Antibiosis Effects of Rice Carrying Bph14 and Bph15 on the Brown Planthopper, Nilaparvata lugens


Abstract
:1. Introduction
2. Materials and Methods
2.1. Rice Plants and BPHs
2.2. Evaluation of Resistance and Host Choice of Rice to BPH
2.3. Determination of Population Growth Rate, Survival Rate, and Emergence Rate of BPHs
2.4. Individual Development Duration of Nymphs on Rice Plants
2.5. Quantification of Honeydew Excretion of Female Adults
2.6. Wing Formations, Genders and Brachypterous Ratio of BPH Adults
2.7. Statistical Analysis
3. Results
3.1. BPH Resistance Level
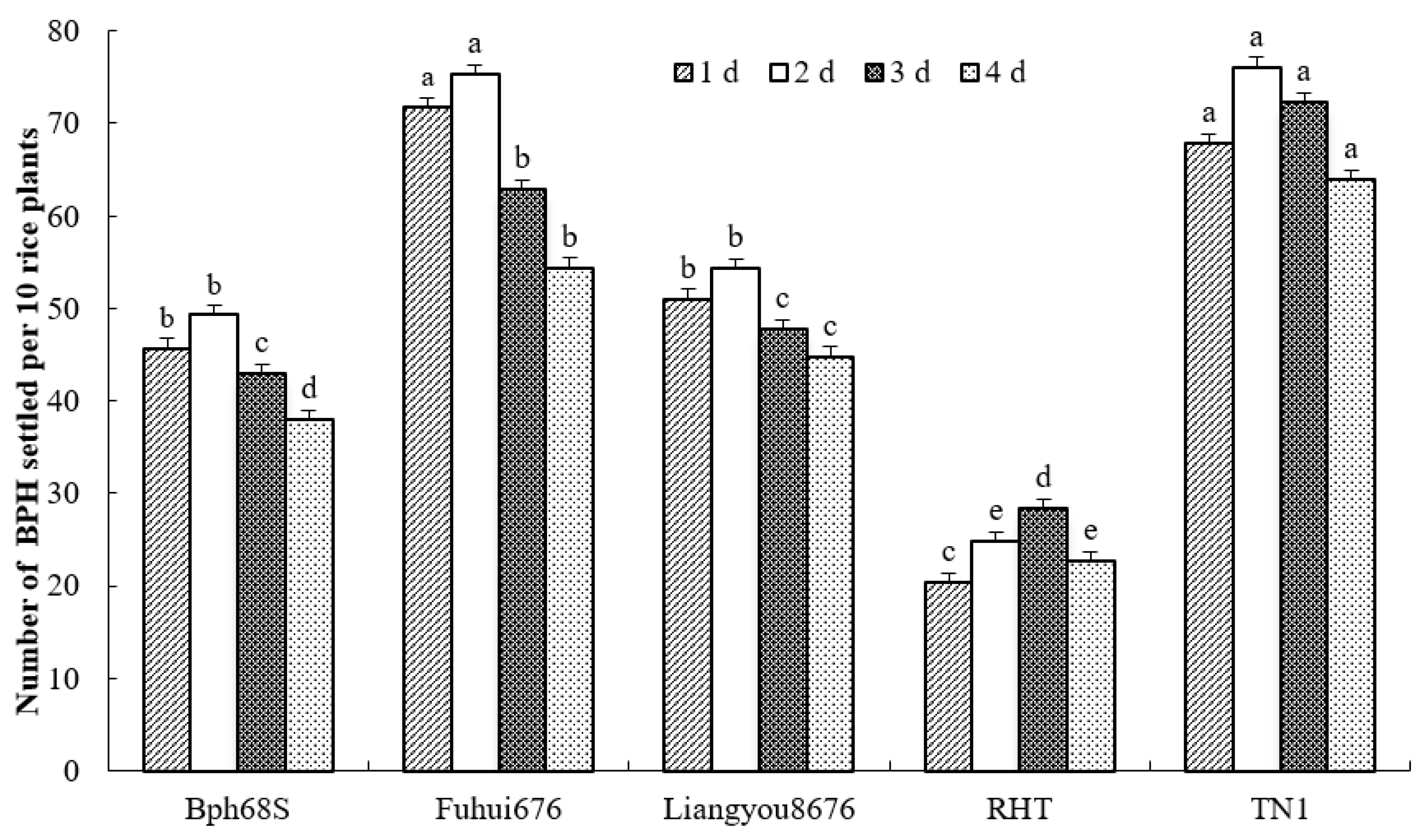
3.2. Host Feeding Choice of BPH Rice to Lines
3.3. Population Growth Rate, Survival Rate, Emergence Rate, and Developmental Duration of BPH Nymphs
3.4. Effect on Honeydew Excretion, Female Ratio, and Brachypterous Ratio of BPH Adults
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Watanabe, T.; Kitagawa, H. Photosynthesis and translocation of assimilates in rice plants following phloem feeding by the planthopper Nilaparvata lugens (Homoptera: Delphacidae). J. Econ. Èntomol. 2000, 93, 1192–1198. [Google Scholar] [CrossRef]
- Jena, K.K.; Jeung, J.U.; Lee, J.H.; Choi, H.C.; Brar, D.S. High resolution mapping of a new brown planthopper(BPH) resistance gene, Bph18(t), and marker-assisted selection for BPH resistance in rice (Oryzu saliva L.). Theor. Appl. Genet. 2006, 112, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Hu, J.; Li, Z.; Liu, J.; Gao, G.; Zhang, Q.; Xiao, J.; He, Y. Evaluation and breeding application of six brown planthopper resistance genes in rice maintainer line Jin 23B. Rice 2018, 11, 22. [Google Scholar] [CrossRef] [PubMed]
- Normile, D. Agricultural research: Reinventing Rice to Feed the World. Science 2008, 321, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Ali, P.; Chowdhury, T.R. Tagging and mapping of genes and QTLs of Nilaparvata lugens resistance in rice. Euphytica 2013, 195, 1–30. [Google Scholar] [CrossRef]
- Liu, W.C.; Liu, Z.D.; Huang, C.; Lu, M.H.; Liu, J.; Yang, Q.B. Statistics and analysis of crop yield losses caused by main diseases and insect pests in the recent 10 years. Plant Prot. 2016, 42, 1–9. (In Chinese) [Google Scholar]
- Tanaka, K.; Endo, S.; Kazano, H. Toxicity of insecticides to predators of rice planthoppers: Spiders, the mirid bug and the dryinid wasp. Appl. Èntomol. Zool. 2000, 35, 177–187. [Google Scholar] [CrossRef]
