The Evaluation of Growth Performance, Photosynthetic Capacity, and Primary and Secondary Metabolite Content of Leaf Lettuce Grown under Limited Irradiation of Blue and Red LED Light in an Urban Plant Factory

 ,
, 
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Leaf Photosynthesis and Chlorophyll Fluorescence
2.3. Growth Analysis
2.4. Extraction for Polyphenol Analysis
2.5. Estimation of Flavonoid, Anthocyanin, and Chlorogenic Acid by HPLC
2.6. Carbohydrate and Pigments Analysis
2.7. Statistical Analysis
3. Results and Discussion
3.1. Lettuce Plants Adapt Their Photosynthetic Apparatus to the Light Conditions
3.2. Lettuce Morphology Characteristics Are Affected by the B/R Light Ratio
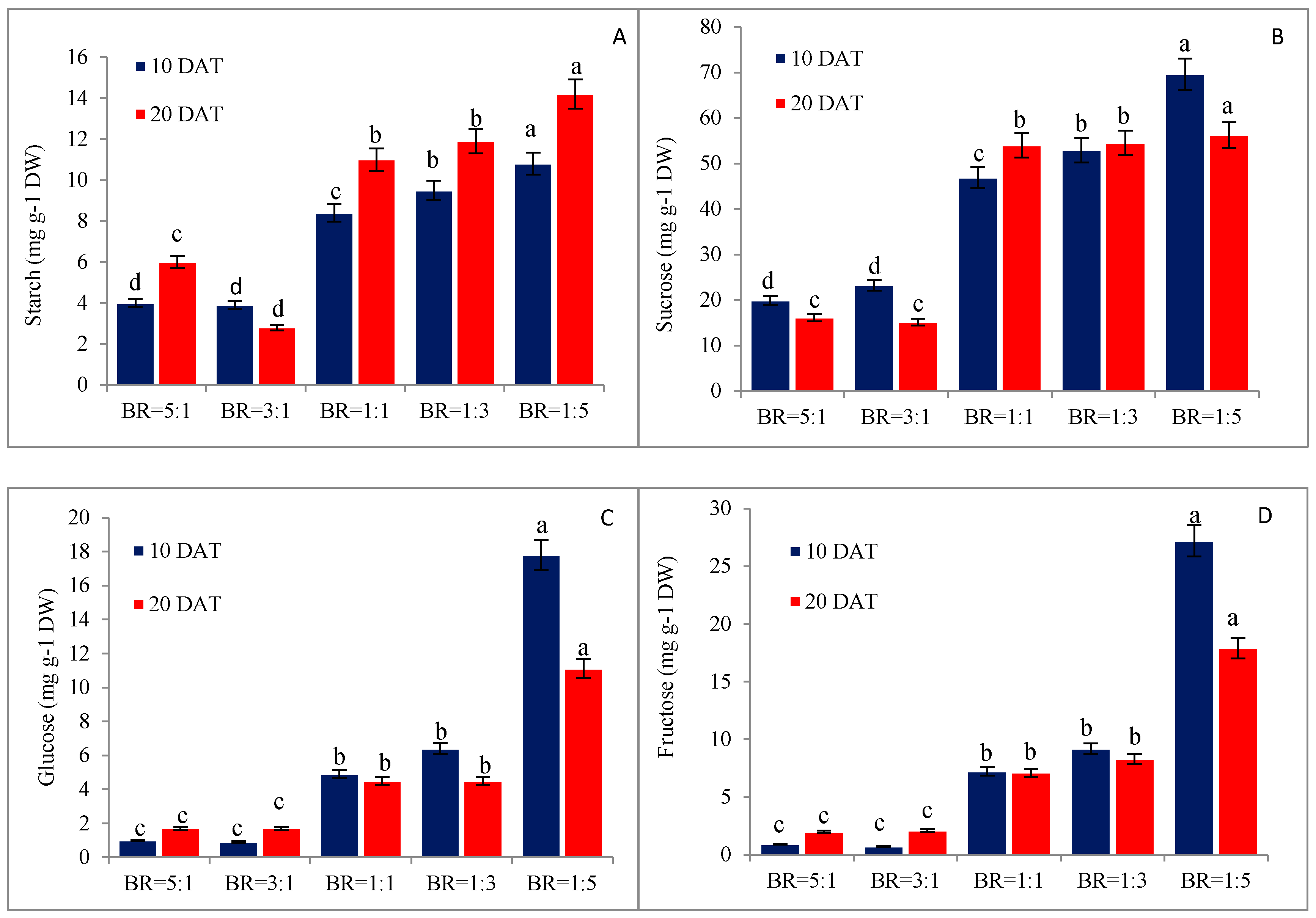
3.3. Red Light Increase Carbohydrate Contents
3.4. Blue Light Increases the Leaf Content of Pigments
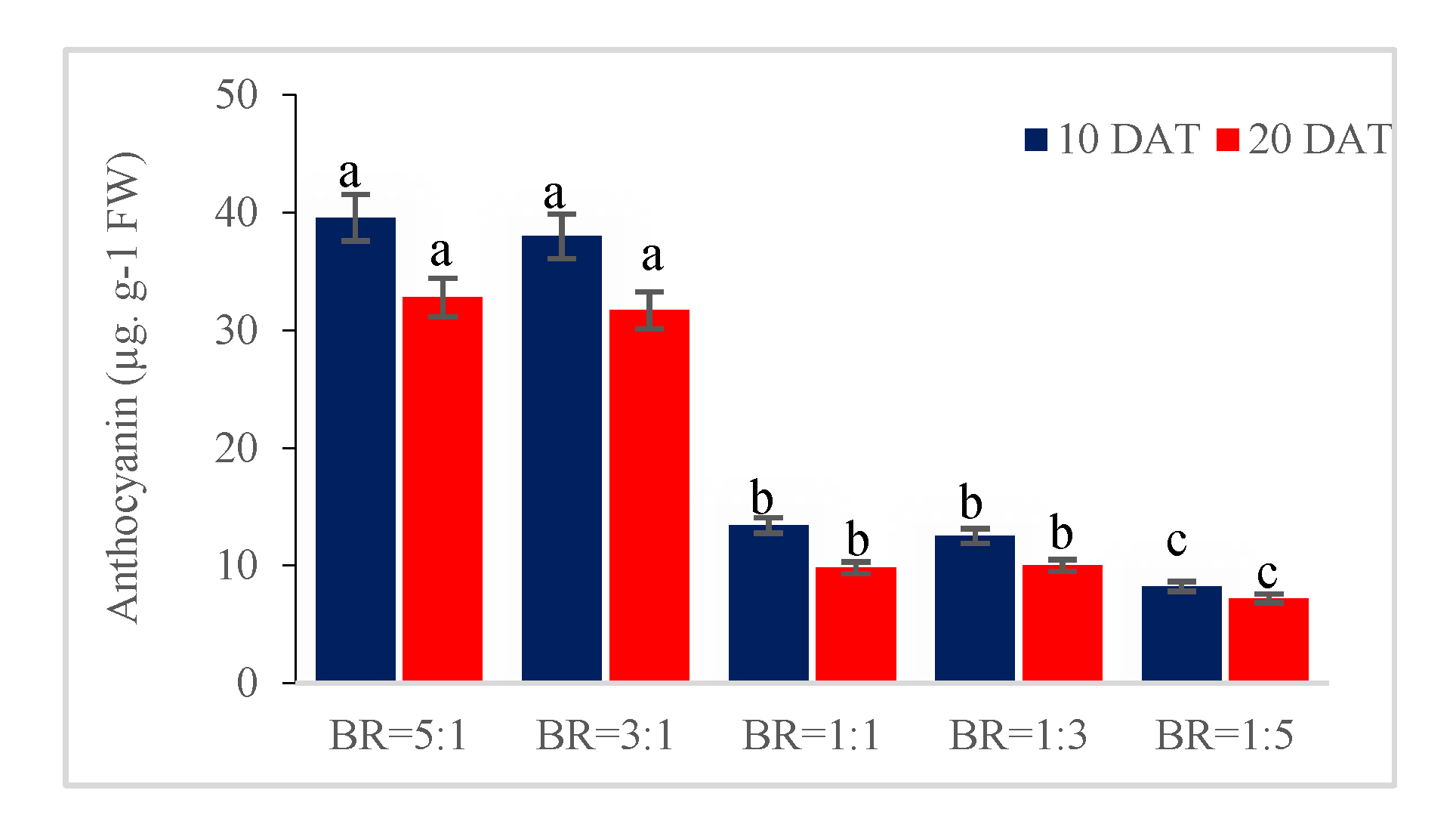
3.5. Blue Light Increases the Content of Phenolic Compounds
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kozai, T. Smart Plant Factory—The Next Generation Indoor Vertical Farms; Springer: Berlin, Germany, 2018. [Google Scholar]
- Mou, B. Nutritional quality of lettuce. Curr. Nutr. Food Sci. 2012, 8, 177–187. [Google Scholar] [CrossRef]
- Konstantopoulou, E.; Kapotis, G.; Salachas, G.; Petropoulos, S.A.; Karapanos, I.C.; Passam, H.C. Nutritional quality of greenhouse lettuce at harvest and after storage in relation to N application and cultivation season. Sci. Hortic. 2010, 125, 93–94. [Google Scholar] [CrossRef]
- Park, Y.G.; Park, J.E.; Hwang, S.J.; Jeong, B.R. Light source and CO2 concentration affect growth and anthocyanin content of lettuce under controlled environment. Hortic. Environ. Biotechnol. 2012, 53, 460–466. [Google Scholar] [CrossRef]
- Naznin, M.T.; Lefsrud, M.; Gravel, V.; Azad, M.O.K. Blue light added with red LEDs enhance growth characteristics, pigments content, and antioxidant capacity in lettuce, spinach, kale, basil, and sweet pepper in a controlled environment. Plants 2019, 8, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naznin, M.T.; Lefsrud, M.; Azad, M.O.K.; Park, C.H. Effect of different combinations of red and blue LED light on growth characteristics and pigment content of in vitro tomato plantlets. Agriculture 2019, 9, 196. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.L.; Yang, Q.C.; Song, W.P.; Wang, L.C.; Go, W.Z. Growth and nutritional properties of lettuce affected by different alternating intervals of red and blue LED irradiation. Sci. Hortic. 2017, 223, 4–52. [Google Scholar] [CrossRef]
