean non–cultivated vegetables as dietary sources of compounds with antioxidant and biological activity. LWT–Food Sci. Technol. 2014, 55, 389–396. [Google Scholar] [CrossRef] [Green Version]
- Zou, W.; Meng, J.; Lu, H.; Chen, G.; Shi, G.; Zhang, T.; Tan, R. Metabolites of Colletotrichum gloeosporioides, an endophytic fungus in Artemisia mongolica. J. Nat. Prod. 2000, 63, 1529–1530. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, R.; Coppo, E.; Marchese, A.; Daglia, M.; Sobarzo-Sánchez, E.; Nabavi, S.F.; Nabavi, S.M. Phytochemicals for human disease: An update on plant–derived compounds antibacterial activity. Microbiol. Res. 2017, 196, 44–68. [Google Scholar] [CrossRef]
- Alvarenga, S.; Ferreira, M.; Emerenciano, V.d.P.; Cabrol-Bass, D. Chemosystematic studies of natural compounds isolated from Asteraceae: Characterization of tribes by principal component analysis. Chemom. Intell. Lab. Syst. 2001, 56, 27–37. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853. [Google Scholar] [CrossRef]
- Rosa, L.H.; Gonçalves, V.N.; Caligiorne, R.B.; Alves, T.; Rabello, A.; Sales, P.A.; Romanha, A.J.; Sobral, M.E.; Rosa, C.A.; Zani, C.L. Leishmanicidal, trypanocidal, and cytotoxic activities of endophytic fungi associated with bioactive plants in Brazil. Braz. J. Microbiol. 2010, 41, 420–430. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, R.M.; Herrera, P.; Furrazola, E. Arbuscular mycorrhizal colonization in Asteraceae from white sand savannas, in Pinar del Río, Cuba. Biota Neotrop. 2013, 13, 136–140. [Google Scholar] [CrossRef]
- Reis, I.M.A.; Ribeiro, F.P.C.; Almeida, P.R.M.; Costa, L.C.B.; Kamida, H.M.; Uetanabaro, A.P.T.; Branco, A. Characterization of the secondary metabolites from endophytic fungi Nodulisporium sp. isolated from the medicinal plant Mikania laevigata (Asteraceae) by reversed–phase high–performance liquid chromatography coupled with mass spectrometric multistage. Pharmacogn. Mag. 2018, 14, 495. [Google Scholar]
- Zhao, K.; Penttinen, P.; Guan, T.; Xiao, J.; Chen, Q.; Xu, J.; Lindström, K.; Zhang, L.; Zhang, X.; Strobel, G.A. The diversity and anti–microbial activity of endophytic actinomycetes isolated from medicinal plants in Panxi plateau, China. Curr. Microbiol. 2011, 62, 182–190. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.M.; Zhang, X.P.; Zhao, K.; Zhong, K.; Gu, Y.F.; Lindström, K. Genetic characterisation of endophytic actinobacteria isolated from the medicinal plants in Sichuan. Ann. Microbiol. 2008, 58, 597–604. [Google Scholar] [CrossRef]
- Vieira, M.L.; Johann, S.; Hughes, F.M.; Rosa, C.A.; Rosa, L.H. The diversity and antimicrobial activity of endophytic fungi associated with medicinal plant Baccharis trimera (Asteraceae) from the Brazilian savannah. Can. J. Microbiol. 2014, 60, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Jeewon, R.; Liew, E.C.; Simpson, J.A.; Hodgkiss, I.J.; Hyde, K.D. Phylogenetic significance of morphological characters in the taxonomy of Pestalotiopsis species. Mol. Phylogen. Evol. 2003, 27, 372–383. [Google Scholar] [CrossRef]
- Carvalho, C.R.; Gonçalves, V.N.; Pereira, C.B.; Johann, S.; Galliza, I.V.; Alves, T.M.; Rabello, A.; Sobral, M.E.; Zani, C.L.; Rosa, C.A. The diversity, antimicrobial and anticancer activity of endophytic fungi associated with the medicinal plant Stryphnodendron adstringens (Mart.) Coville (Fabaceae) from the Brazilian savannah. Symbiosis 2012, 57, 95–107. [Google Scholar] [CrossRef]
- Hatamzadeh, S.; Rahnama, K.; Nasrollahnejad, S.; Fotouhifar, K.B.; Hemmati, K.; White, J.F.; Taliei, F. Isolation and identification of L–asparaginase–producing endophytic fungi from the Asteraceae family plant species of Iran. PeerJ 2020, 8, e8309. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Tian, Z.; Feng, L.; Xu, L.; Wang, H. Diversity analysis of the rhizospheric and endophytic bacterial communities of Senecio vulgaris L.(Asteraceae) in an invasive range. PeerJ 2019, 6, e6162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.-Y.; Cai, Y.-Z.; Xing, J.; Corke, H.; Sun, M. A potential antioxidant resource: Endophytic fungi from medicinal plants. Econ. Bot. 2007, 61, 14. [Google Scholar] [CrossRef]