- Zhang, X.; Liao, X.; Mao, K.; Yang, P.; Li, N.; Alia, E.; Wan, H.; Li, J. The role of detoxifying enzymes in field-evolved resistance to nitenpyram in the brown planthopper Nilaparvata lugens in China. Crop. Prot. 2017, 94, 106–114. [Google Scholar] [CrossRef]
- Gorman, K.; Liu, Z.; Denholm, I.; Brüggen, K.U.; Nauen, R. Neonicotinoid resistance in rice brown planthopper, Nilaparvata lugens. Pest Manag. Sci. 2008, 64, 1122–1125. [Google Scholar] [CrossRef]
- Du, B.; Zhang, W.; Liu, B.; Hu, J.; Wei, Z.; Shi, Z.; He, R.; Zhu, L.; Chen, R.; Han, B.; et al. Identification and characterization of Bph14, a gene conferring resistance to brown planthopper in rice. Proc. Natl. Acad. Sci. USA 2009, 106, 22163–22168. [Google Scholar] [CrossRef] [Green Version]
- Naik, S.B.; Divya, D.; Sahu, N.; Sundaram, R.M.; Sarao, P.S.; Singh, K.; Lakshmi, V.J.; Bentur, J. A new gene Bph33(t) conferring resistance to brown planthopper (BPH), Nilaparvata lugens (Stål) in rice line RP2068-18-3-5. Euphytica 2018, 214, 53. [Google Scholar] [CrossRef]
- Khush, G.S. Green revolution: The way forward. Nat. Rev. Genet. 2001, 2, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Li, J.B.; Xia, M.Y.; Qi, H.X.; He, G.C.; Wan, B.L.; Zha, Z.P. Marker-Assisted Selection for Brown Planthopper (Nilaparvata lugens Stål) Resistance Genes Bph 14 and Bph 15 in Rice. Sci. Agric. Sin. 2006, 10, 2132–2137. (In Chinese) [Google Scholar]
- Deepa, K.; Pillai, M.A.; Murugesan, N. Studying the mechanisms of resistance in different rice accessions against brown planthopper (Nilaparvata lugens). J. Èntomol. Res. 2018, 42, 167. [Google Scholar] [CrossRef]
- Deng, Z.; Shi, S.J.; Wang, H.; Shangguan, X.; Liu, B.; Jing, S.L.; Du, B.; Chen, R.Z.; Zhu, L.; He, G. Analysis of QTLs for Brown Planthopper Resistance in Indica Rice WD15515. Acta Agron. Sin. 2016, 42, 353. [Google Scholar] [CrossRef]
- He, C.; Xiao, Y.; Yu, J.; Li, J.; Meng, Q.; Qing, X.; Xiao, G. Pyramiding Xa21, Bph14, and Bph15 genes into the elite restorer line Yuehui9113 increases resistance to bacterial blight and the brown planthopper in rice. Crop. Prot. 2019, 115, 31–39. [Google Scholar] [CrossRef]