- Terashima, I.; Fujita, T.; Inoue, T.; Chow, W.S.; Oguchi, R. Green light drives leaf photosynthesis more efficiently than red light in strong white light: Revisiting the enigmatic question of why leaves are green. J. Plant Cell Physiol. 2009, 50, 684–697. [Google Scholar] [CrossRef] [Green Version]
- Hogewoning, S.W.; Trouwborst, G.; Maljiaars, H.; Poorter, H.; Van leperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Abidi, F.; Girault, T.; Douillet, O.; Guillemain, G.; Sintes, G.; Laffaire, M.; Leduc, N. Blue light effects on rose photosynthesis and photomorphogenesis. Plant Biol. 2013, 15, 67–74. [Google Scholar] [CrossRef]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C.O. Spectral effects of artificial light on plant physiology and secondary metabolism: A review. Hortscience 2015, 50, 1128–1135. [Google Scholar] [CrossRef] [Green Version]
- Perez-Balibrea, S.; Moreno, D.A.; Garcia- Viguera, C. Influence of light on health-promoting phytochemicals of broccoli sprouts. J. Sci. Food Agric. 2008, 88, 904–910. [Google Scholar] [CrossRef]
- Kim, S.J.; Hahn, E.J.; Heo, J.W.; Paek, K.Y. Effects of LEDs on net photosynthetic rate, growth and leaf stomata of chrysanthemum plantlets in vitro. Sci. Hortic. 2004, 101, 143–151. [Google Scholar] [CrossRef]
- Wu, M.C.; Hou, C.Y.; Jiang, C.M.; Wang, C.Y.; Chen, H.H.; Chang, H.M. A novel approach of LED light radiation improves the antioxidant activity of pea seedlings. Food Chem. 2007, 101, 1753–1758. [Google Scholar] [CrossRef]
- Son, K.H.; Oh, M.M. Leaf shape, growth and antioxidant, phenolic compounds of two lettuce cultivars grown under various combinations of blue and red light-emitting diodes. HortScience 2013, 48, 988–995. [Google Scholar] [CrossRef]
- Shin, K.S.; Mrthy, H.N.; Heo, J.W.; Hahn, E.J.; Paek, K.Y. The effect of light quality on the growth and development of in vitro cultured doritaenopsis plants. Acta Physiol. Plant. 2008, 30, 339–343. [Google Scholar] [CrossRef]
- Lee, J.G.; Oh, S.S.; Cha, S.H.; Jang, Y.A.; Kim, S.Y.; Um, Y.C.; Cheong, S.R. Effects of red/blue light ratio and short-term light quality conversion on growth and anthocyanin contents of baby leaf lettuce. Prot. Hortic. Plant Fact. 2010, 19, 351–359. [Google Scholar]
- Becker, C.; Klaering, H.P.; Schreiner, M.; Kroh, L.W.; Krumbein, A. Unlike quercetin glycosides, cyanidin glycoside in red leaf lettuce responds more sensitively to increasing low radiation intensity before than after head formation has started. J. Agric. Food Chem. 2014, 62, 6911–6917. [Google Scholar] [CrossRef]