- Christian, N.; Sullivan, C.; Visser, N.D.; Clay, K. Plant host and geographic location drive endophyte community composition in the face of perturbation. Microb. Ecol. 2016, 72, 621–632. [Google Scholar] [CrossRef]
- Jiang, H.; Shi, Y.-T.; Zhou, Z.-X.; Yang, C.; Chen, Y.-J.; Chen, L.-M.; Yang, M.-Z.; Zhang, H.-B. Leaf chemistry and co–occurring species interactions affecting the endophytic fungal composition of Eupatorium adenophorum. Ann. Microbiol. 2011, 61, 655–662. [Google Scholar] [CrossRef]
- Mei, L.; Zhu, M.; Zhang, D.-Z.; Wang, Y.-Z.; Guo, J.; Zhang, H.-B. Geographical and temporal changes of foliar fungal endophytes associated with the invasive plant Ageratina adenophora. Microb. Ecol. 2014, 67, 402–409. [Google Scholar] [CrossRef]
- Fang, K.; Miao, Y.-F.; Chen, L.; Zhou, J.; Yang, Z.-P.; Dong, X.-F.; Zhang, H.-B. Tissue–specific and geographical variation in endophytic fungi of Ageratina adenophora and fungal associations with the environment. Front. Microbiol. 2019, 10, 2919. [Google Scholar] [CrossRef] [Green Version]
- Pereira, J.M.; Peixoto, V.; Teixeira, A.; Sousa, D.; Barros, L.; Ferreira, I.C.; Vasconcelos, M.H. Achillea millefolium L. hydroethanolic extract has phenolic compounds and inhibits the growth of human tumor cell lines. Free Radic. Biol. Med. 2018, 120, S145. [Google Scholar] [CrossRef]
- Moghadam, M.H.; Mosaddegh, M.; Irani, M. Programmed cell death in breast adeno–carcinoma induced by Achillea filipendulina. Med. Plants–Int. J. Phytomed. Relat. Ind. 2019, 11, 435–439. [Google Scholar] [CrossRef]
- Aminkhani, A.; Sharifi, S.; Ekhtiyari, S. Achillea filipendulina Lam.: Chemical constituents and antimicrobial activities of essential oil of stem, leaf, and flower. Chem. Biodivers. 2020, 17, e2000133. [Google Scholar] [CrossRef] [PubMed]
- Boukhary, R.; Aboul-ElA, M.; El-Lakany, A. Review on chemical constituents and biological activities of genus Anthemis. Pharm. J. 2019, 11, 1155–1166. [Google Scholar] [CrossRef] [Green Version]
- Buommino, E.; Baroni, A.; Canozo, N.; Petrazzuolo, M.; Nicoletti, R.; Vozza, A.; Tufano, M.A. Artemisinin reduces human melanoma cell migration by down–regulating αvβ3 integrin and reducing metalloproteinase 2 production. Investig. New Drugs 2009, 27, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Kontogianni, V.G.; Primikyri, A.; Sakka, M.; Gerothanassis, I.P. Simultaneous determination of artemisinin and its analogs and flavonoids in Artemisia annua crude extracts with the use of NMR spectroscopy. Magn. Reson. Chem. 2020, 58, 232–244. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Desta, K.T.; Kim, G.S.; Lee, S.J.; Lee, W.S.; Kim, Y.H.; Jin, J.S.; Abd El-Aty, A.; Shin, H.C.; Shim, J.H. Polyphenolic profile and antioxidant effects of various parts of Artemisia annua L. Biomed. Chromatogr. 2016, 30, 588–595. [Google Scholar] [CrossRef]
- Lu, H.; Zou, W.X.; Meng, J.C.; Hu, J.; Tan, R.X. New bioactive metabolites produced by Colletotrichum sp., an endophytic fungus in Artemisia annua. Plant Sci. 2000, 151, 67–73. [Google Scholar] [CrossRef]
- Abiri, R.; Silva, A.L.M.; de Mesquita, L.S.S.; de Mesquita, J.W.C.; Atabaki, N.; de Almeida, E.B., Jr.; Shaharuddin, N.A.; Malik, S. Towards a better understanding of Artemisia vulgaris: Botany, phytochemistry, pharmacological and biotechnological potential. Food Res. Int. 2018, 109, 403–415. [Google Scholar] [CrossRef]
- Lösgen, S.; Magull, J.; Schulz, B.; Draeger, S.; Zeeck, A. Isofusidienols: Novel chromone-3-oxepines produced by the endophytic fungus Chalara sp. Eur. J. Org. Chem. 2008, 2008, 698–703. [Google Scholar] [CrossRef]