- Jena, K.K.; Kim, S.M. Current Status of Brown Planthopper (BPH) Resistance and Genetics. Rice 2010, 3, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Sarao, P.S.; Sahi, G.K.; Neelam, K.; Mangat, G.S.; Patra, B.C.; Singh, K. Donors for Resistance to Brown Planthopper Nilaparvata lugens (Stål) from Wild Rice Species. Rice Sci. 2016, 23, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Laksminarayana, A.; Khush, G.S. New Genes for Resistance to the Brown Planthopper in Rice. Crop. Sci. 1977, 17, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Jairin, J.; Phengrat, K.; Teangdeerith, S.; Vanavichit, A.; Toojinda, T. Mapping of a broad-spectrum brown planthopper resistance gene, Bph3, on rice chromosome 6. Mol. Breed. 2006, 19, 35–44. [Google Scholar] [CrossRef]
- Wang, H.; Yan, Z.; Chen, J.J.; Zhou, G.X.; Fang, Y.; Wang, L.; Huang, Y.L.; Zhang, C.H.; Zhang, Y.F.; Chen, L. Research progress and prospect of brown planthopper resistance genes in rice. Hybrid Rice 2016, 4, 1–5. (In Chinese) [Google Scholar]
- Ren, X.M.; Xiang, C.; Lei, D.Y.; Guan, L.F. Present status and prospect of resistance breeding of brown planthopper in rice. Crop Res. 2017, 4, 453–458, 462. (In Chinese) [Google Scholar]
- Kang, K.; Yue, L.; Xia, X.; Liu, K.; Zhang, W.Q. Comparative metabolomics analysis of different resistant rice lines in response to the brown planthopper Nilaparvata lugens Hemiptera: Delphacidae. Metabolomics 2019, 15, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, B.Y.; Jiao, G.A.; Wang, L. Identification and application of resistance genes to brown planthopper. Acta Phytoph. Sin. 2008, 3, 279–284. (In Chinese) [Google Scholar]
- Huang, Z.; He, G.; Shu, L.; Li, X.; Zhang, Q. Identification and mapping of two brown planthopper resistance genes in rice. Theor. Appl. Genet. 2001, 102, 929–934. [Google Scholar] [CrossRef]
- Wang, B.N.; Huang, Z.; Shu, L.H.; Ren, X.; Li, X.H.; He, G.C. Molecular mapping of two new resistant genes to rice brown planthopper from wild rice. Chin. Sci. Bull. 2001, 1, 46–49. (In Chinese) [Google Scholar]
- Hu, J.; Li, X.; Wu, C.; Yang, C.; Hua, H.; Gao, G.; Xiao, J.; He, Y. Pyramiding and evaluation of the brown planthopper resistance genes Bph14 and Bph15 in hybrid rice. Mol. Breed. 2010, 29, 61–69. [Google Scholar] [CrossRef]
- Yan, Y.; Su, X.J.; Liang, M.L.; Huang, F.K.; Chen, C.H. Resistance evaluation and marker-assisted selection(MAS) for planthopper resistance genes Bph14 and Bph15 in hybrid rice. Mol. Plant Breed. 2015, 7, 1450–1456. (In Chinese) [Google Scholar]