- Urrestarazu, M.; Najera, C.; del Mar Gea, M. Effect of the spectral quality and intensity of light-emitting diodes on several horticultural crops. HortScience 2016, 51, 268–271. [Google Scholar] [CrossRef] [Green Version]
- Zhen, S.; van Iersel, M.W. Photochemical acclimation of three contrasting species to different light levels: Implications for optimizing supplemental lighting. J. Am. Soc. Hort. Sci. 2017, 142, 346–354. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Petropoulos, S.A.; El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Giordano, M.; Troise, A.D.; Vitaglione, P.; De Pascale, S. Reducing energy requirements in future bioregenerative life support systems (blsss): Performance and bioactive composition of diverse lettuce genotypes grown under optimal and suboptimal light conditions. Front. Plant Sci. 2019, 10, 1305. [Google Scholar] [CrossRef]
- Huang, L.C.; Chen, Y.H.; Chen, Y.H.; Wang, C.F.; Hu, M.C. Food-energy interactive tradeoff analysis of sustainable urban plant factory production systems. Sustainability 2018, 10, 446. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, Y.; Kanematsu, Y.; Yoshikawa, N.; Okubo, T.; Takagaki, M. Environmental and resource use analysis of plant factories with energy technology options: A case study in Japan. J. Clean. Prod. 2018, 186, 703–717. [Google Scholar] [CrossRef]
- Yeh, N.; Chung, J.P. High-brightness LEDs—Energy efficient lighting sources and their potential in indoor plant cultivation. Renew. Sustain. Energy 2009, 13, 2175–2180. [Google Scholar] [CrossRef]
- Perez, V.M. Study of the Sustainability Issues of Food Production Using Vertical Farm Methods in an Urban Environment Within the State of Indiana. Master’s Thesis, Purdue University, West Lafayette, IN, USA, 2014. [Google Scholar]
- Loconsole, D.; Cocetta, G.; Santoro, P.; Ferrante, A. Optimization of led lighting and quality evaluation of romaine lettuce grown in an innovative indoor cultivation system. Sustainability 2019, 11, 841. [Google Scholar] [CrossRef] [Green Version]
- Han, T.; Vaganov, V.; Cao, S.; Li, Q.; Ling, L.; Cheng, X.; Peng, L.; Zhang, C.; Yakovlev, A.N.; Zhong, Y.; et al. Improving “color rendering” of LED lighting for the growth of lettuce. Sci. Rep. 2017, 7, 45944. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Ferreres, F.; Gil, M.I.; Castaner, M.; Tomas-Barberan, F.A. Phenolic metabolites in red pigmented lettuce (Lactuca sativa L.) changes with minimal processing and cold Storage. J. Agric. Food Chem. 1997, 45, 4249–4254. [Google Scholar] [CrossRef]