- Pandey, A.K.; Singh, P. The genus Artemisia: A 2012–2017 literature review on chemical composition, antimicrobial, insecticidal and antioxidant activities of essential oils. Medicines 2017, 4, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.; Jiang, J.-S.; Feng, Z.-M.; Yang, Y.-N.; Li, L.; Zang, C.-X.; Zhang, P.-C. Bioactive sesquiterpenoid and polyacetylene glycosides from Atractylodes lancea. J. Nat. Prod. 2016, 79, 1567–1575. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.-G.; Dai, C.-C. Jasmonic acid is involved in the signaling pathway for fungal endophyte–induced volatile oil accumulation of Atractylodes lancea plantlets. BMC Plant Biol. 2012, 12, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheriyan, B.V.; Joshi, S.; Mohamed, S. Eupatorium triplinerve (Vahl): An ethnobotanical review. Asian J. Pharm. Res. 2019, 9, 200–202. [Google Scholar] [CrossRef]
- Da Silva Paes, L.; de Lucena, J.M.V.M.; da Silva Bentes, J.L.; de Oliveira Marques, J.D.; Casas, L.L.; Mendonca, M.S. Endophytic mycobiota of three Amazonian medicinal herbs: Stachytarpheta cayennensis (Verbenaceae), Ayapana triplinervis (Asteraceae) and Costus spicatus (Costaceae). Int. J. Bot. 2014, 10, 24. [Google Scholar]
- Bonin, E.; Carvalho, V.M.; Avila, V.D.; dos Santos, N.C.A.; Benassi–Zanqueta, É.; Lancheros, C.A.C.; Previdelli, I.T.S.; Ueda–Nakamura, T.; de Abreu Filho, B.A.; do Prado, I.N. Baccharis dracunculifolia: Chemical constituents, cytotoxicity and antimicrobial activity. LWT 2020, 120, 108920. [Google Scholar] [CrossRef]
- Cuzzi, C.; Link, S.; Vilani, A.; Sartori, C.; Onofre, S.B. Endophytic fungi of the “vassourinha” (Baccharis dracunculifolia DC, Asteraceae). Rev. Bras. Bioc. 2012, 10, 135. [Google Scholar]
- Gonçalves, L.; Tavares, B.; Galdino, F.; Andrade, J.; Abrantes, L.; De Sousa, M.R.; Lira, P.G.; Pereira, L.R. Carqueja: Chemical composition and pharmacological effects. In Proceedings of the MOL2NET 2018, International Conference on Multidisciplinary Sciences, Paraiba, Brazil, 25 October 2018; p. 1. [Google Scholar]
- Xuan, T.D.; Khanh, T.D. Chemistry and pharmacology of Bidens pilosa: An overview. J. Pharm. Investig. 2016, 46, 91–132. [Google Scholar] [CrossRef]
- Abdou, R.; Scherlach, K.; Dahse, H.-M.; Sattler, I.; Hertweck, C. Botryorhodines A–D, antifungal and cytotoxic depsidones from Botryosphaeria rhodina, an endophyte of the medicinal plant Bidens pilosa. Phytochemistry 2010, 71, 110–116. [Google Scholar] [CrossRef]
- Saeed, M.; Abd El-Hack, M.E.; Alagawany, M.; Arain, M.A.; Arif, M.; Mirza, M.A.; Naveed, M.; Chao, S.; Sarwar, M.; Sayab, M. Chicory (Cichorium intybus) herb: Chemical composition, pharmacology, nutritional and healthical applications. Int. J. Pharm. 2017, 13, 351–360. [Google Scholar]
- Siriwatanametanon, N.; Heinrich, M. The Thai medicinal plant Gynura pseudochina var. hispida: Chemical composition and in vitro NF–κB inhibitory activity. Nat. Prod. Commun. 2011, 6, 627–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siriwach, R.; Kinoshita, H.; Kitani, S.; Igarashi, Y.; Pansuksan, K.; Panbangred, W.; Nihira, T. Bipolamides A and B, triene amides isolated from the endophytic fungus Bipolaris sp. MU34. J. Antibiot. 2014, 67, 167–170. [Google Scholar] [CrossRef]
- Akpor, O.; Olaolu, T.; Rotimi, D. Antibacterial and antioxidant potentials of leave extracts of Helianthus annuus. Potrav. Slovak J. Food Sci. 2019, 13, 1026–1033. [Google Scholar] [CrossRef] [Green Version]
- Al-Snafi, A.E. The pharmacological effects of Helianthus annuus—A review. Indo Am. J. Pharm. Sci. 2018, 5, 1745–1756. [Google Scholar]
- Waqas, M.; Khan, A.L.; Hamayun, M.; Shahzad, R.; Kang, S.-M.; Kim, J.-G.; Lee, I.-J. Endophytic fungi promote plant growth and mitigate the adverse effects of stem rot: An example of Penicillium citrinum and Aspergillus terreus. J. Plant Int. 2015, 10, 280–287. [Google Scholar]