- Hu, W.; Xiao, H.; Hu, K.; Jiang, Y.; Zhang, Y. Application of marker-assisted backcross to introgress Bph3, Bph14 and Bph15 into an eliteindicarice variety for improving its resistance to brown planthopper. Plant Breed. 2016, 135, 291–300. [Google Scholar] [CrossRef]
- Zhu, R.S.; Huang, W.C.; Hu, J.; Liu, W.J.; Zhu, Y.G. Breeding and utilization of hybrid rice Lianyou 234 and NMS line Bph68S resistance to brown planthoppper. J. Wuhan Univ. 2013, 1, 24–28. (In Chinese) [Google Scholar]
- Ainara, P.C.; Arriza, A.; Kong, L.H.; Finbarr, G.H. Aspects of brown planthopper adaptation to resistant rice lines with the Bph3 gene. Entomol. Exp. Appl. 2011, 3, 245–257. [Google Scholar]
- IRRI. Stand Evaluation System for Rice; International Rice Research Institute: Manila, Philippines, 1988. [Google Scholar]
- Qiu, Y.F.; Guo, J.P.; Jing, S.L.; Zhu, L.L.; He, G.C. High-resolution mapping of the brown planthopper resistance gene Bph6 in rice and characterizing its resistance in the 9311 and Nipponbare near isotonic backgrounds. Theor. Appl. Genet. 2010, 121, 1601–1611. [Google Scholar] [CrossRef] [PubMed]
- Klingler, J.; Creasy, R.; Gao, L.; Nair, R.M.; Calix, A.S.; Jacob, H.S.; Edwards, O.R.; Singh, K.B. Aphid Resistance in Medicago truncatula Involves Antixenosis and Phloem-Specific, Inducible Antibiosis, and Maps to a Single Locus Flanked by NBS-LRR Resistance Gene Analogs1. Plant Physiol. 2005, 137, 1445–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathak, P.K.; Saxena, R.C.; Heinrich, E.A. Para film sachet for measuring honeydew excretion by Nilaparvata lugens on rice. J. Econ. Entomol. 1982, 75, 194–195. [Google Scholar] [CrossRef]
- Li, J.; Chen, Q.; Wang, L.; Liu, J.; Shang, K.; Hua, H. Biological effects of rice carrying Bph14 and Bph15 on brown planthopper, Nilaparvata lugens. Pest Manag. Sci. 2011, 67, 528–534. [Google Scholar] [CrossRef]
- Qiu, Y.F.; Guo, J.P.; Jing, S.L.; Tang, M.; Zhu, L.L.; He, G.C. Identification of antibiosis and tolerance in rice lines carrying brown planthopper resistance genes. Entomol. Exp. Appl. 2011, 141, 224–234. [Google Scholar] [CrossRef]
- Chen, S.; Liu, F.; Xue, Y.X.; Zhu, Y.; Qin, B.X.; Wei, Z.; Qiu, Y.F.; Li, R.B. Study on evaluation and mechanism of rice resistance to brown planthopper. Southwest Chin. J. Agric. Sci. 2016, 9, 2125–2130. (In Chinese) [Google Scholar]