- Kjaer, K.H.; Ottosen, C.O. Growth of chrysanthemum in response to supplemental light provided by irregular light breaks during the night. J. Am. Soc. Hort. Sci. 2011, 136, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Van Iersel, M.W.; Weaver, G.; Martin, M.T.; Ferrarezi, R.S.; Mattos, E.; Haidekker, M. A chlorophyll fluorescence-based biofeedback system to control photosynthetic lighting in controlled environment agriculture. J. Am. Soc. Hort. Sci. 2016, 141, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Van Iersel, M.W.; Gianino, D. An adaptive control approach for LED lights can reduce the energy costs of supplemental lighting in greenhouses. HortScience 2017, 52, 72–77. [Google Scholar] [CrossRef]
- Matsuda, R.; Ohashi-kaneko, K.; Fujiwara, K.; Goto, E.; Kurata, K. Photosyntehtic characteristics of rice leaves grown under red light with or without supplemental blue light. Plant Cell Physiol. 2004, 45, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Johkan, M.; Shoji, K.; Goto, F.; Hahida, S.; Yoshihara, T. Blue light emitting diode light irradiation of seedlings improves seedling quality and growth after transplanting in red leaf lettuce. HortScience 2010, 45, 1809–1814. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 2016, 7, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C. Phototropin blue light receptors and light—induced movement response in plants. Sci. Stke 2002, 118, pe5. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, J.I.; Allen, G.J.; Hugouvieux, V.; Kwak, J.M.; Waner, D. Guard cell signal transduction. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 627–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimazaki, K.I.; Doi, M.; Assmann, S.M.; Kinoshita, T. Light regulation of stomatal movement. Annu. Rev. Plant Biol. 2007, 58, 219–247. [Google Scholar] [CrossRef] [Green Version]
- Rosenqvist, E. Light acclimation maintains the redox state of the PS II electron accepter QA within a narrow range over a broad range of light intensities. Photosynth. Res. 2002, 70, 299–310. [Google Scholar] [CrossRef]
- Heo, J.W.; Kang, D.H.; Bang, H.S.; Hong, S.G.; Chun, C.; Kang, K.K. Early growth, pigmentation, protein content, and phenylalanine ammonia-lyase activity of red colored lettuces grown under different lighting condition. Korean J. Hort. Sci. Technol. 2012, 30, 6–12. [Google Scholar]