- Waqas, M.; Khan, A.L.; Hamayun, M.; Shahzad, R.; Kim, Y.-H.; Choi, K.-S.; Lee, I.-J. Endophytic infection alleviates biotic stress in sunflower through regulation of defence hormones, antioxidants and functional amino acids. Eur. J. Plant Pathol. 2015, 141, 803–824. [Google Scholar] [CrossRef]
- Getahun, T.; Sharma, V.; Gupta, N. The genus Laggera (Asteraceae)–Ethnobotanical and ethnopharmacological information, chemical composition as well as biological activities of its essential oils and extracts: A review. Chem. Biodiver. 2019, 16, e1900131. [Google Scholar] [CrossRef] [PubMed]
- Matasyoh, J.C.; Dittrich, B.; Schueffler, A.; Laatsch, H. Larvicidal activity of metabolites from the endophytic Podospora sp. against the malaria vector Anopheles gambiae. Parasitol. Res. 2011, 108, 561–566. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, P. Complementary, holistic, and integrative medicine: Chamomile. Pediatr. Rev. 2007, 28, e16. [Google Scholar] [CrossRef]
- Della Pasqua, C.; Iwamoto, R.; Antunes, E.; Borghi, A.; Sawaya, A.; Landucci, E. Pharmacological study of anti–inflammatory activity of aqueous extracts of Mikania glomerata (Spreng.) and Mikania laevigata (Sch. Bip. ex Baker). J. Ethnopharm. 2019, 231, 50–56. [Google Scholar] [CrossRef]
- Polonio, J.; Almeida, T.; Garcia, A.; Mariucci, G.; Azevedo, J.; Rhoden, S.; Pamphile, J. Biotechnological prospecting of foliar endophytic fungi of guaco (Mikania glomerata Spreng.) with antibacterial and antagonistic activity against phytopathogens. Gen. Mol. Res. 2015, 14, 7297–7309. [Google Scholar] [CrossRef] [PubMed]
- Azab, A. A facile method for testing antioxidant capacity and total phenolic content of Notobasis syriaca and Scolymus maculatus extracts and their antifungal activity. Eur. Chem. Bull. 2018, 7, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Hussain, H.; Tchimene, M.K.; Ahmed, I.; Meier, K.; Steinert, M.; Draeger, S.; Schulz, B.; Krohn, K. Antimicrobial chemical constituents from the endophytic fungus Phomopsis sp. from Notobasis syriaca. Nat. Prod. Commun. 2011, 6, 1905–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, M.; Xu, Y.; Rehman, Z.U.; Arain, M.A.; Soomro, R.N.; Abd El-Hack, M.E.; Bhutto, Z.A.; Abbasi, B.; Dhama, K.; Sarwar, M. Nutritional and healthical aspects of yacon (Smallanthus sonchifolius) for human, animals and poultry. Int. J. Pharm. 2017, 13, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Gallo, M.B.; Chagas, F.O.; Almeida, M.O.; Macedo, C.C.; Cavalcanti, B.C.; Barros, F.W.; de Moraes, M.O.; Costa-Lotufo, L.V.; Pessoa, C.; Bastos, J.K. Endophytic fungi found in association with Smallanthus sonchifolius (Asteraceae) as resourceful producers of cytotoxic bioactive natural products. J. Basic Microbiol. 2009, 49, 142–151. [Google Scholar] [CrossRef]
- Gallo, M.B.; Cavalcanti, B.C.; Barros, F.W.; Odorico de Moraes, M.; Costa-Lotufo, L.V.; Pessoa, C.; Bastos, J.K.; Pupo, M.T. Chemical constituents of Papulaspora immersa, an endophyte from Smallanthus sonchifolius (Asteraceae), and their cytotoxic activity. Chem. Biodivers. 2010, 7, 2941–2950. [Google Scholar] [CrossRef]
- Chagas, F.O.; Dias, L.G.; Pupo, M.T. A mixed culture of endophytic fungi increases production of antifungal polyketides. J. Chem. Ecol. 2013, 39, 1335–1342. [Google Scholar] [CrossRef]
- Rosa, L.H.; Queiroz, S.C.; Moraes, R.M.; Wang, X.; Techen, N.; Pan, Z.; Cantrell, C.L.; Wedge, D.E. Coniochaeta ligniaria: Antifungal activity of the cryptic endophytic fungus associated with autotrophic tissue cultures of the medicinal plant Smallanthus sonchifolius (Asteraceae). Symbiosis 2013, 60, 133–142. [Google Scholar] [CrossRef]
- Porwal, O.; Ameen, M.S.M.; Anwer, E.T.; Uthirapathy, S.; Ahamad, J.; Tahsin, A. Silybum marianum (Milk Thistle): Review on its chemistry, morphology, ethno medical uses, phytochemistry and pharmacological activities. J. Drug Deliv. Ther. 2019, 9, 199–206. [Google Scholar] [CrossRef]