- Han, Y.; Wu, C.; Yang, L.; Zhang, D.; Xiao, Y. Resistance to Nilaparvata lugens in rice lines introgressed with the resistance genes Bph14 and Bph15 and related resistance types. PLoS ONE 2018, 13, e0198630. [Google Scholar] [CrossRef]
- Yuan, L.P. Development of Hybrid Rice to Ensure Food Security. Rice Sci. 2014, 21, 1–2. [Google Scholar] [CrossRef]
- Xie, F.; Peng, S. History and prospects of hybrid rice development outside of China. Chin. Sci. Bull. 2016, 61, 3858–3868. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Rice Line | Population Growth Rate (mg/BPH/d) | Survival Rate (%) | Emergence Rate (%) | Developmental Duration (d) |
|---|---|---|---|---|
| Bph68S | 0.19 ± 0.022 b | 47.92 ± 5.89 d | 43.75 ± 3.86 d | 15.1 ± 0.9 ab |
| Fuhui676 | 0.27 ± 0.018 a | 72.91 ± 5.89 b | 63.54 ± 4.31 b | 13.8 ± 0.7 c |
| Liangyou8676 | 0.21 ± 0.019 b | 58.33 ± 6.30 c | 51.04 ± 6.95 c | 14.7 ± 0.9 b |
| RHT | 0.15 ± 0.020 c | 40.63 ± 5.34 e | 34.38 ± 5.34 e | 15.6 ± 0.9 a |
| TN1 | 0.28 ± 0.021 a | 82.29 ± 5.34 a | 77.08 ± 5.89 a | 13.4 ± 1.1 c |
| Rice Line | Honeydew (mg/BPH/d) | Female Ratio (%) | Brachypterous Ratio (%) | |
|---|---|---|---|---|
| Female | Male | |||
| Bph68S | 5.43 ± 0.84 c | 43.68 ± 2.51 c | 76.97 ±4.45 c | 3.20 ±0.15 c |
| Fuhui676 | 17.49 ± 1.80 b | 54.78 ± 2.75 a | 93.78 ± 3.46 a | 6.81 ± 1.23 b |
| Liangyou8676 | 6.14 ± 0.42 c | 48.70 ± 1.21 b | 85.45 ± 1.15 b | 3.95 ± 1.67 c |
| RHT | 1.09 ± 0.44 d | 37.60 ± 2.36 d | 63.65 ± 2.81 d | 0.00 ± 0.00 d |
| TN1 | 22.44 ± 2.84 a | 56.74 ± 2.81 a | 95.39 ± 0.86 a | 10.64 ± 1.46 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, L.; Wang, W.; Shi, L.; Liu, Q.; Zhan, Z. Antibiosis Effects of Rice Carrying Bph14 and Bph15 on the Brown Planthopper, Nilaparvata lugens. Agriculture 2020, 10, 109. https://doi.org/10.3390/agriculture10040109
Qiu L, Wang W, Shi L, Liu Q, Zhan Z. Antibiosis Effects of Rice Carrying Bph14 and Bph15 on the Brown Planthopper, Nilaparvata lugens. Agriculture. 2020; 10(4):109. https://doi.org/10.3390/agriculture10040109
Chicago/Turabian StyleQiu, Liangmiao, Wuqi Wang, Longqing Shi, Qiquan Liu, and Zhixiong Zhan. 2020. "Antibiosis Effects of Rice Carrying Bph14 and Bph15 on the Brown Planthopper, Nilaparvata lugens" Agriculture 10, no. 4: 109. https://doi.org/10.3390/agriculture10040109