- Quail, P.H. Photosensory perception and signaling in plant cells: New paradigms. Curr. Opin. Cell Biol. 2002, 14, 180–188. [Google Scholar] [CrossRef]
- Wang, H.; Ma, L.G.; Li, J.M.; Zhao, H.Y.; Deng, X.W. Direct interaction of Arabidopsis cryptochromes with COP1 in light control development. Science 2001, 294, 154–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Garcia, J.F.; Huq, E.; Quail, P.H. Direct targeting of light signals to a promoter element-bound transcription factor. Science 2000, 288, 859–863. [Google Scholar] [PubMed] [Green Version]
- Hoenecke, M.E.; Bula, R.J.; Tibbitts, T.W. Importance of blue photon levels for lettuce seedlings grown under red light emitting diodes. HortScience 1992, 27, 427–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dougher, T.A.O.; Bugbee, B. Long term blue light effects on the histology of lettuce and soybean leaves and stem. J. Am. Soc. Hort. Sci. 2004, 129, 467–472. [Google Scholar] [CrossRef] [Green Version]
- Samuoliene, G.; Sirtautas, R.; Brazaityte, A.; Sakalauskaite, J.; Sakalauskiene, S.; Duchovskis, P. The impact of red and blue light-emitting diode illumination on radish physiological indices. Cent. Eur. J. Biol. 2011, 6, 821–828. [Google Scholar] [CrossRef]
- Hernandez, R.; Kubota, C. Physiological responses of cucumber seedlings under different blue and red photon flux ratios using LEDs. Environ. Exp. Bot. 2016, 121, 66–74. [Google Scholar] [CrossRef]
- Kang, W.H.; Park, J.S.; Park, K.S.; Son, J.E. Leaf photosynthetic rate, growth, and morphology of lettuce under different fractions of red, blue, and green light from light-emitting diodes (LEDs). Hortic. Environ. Biotechnol. 2016, 57, 573. [Google Scholar] [CrossRef]
- Chatterjee, M.; Sharma, P.; Khurana, J.P. Cryptochrome 1 from Brassica napus is up-regulated by blue light and controls hypocotyl/stem growth and anthocyanin accumulation. Plant Physiol. 2006, 141, 61–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metallo, R.M.; Kopsell, D.A.; Sams, C.E.; Bumgarner, N.R. Influence of blue/red vs. white LED light treatments on biomass, shoot morphology, and quality parameters of hydroponically grown kale. Sci. Hortic. 2018, 235, 189–197. [Google Scholar] [CrossRef]