- El-Elimat, T.; Raja, H.A.; Graf, T.N.; Faeth, S.H.; Cech, N.B.; Oberlies, N.H. Flavonolignans from Aspergillus iizukae, a fungal endophyte of milk thistle (Silybum marianum). J. Nat. Prod. 2014, 77, 193–199. [Google Scholar] [CrossRef]
- Sampaio, B.L.; Edrada-Ebel, R.; Da Costa, F.B. Effect of the environment on the secondary metabolic profile of Tithonia diversifolia: A model for environmental metabolomics of plants. Sci. Rep. 2016, 6, 29265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimarães, D.O.; Borges, W.S.; Kawano, C.Y.; Ribeiro, P.H.; Goldman, G.H.; Nomizo, A.; Thiemann, O.H.; Oliva, G.; Lopes, N.P.; Pupo, M.T. Biological activities from extracts of endophytic fungi isolated from Viguiera arenaria and Tithonia diversifolia. FEMS Immunol. Med. Microbiol. 2008, 52, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Borges, W.d.S.; Pupo, M.T. Novel anthraquinone derivatives produced by Phoma sorghina, an endophyte found in association with the medicinal plant Tithonia diversifolia (Asteraceae). J. Braz. Chem. Soc. 2006, 17, 929–934. [Google Scholar] [CrossRef]
- Sales Junior, P.A.; Zani, C.L.; de Siqueira, E.P.; Kohlhoff, M.; Marques, F.R.; Caldeira, A.S.P.; Cota, B.B.; Maia, D.N.B.; Tunes, L.G.; Murta, S.M.F. Trypanocidal trixikingolides from Trixis vauthieri. Nat. Prod. Res. 2019, 33, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cota, B.B.; Rosa, L.H.; Caligiorne, R.B.; Rabello, A.L.T.; Almeida Alves, T.M.; Rosa, C.A.; Zani, C.L. Altenusin, a biphenyl isolated from the endophytic fungus Alternaria sp., inhibits trypanothione reductase from Trypanosoma cruzi. FEMS Microbiol. Lett. 2008, 285, 177–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petropoulos, S.A.; Fernandes, Â.; Tzortzakis, N.; Sokovic, M.; Ciric, A.; Barros, L.; Ferreira, I.C. Bioactive compounds content and antimicrobial activities of wild edible Asteraceae species of the Mediterranean flora under commercial cultivation conditions. Food Res. Int. 2019, 119, 859–868. [Google Scholar] [CrossRef] [Green Version]
- Aly, A.H.; Edrada-Ebel, R.; Wray, V.; Müller, W.E.; Kozytska, S.; Hentschel, U.; Proksch, P.; Ebel, R. Bioactive metabolites from the endophytic fungus Ampelomyces sp. isolated from the medicinal plant Urospermum picroides. Phytochemistry 2008, 69, 1716–1725. [Google Scholar] [CrossRef]
- Marangoni, S.; Moraes, T.d.S.; Utrera, S.H.; Casemiro, L.A.; de Souza, M.G.; de Oliveira, P.F.; Veneziani, R.C.; Ambrósio, S.R.; Tavares, D.C.; Martins, C.H. Diterpenes of the pimarane type isolated from Viguiera arenaria: Promising in vitro biological potential as therapeutic agents for endodontics. J. Pharm. Phytother. 2018, 10, 34–44. [Google Scholar]
- Verza, M.; Arakawa, N.S.; Lopes, N.P.; Kato, M.J.; Pupo, M.T.; Said, S.; Carvalho, I. Biotransformation of a tetrahydrofuran lignan by the endophytic fungus Phomopsis sp. J. Braz. Chem. Soc. 2009, 20, 195–200. [Google Scholar] [CrossRef]
- Fattori, V.; Zarpelon, A.C.; Staurengo-Ferrari, L.; Borghi, S.M.; Zaninelli, T.H.; Da Costa, F.B.; Alves-Filho, J.C.; Cunha, T.M.; Cunha, F.Q.; Casagrande, R. Budlein a, a sesquiterpene lactone from Viguiera robusta, alleviates pain and inflammation in a model of acute gout arthritis in mice. Front. Pharm. 2018, 9, 1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Momesso, L.d.S.; Kawano, C.Y.; Ribeiro, P.H.; Nomizo, A.; Goldman, G.H.; Pupo, M.T. Chaetoglobosinas produzidas por Chaetomium globosum, fungo endofítico associado a Viguiera robusta Gardn. (Asteraceae). Química Nova 2008, 31, 1680–1685. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.A.; Hamayun, M.; Yoon, H.; Kim, H.-Y.; Suh, S.-J.; Hwang, S.-K.; Kim, J.-M.; Lee, I.-J.; Choo, Y.-S.; Yoon, U.-H. Plant growth promotion and Penicillium citrinum. BMC Microbiol. 2008, 8, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.L.; Hussain, J.; Al-Harrasi, A.; Al-Rawahi, A.; Lee, I.-J. Endophytic fungi: Resource for gibberellins and crop abiotic stress resistance. Crit. Rev. Biotechnol. 2015, 35, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Hamayun, M.; Khan, S.A.; Iqbal, I.; Ahmad, B.; Lee, I.-J. Isolation of a gibberellin–producing fungus (Penicillium sp. MH7) and growth promotion of crown daisy (Chrysanthemum coronarium). J. Microbiol. Biotechnol. 2010, 20, 202–207. [Google Scholar] [CrossRef] [Green Version]