- Ouzounis, T.; Frette, X.; Rosenqvist, E.; Ottosen, C.O. Spectral effects of supplementary lighting on the secondary metabolites in roses, chrysanthemums, and campanulas. J. Plant Physiol. 2014, 171, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Kang, J.H.; Jeong, B.R.; Hwang, S.J. Light quality and photoperiod affect growth of sowthistle (Ixeris dentata Nakai) in a closed-type plant production system. Korean J. Hort. Sci. Technol. 2016, 34, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Lin, K.H.; Huang, M.Y.; Huang, W.D.; Hsu, M.H.; Yang, Z.W.; Yang, C.M. The effect of red, blue and white light emitting diodes on the growth, development and edible quality of hydroponically grown lettuce (Lactusa sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Li, H.; Tang, C.; Xu, Z.; Liu, X.; Han, X. Effect of different light sources on the growth of non-heading Chinese cabbage (Brassica campestris L.). J. Agr. Sci. 2012, 4, 262–273. [Google Scholar] [CrossRef] [Green Version]
- Saebo, A.; Krekling, T.; Appelgren, M. Light quality affects photosynthesis and leaf anatomy of brich plantlets in vitro. Plant Cell Tiss. Organ Cult. 1995, 41, 177–185. [Google Scholar] [CrossRef]
- Li, H.; Tang, C.; Xu, Z. The effect of different light qualities on rape seed (Bassica napus L.) plantlet growth and morphogenesis in vitro. Sci. Hortic. 2013, 150, 117–124. [Google Scholar] [CrossRef]
- Poffenroth, M.; Green, D.B.; Tallman, G. Sugar concentration in guard cells of Vicia faba illuminated with red or blue light. Plant Physiol. 1992, 98, 1460–1471. [Google Scholar] [CrossRef] [Green Version]
- Yagi, K.; Hamada, K.; Hirata, K.; Miyamoto, K.; Miura, Y.; Akano, T.; Fukatu, K.; Ikuta, Y.; Nakamura, H.K. Stimulatory effect of red light on starch accumulation in a marine green alga, Chlamydomonas sp. strain MGA161. Appl. Biochem. Biotech. 1994, 45, 225–232. [Google Scholar] [CrossRef]
- Cheng, J.; Fan, P.; Liang, Z.; Wang, Y.; Nie, N.; Li, W.; Li, S. Accumulation of end products in source leaves affects photosynthetic rate in peach via alteration of stomatal conductance and photosynthetic efficiency. J. Am. Soc. Hort. Sci. 2009, 134, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Zang, L.; Kato, M.; Yamawaki, K.; Kiriiwa, Y.; Yahata, M.; Ikoma, Y.; Matsumoto, H. Effect of blue light and red LED light irradiation on β-cryptoxanthin accumulation in the flavedo of citrus fruits. J. Agr. Food Chem. 2012, 60, 197–201. [Google Scholar] [CrossRef]
- Bukhov, N.G.; Drozdova, I.S.; Bondar, V.V.; Mokronosov, A.T. Blue, red and blueplus red light control of chlorophyll content and CO2gas exchange in barleyleaves: Quantitative description of the effects of light quality and fluence rate. Physiol. Plant. 1992, 85, 632–638. [Google Scholar] [CrossRef]
- Evans, J.R. Acclimation by the thylakoid membranes to growth irradiance and the partitioning of nitrogen between soluble and thylakoid proteins. Aust. J. Plant Physiol. 1988, 15, 93–106. [Google Scholar] [CrossRef]
- Yu, X.; Liu, H.; Klejnot, J.; Lin, C. The Cryptochrome Blue Light Receptors; The Arabidopsis Book; ASPB: Rockville, MD, USA, 2010. [Google Scholar]
- Stutte, G.W.; Edney, S.L.; Newsham, G.J. Photoregulation of Anthoycyanin Production in Red Leaf Lettuce with Blue LEDs is Affected by Timing and Leaf Age. In Proceedings of the 36th Annual Meeting of the Plant Growth Regulation Society of America, North Carolina, CA, USA, 2–6 August 2009; pp. 125–130. [Google Scholar]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effect of light quality on the accumulation of phytochemcials in vegetables produced in controlled environments: A review. J. Sci. Food Agric. 2013, 95, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Chung, M.G.; Kim, J.B.; Hahn, B.S.; Kim, J.B.; Bae, S.C.; Roh, K.H.; Kim, Y.H.; Cheon, C.I.; Sung, M.K.; et al. Genes up—regulated during red coloration in UVB irradiated lettuce leaves. Plant Cell Rep. 2007, 26, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Awad, M.A.; Patricia, S.; Wagenmakers, P.S.; de Jager, A. Effects of light on flavonoid and chlorogenic acid levels in the skin of Jona gold apples. Sci. Hortic. 2001, 88, 289–298. [Google Scholar] [CrossRef]
- Giliberto, L.; Perrotta, G.; Pallara, P.; Weller, J.L.; Fraser, P.D.; Bramley, P.M.; Fiore, A.; Tavazza, M.; Giuliano, G. Manipulation of the blue light photoreceptor cryptochrome-2 in tomato affects vegetative development, flowering time and fruit antioxidant content. Plant Physiol. 2005, 137, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narukawa, M.; Watanable, K.; Inoue, Y. Light-induced root hair formation in lettuce (Lactusasativa L.cv. Grand Rapids) roots at low PH is brought by chlorogenic acid synthesis and sugar. J. Plant Res. 2010, 123, 789–799. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height (cm) | Leaf Number Plant−1 | LA Plant−1 (cm2) | SLA Plant−1 (cm2 g−1) | FW Plant−1 (g) | DW Plant−1 (g) |
|---|---|---|---|---|---|---|
| B/R = 5:1 | 7.10 ± 0.96d y | 6± 0.69a | 87 ± 25.5d | 473 ± 102b | 3.0 ± 1.00b | 0.18 ± 0.06ab |
| B/R = 3:1 | 8.3 ± 0.94c | 6± 0.48a | 119 ±21.4 bc | 469 ± 85b | 4.0 ± 0.89a | 0.26 ±0.07a |
| B/R = 1:1 | 9.7 ± 0.63b | 6± 0.42a | 111 ± 12.6c | 602 ± 108a | 4.2 ± 0.70b | 0.19 ± 0.05ab |
| B/R = 1:3 | 10.0 ± 0.55b | 6± 0.56a | 137± 14.2ab | 546 ± 69ab | 5.0 ± 0.74a | 0.25 ±0.04a |