- Radić, N.; Štrukelj, B. Endophytic fungi—The treasure chest of antibacterial substances. Phytomedicine 2012, 19, 1270–1284. [Google Scholar] [CrossRef] [PubMed]
- Fondi, M.; Fani, R. The horizontal flow of the plasmid resistome: Clues from inter-generic similarity networks. Environ. Microbiol. 2010, 12, 3228–3242. [Google Scholar] [CrossRef]
- Dwivedi, G.R.; Sanchita; Singh, D.P.; Sharma, A.; Darokar, M.P.; Srivastava, S.K. Nano particles: Emerging warheads against bacterial superbugs. Curr. Top. Med. Chem. 2016, 16, 1963–1975. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Q.-Y.; Jia, M.; Ming, Q.-L.; Yue, W.; Rahman, K.; Qin, L.-P.; Han, T. Endophytic fungi with antitumor activities: Their occurrence and anticancer compounds. Crit. Rev. Microbiol. 2016, 42, 454–473. [Google Scholar] [CrossRef]
- Shishkoff, N.; McGrath, M. AQ10 biofungicide combined with chemical fungicides or AddQ spray adjuvant for control of cucurbit powdery mildew in detached leaf culture. Plant Dis. 2002, 86, 915–918. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Shi, Q.; Lin, G.; Guo, S.; Yang, J. Spirobisnaphthalene analogues from the endophytic fungus Preussia sp. J. Nat. Prod. 2009, 72, 1712–1715. [Google Scholar] [CrossRef]
- Du, L.; King, J.B.; Morrow, B.H.; Shen, J.K.; Miller, A.N.; Cichewicz, R.H. Diarylcyclopentendione metabolite obtained from a Preussia typharum isolate procured using an unconventional cultivation approach. J. Nat. Prod. 2012, 75, 1819–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klatt, S.; Simpson, L.; Maslov, D.A.; Konthur, Z. Leishmania tarentolae: Taxonomic classification and its application as a promising biotechnological expression host. PLoS Negl. Trop. Dis. 2019, 13, e0007424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanvir, R.; Sajid, I.; Hasnain, S. Larvicidal potential of Asteraceae family endophytic actinomycetes against Culex quinquefasciatus mosquito larvae. Nat. Prod. Res. 2014, 28, 2048–2052. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, V.; Negi, A.S.; Kumar, J.; Gupta, M.; Khanuja, S.P. Plant–based anticancer molecules: A chemical and biological profile of some important leads. Bioorg. Med. Chem. 2005, 13, 5892–5908. [Google Scholar] [CrossRef]
- Global Biodiversity Information Facility. Available online: https://www.gbif.org/ (accessed on 12 June 2020).
- MycoBank. Available online: http://www.mycobank.org/ (accessed on 12 June 2020).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Asteraceae Species and the Tissue of Endophytes Isolation | Main Therapeutic acTivities of the Host | Dominating Endophyte Genera | Main Activities of the Endophyte | Main Metabolites/Enzymes Linked to Endophyte Bioactivities |
|---|---|---|---|---|
| Achillea millefolium (stem, leaf, root) | Antioxidant, anti-inflammatory, antimicrobial, antitumor [63] | Didymella, Septoria, Stemphylium, Cladosporium, Fusarium, Alternaria, Nemania | Antitumor against lymphoblastic leukemia | L-asparaginase [56] * |
| Achillea filipendulina (stem, leaf) | Antioxidant, antidiabetic, anti-inflammatory, antimicrobial, antitumor, lubricant, antiparasitic [64,65] | Plectosphaerella, Fusarium | Antitumor against lymphoblastic leukemia | L-asparaginase [56] |
| Anthemis segetali (synonym of Cota segetalis) (stem, leaf, root, inflorescence) | Antioxidant anti-inflammatory, antitumor, antimicrobial, hepatoprotective [66] | Alternaria, Aspergillus, Bjerkandera, Schizophyllum, Fusarium, Plenodomus, Cladosporium, Didymella, Stemphylium, Nemania, Phoma, Plectosphaerella, Sarocladium | Antitumor against lymphoblastic leukemia | L-asparaginase [56] |
| Anthemis triumfettii (synonym of Cota triumfettii subsp. Triumfettii (stem, leaf) | Antioxidant anti-inflammatory, antitumor, antimicrobial, hepatoprotective [66] | Chaetosphaeronema, Stemphylium, Alternaria | Antitumor against lymphoblastic leukemia | L-asparaginase [56] |
| Artemisia annuua (stem) | Anti-inflammatory, antipyretic, antitumor, antifungal, antiparasitic, antiulcerogenic, cytotoxic [67,68,69] | Colletotrichum | Antibacterial against Bacillus subtilis, Staphylococcus aureus, Sarcina lutea, and Pseudomonas sp., and antifungal against Candida albicans and Aspergillus niger | 3β,5α-dihydroxy-6β-acetoxy-ergosta-7,22-diene; 3β,5α-dihydroxy-6β-phenylacetyloxy-ergosta-7,22-diene; 3β-hydroxy-ergosta-5-ene; 3-oxo-ergosta-4,6,8(14),22-tetraene; 3β-hydroxy-5α,8α-epidioxy-ergosta-6,22-diene [70] |
| Artemisia vulgaris | Antimalarial, anti-inflammatory, antihypertensive, antioxidant, antitumor, immunomodulatory, hepatoprotective, antispasmodic, antiseptic [71] | Chalara | Antibacterial against B. subtilis and antifungal against C. albicans | Isofusidienol A, B, C, and D [72] |
| Artemisia mongolica (stem) | Antimicrobial, insecticidal, antioxidant [73] | Colletotrichum | Antibacterial against B. subtilis, S. aureus, and S. lutea; antifungal against Bipolaris sorokiniana | Colletotric acid [44] |
| Atractylodes lancea | Anti-inflammatory, hepatoprotective [74] | Gilmaniella | Antimicrobial | Jasmonic acid [75] |
| Ayapana triplinervis | Antimicrobial, anti-inflammatory [76] | Paecilomyces, Aspergillus, Fusarium, Trichoderma, Penicillium, Curvularia | Not investigated | Not investigated [77] |
| Baccharis dracunculifolia | Immunostimulatory, anti-inflammatory, cytotoxic, antitumor, hepatoprotective [78] | Penicillium, Aspergillus, Fusarium, Colletotrichum | Not investigated | Not investigated [79] |
| Baccharis dracunculifolia (leaf) | Antioxidant, anti-inflammatory, antiviral, antimicrobial, antiparasitic [80] | Epicoccum, Pestalotiopsis, Cochliobolus, Nigrospora | Antimicrobial | Not investigated [53] |
| Bidens pilosa | Antimalarial, anti-allergic, antihypertensive, antitumor, antidiabetic, anti-inflammatory, antimicrobial, antioxidant [81] | Botryosphaeria | Antifungal, cytotoxic, antiproliferative against carcinoma cell lines | Botryorhodine A and B [82] |
| Cichorium intybus (stem, leaf, root) | Antioxidant, anti-inflammatory, cardiovascular, hypolipidemic, antitumor, antidiabetic, antimicrobial, antiparasitic [83] | Cladosporium, Epicoccum, Septoria, Plectosphaerella, Alternaria | Antitumor against lymphoblastic leukemia | L-asparaginase [56] |
| Gynura hispida | Anti-inflammatory, antiviral, hepatotoxic [84] | Bipolaris | Antifungal against Cladosporium cladosporioides, C. cucumerinum, Saccharomyces cerevisiae, Aspergillus niger, and Rhisopus oryzae | Bipolamide B [85] |
| Helianthus annuus (root) | Antibacterial, antioxidant, hepato-, nephro- and cardioprotective [86,87] | Penicillium, Aspergillus | Antifungal against Sclerotium rolfsii | Gibberellins (GA1, GA3, GA4, GA9, GA12, andGA20); organic acids (jasmonic, malic, quinic, salicylic, and succinic acid); siderophores [88,89] |
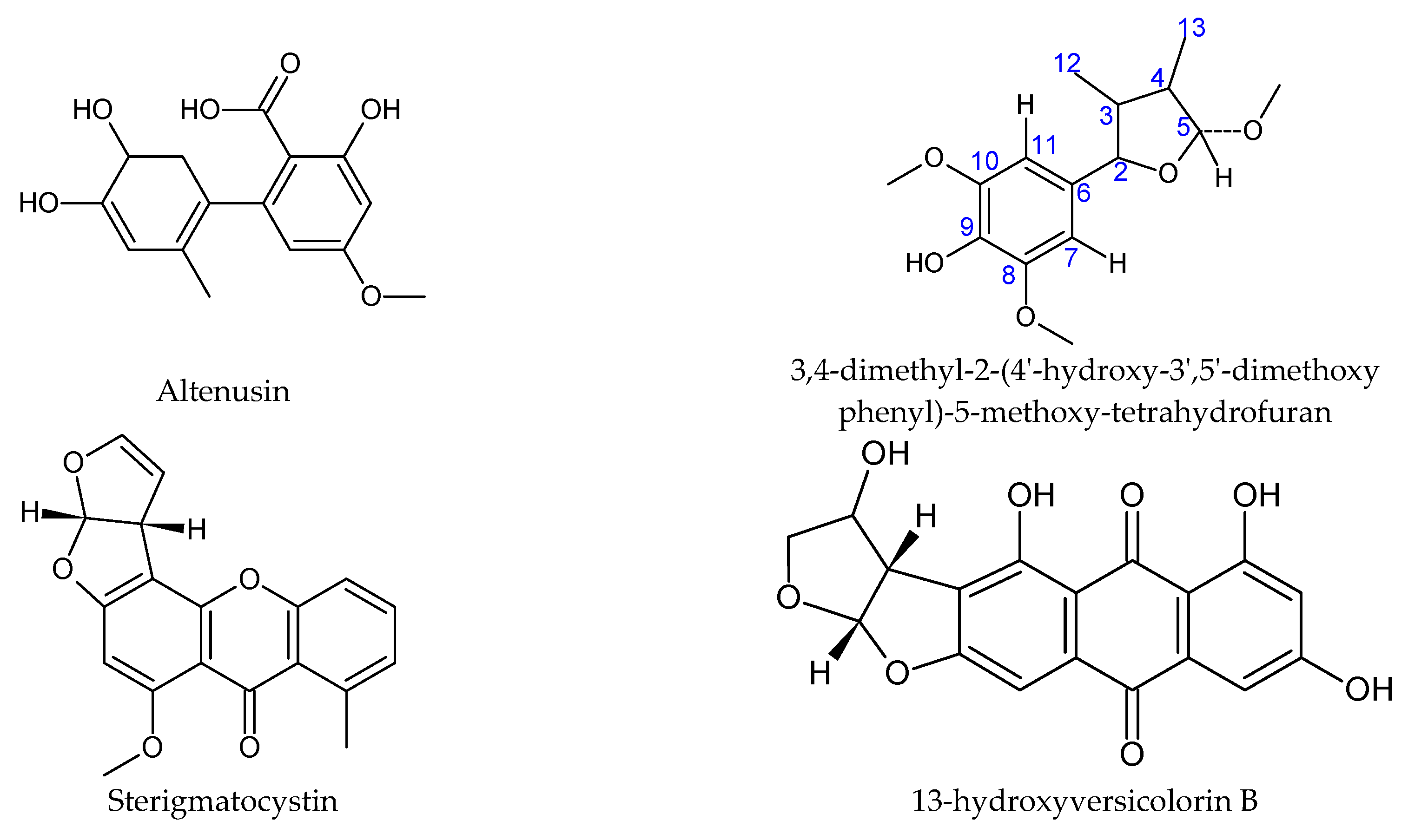
| Laggera alata | Anti-inflammatory, antioxidative, antibacterial, larvicidal [90] | Podospora | Larvicidal against Anopheles gambiae | Sterigmatocystin; 13-hydroxyversicolorin B [91] |
| Matricaria chamomilla (stem, leaf) | Anti-inflammatory, analgesic, antimicrobial, antispasmodic, sedative [92] | Epicoccum, Didymella, Phoma | Against lymphoblastic leukemia | L-asparaginase [56] |
| Mikania glomerata (leaf) | Anti-inflammatory, antispasmodic, anti-hemorrhagic, antiophidic, antiviral, antimicrobial [93] | Diaporthe | Antifungal against Fusarium solani and Didymella bryoniae; antimicrobial against Staphylococcus aureus | Not investigated [94] |
| Mikania laevigata (leaf) | Anti-inflammatory, antispasmodic, antihemorrhagic, antiophidic, antiviral, antimicrobial [93] | Hypoxylon | Not investigated | Not investigated [50] |
| Notobasis syriaca | Antioxidant, antimicrobial [95] | Phomopsis | Antimicrobial against Legionella pneumophila and Escherichia coli | Phomosine K; 2-hydroxymethyl-4β,5α,6β-trihydroxycyclohex-2-en, (-)-phyllostine; (+)-epiepoxydon; (+)-epoxydon monoacetate [96] |
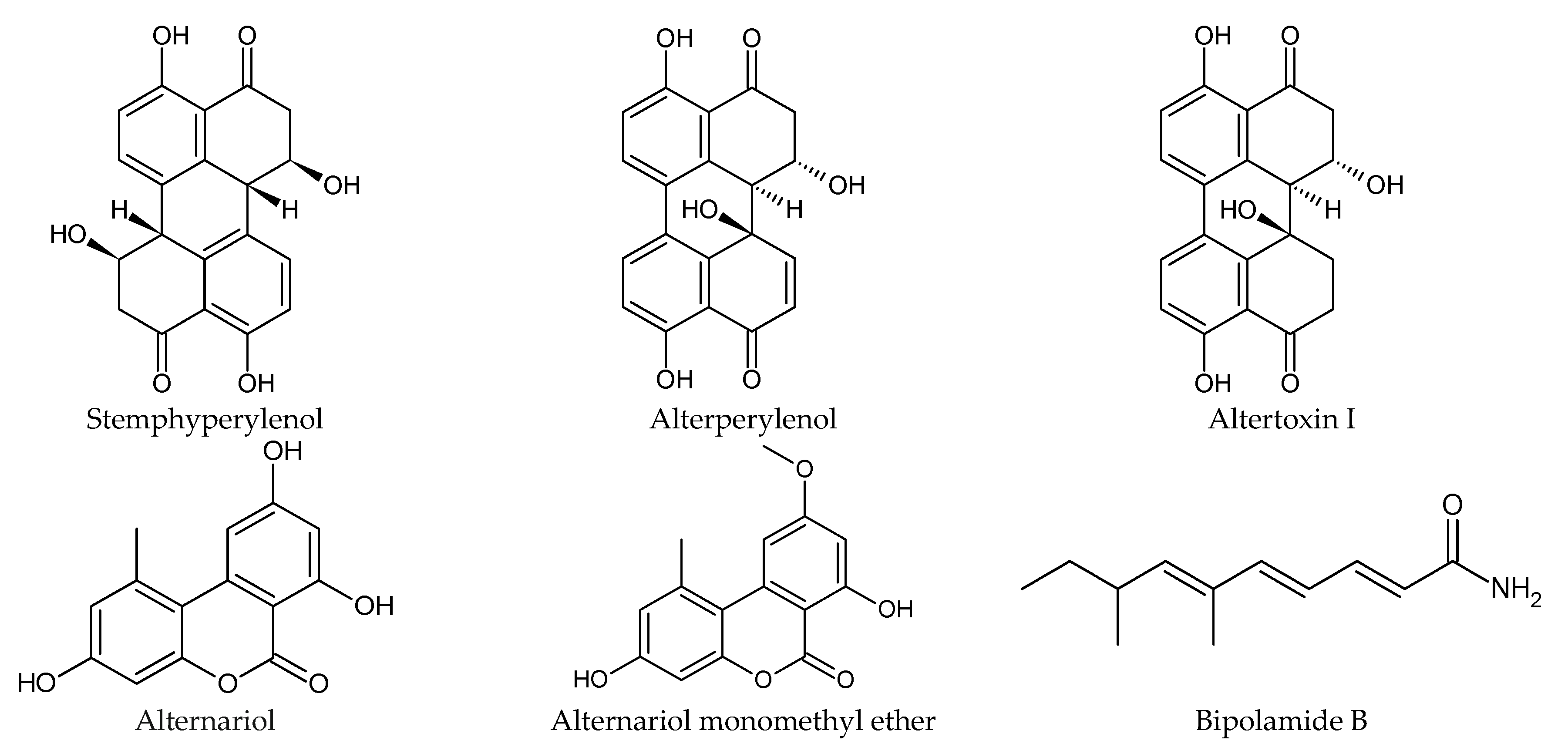
| Smallanthus sonchifolius (root, stem, leaf) | Antidiabetic, nutritious, fertility-enhancing, antioxidant, antimicrobial [97] | Curvularia | Antiparasitic against Trypanosoma cruzi | Stemphyperylenol [98] |
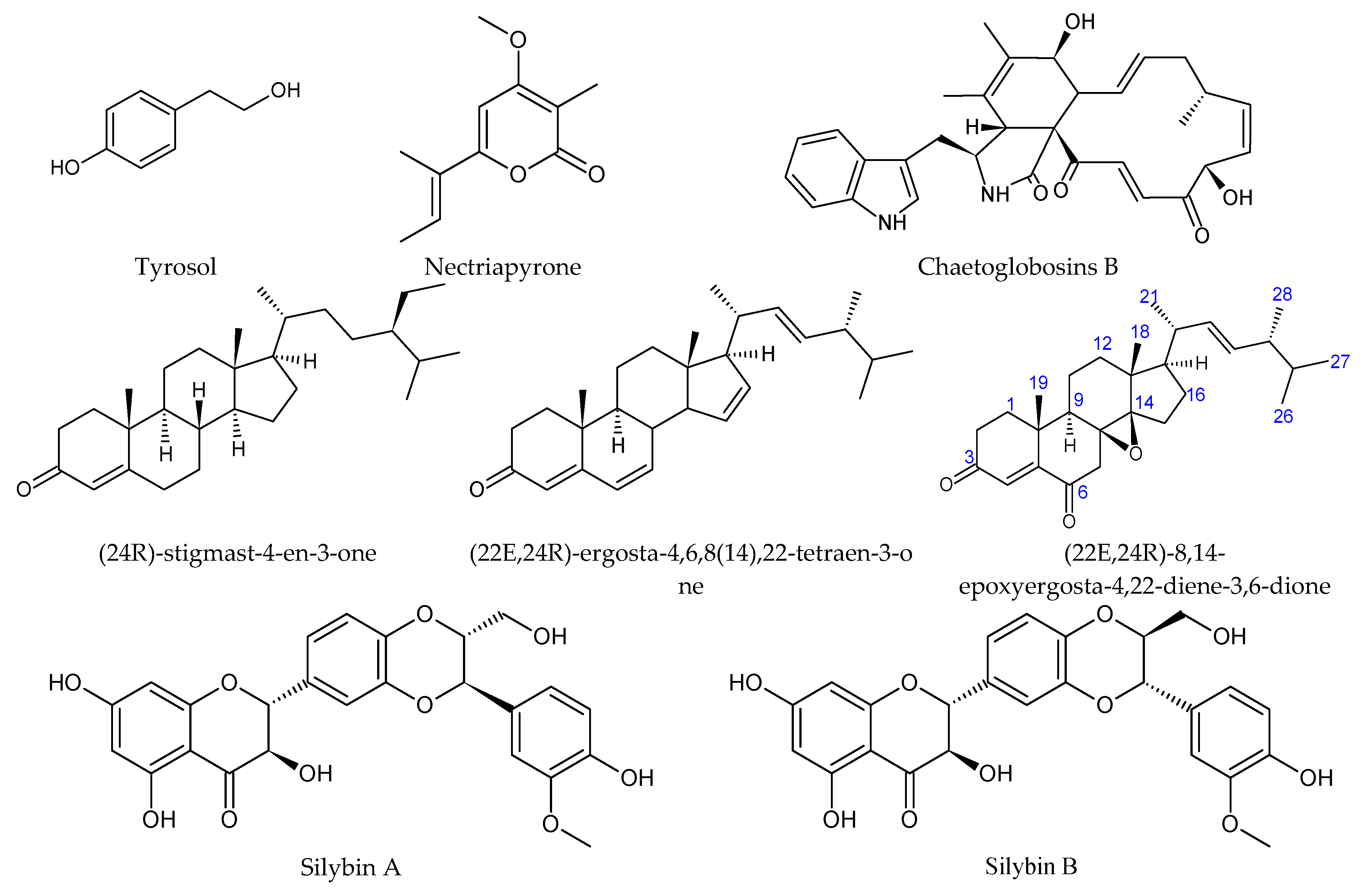
| Papulaspora | Cytotoxic against melanoma, colon, glioblastoma, and promyelocytic leukemia cell lines; antimicrobial | (24R)- stigmast-4-en-3-one; (22E,24R)- ergosta-4,6,8(14),22-tetraen-3- one; (22E,24R)-8,14- epoxyergosta-4,22-diene-3,6-dione [99] | ||
| Alternaria, Nigrospora, Phoma, Fusarium, Papulaspora | Antifungal | Stemphyperylenol, alterperylenol, altertoxin I, alternariol, alternariol monomethyl ether [100] | ||
| Coniochaeta | Antifungal against Colletotrichum acutatum, C. fragariae and C. gloeosporioides | Fatty acids: caproic, caprylic, cis-10-pentadecenoic, heptadecanoic, lauric, linoleic, myristic, oleic, palmitic, palmitoleic, pentadecanoic, stearic [101] | ||
| Silybum marianum | Antidiabetic, hepatoprotective, hypocholesterolemic, antihypertensive, anti-inflammatory, antitumor, antioxidant [102] | Aspergillus | Hepatoprotective | Silybin A, silybin B, isosilybin A [103] |
| Tithonia diversifolia | Anti-inflammatory, antimalarial, cytotoxic, gastroprotective, antimicrobial, antihyperglycemic [104] | Colletotrichum | Cytotoxic against the Jurkat tumor cell line | Nectriapyrone, tyrosol [105] |
| Phoma | Cytotoxic | Anthraquinones [106] | ||
| Trixis vauthieri (leaf) | Antiparasitic [107] | Alternaria | Trypanocidal compound with inhibitory activity of trypanothione reductase | Altenusin [108] |
| Urospermum picroides (flower) | Anti-inflammatory, immunomodulatory, antioxidant, antimicrobial [109] | Ampelomyces | Cytotoxic against L5178Y cells; antibacterial against Staphylococcus aureus, S. epidermidis and Enterococcus faecalis | 3-O-methylalaternin, altersolanol A [110] |
| Viguiera arenaria (synonym of Aldama arenaria) | Antiparasitic, analgesic, anti-inflammatory, antitumor, antimicrobial [111] | Phomopsis | Antiparasitic against T. cruzi | 3,4-dimethyl-2-(4′-hydroxy-3′,5′-dimethoxyphenyl)-5-methoxy-tetrahydrofuran [112] |
| Colletotrichum | Cytotoxic against leukemia tumor cells [106] | Nectriapyrone, tyrosol [105] | ||
| Viguiera robusta (synonym of Aldama robusta) | Anti-inflammatory, analgesic, antitumor, antiparasitic, antimicrobial [113] | Chaetomium | Cytotoxic against the Jurkat (leukemia) and B16F10 (melanoma) tumor cells; antibacterial against S. aureus and E. coli | Chaetoglobosin B [114] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caruso, G.; Abdelhamid, M.T.; Kalisz, A.; Sekara, A. Linking Endophytic Fungi to Medicinal Plants Therapeutic Activity. A Case Study on Asteraceae. Agriculture 2020, 10, 286. https://doi.org/10.3390/agriculture10070286
Caruso G, Abdelhamid MT, Kalisz A, Sekara A. Linking Endophytic Fungi to Medicinal Plants Therapeutic Activity. A Case Study on Asteraceae. Agriculture. 2020; 10(7):286. https://doi.org/10.3390/agriculture10070286
Chicago/Turabian StyleCaruso, Gianluca, Magdi T. Abdelhamid, Andrzej Kalisz, and Agnieszka Sekara. 2020. "Linking Endophytic Fungi to Medicinal Plants Therapeutic Activity. A Case Study on Asteraceae" Agriculture 10, no. 7: 286. https://doi.org/10.3390/agriculture10070286