| B/R = 1:5 | 11.0 ± 0.62a | 6 ± 0.31a | 149 ± 15.9a | 632 ± 75a | 4.6 ± 0.74a | 0.24 ±0.03a |
| Treatments | Plant Height (cm) | Leaf Number Plant−1 | LA Plant−1 (cm2) | SLA Plant−1 (cm2g−1) | FW Plant−1 (g) | DW Plant−1 (g) |
|---|---|---|---|---|---|---|
| B/R = 5:1 | 9.7 ± 0.58d y | 10 ± 1.0a | 234 ± 35.1c | 539 ± 58b | 9.4 ± 1.6a | 0.44 ± 0.08a |
| B/R = 3:1 | 10.1 ± 0.41d | 10± 0.66a | 255 ± 34.0bc | 580 ± 181b | 10.6 ± 1.4a | 0.46 ± 0.10a |
| B/R = 1:1 | 11.0 ± 0.82c | 9 ± 0.82ab | 243 ± 20.4bc | 485 ± 218b | 10.3 ± 1.1a | 0.55 ± 0.15a |
| B/R = 1:3 | 11.9 ± 0.56b | 10 ± 0.69ab | 271 ± 21.9ab | 1044 ± 175a | 10.7 ± 0.8a | 0.26 ± 0.04b |
| B/R = 1:5 | 12.7 ± 0.92a | 9 ± 0.63ab | 295 ± 32.9a | 1275 ± 399a | 10.9 ± 1.6a | 0.25 ± 0.09b |
| Treatments | Chl a mg g−1 DW | Chl b mg g−1 DW | Total Chl mg g−1 DW | Chl a/b Ratio | Carotenoid mg g−1 DW | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 10 DAT | 20 DAT | 10 DAT | 20 DAT | 10 DAT | 20 DAT | 10 DAT | 20 DAT | 10 DAT | 20 DAT | |
| B/R = 5:1 | 9.2 ± 2.0a y | 9.6 ± 2.5a | 2.9 ± 0.7a | 4.1 ± 0.5a | 12.0 ± 2.7a | 13.7 ± 2.9a | 3.2 ± 0.2a | 2.3 ± 0.5b | 3.1 ± 0.4a | 3.8 ± 0.8a |
| B/R = 3:1 | 6.3 ± 1.3b | 6.2 ± 2.1b | 2.4 ± 0.5a | 2.4 ± 0.9b | 8.7 ± 1.4b | 8.6 ± 3.0b | 2.8 ± 0.9a | 2.6 ± 0.3ab | 2.5 ± 0.6b | 2.0 ± 0.7b |
| B/R = 1:1 | 4.7 ± 0.7bc | 5.8 ± 1.5b | 2.4 ± 0.5a | 2.1 ± 0.8b | 7.2 ± 1.1b | 7.9 ± 2.2b | 1.9 ± 0.2b | 2.8 ± 0.6ab | 1.8 ± 0.3c | 2.0 ± 0.7b |
| B/R = 1:3 | 4.6 ± 0.9c | 6.0 ± 0.9b | 2.5 ± 0.5a | 2.0 ± 0.5b | 7.1 ± 1.3b | 8.1 ± 1.4b | 1.8 ± 0.2b | 3.0 ± 0.4a | 1.9 ± 0.4c | 1.5 ± 0.2b |
| B/R = 1:5 | 4.5 ± 0.4c | 4.8 ± 1.5b | 2.3 ± 0.4a | 1.8 ± 0.9b | 6.8 ± 0.7b | 6.6 ± 2.4b | 1.9 ± 0.b | 2.8 ± 0.6ab | 1.7 ± 0.2c | 1.6 ± 0.9b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azad, M.O.K.; Kjaer, K.H.; Adnan, M.; Naznin, M.T.; Lim, J.D.; Sung, I.J.; Park, C.H.; Lim, Y.S. The Evaluation of Growth Performance, Photosynthetic Capacity, and Primary and Secondary Metabolite Content of Leaf Lettuce Grown under Limited Irradiation of Blue and Red LED Light in an Urban Plant Factory. Agriculture 2020, 10, 28. https://doi.org/10.3390/agriculture10020028
Azad MOK, Kjaer KH, Adnan M, Naznin MT, Lim JD, Sung IJ, Park CH, Lim YS. The Evaluation of Growth Performance, Photosynthetic Capacity, and Primary and Secondary Metabolite Content of Leaf Lettuce Grown under Limited Irradiation of Blue and Red LED Light in an Urban Plant Factory. Agriculture. 2020; 10(2):28. https://doi.org/10.3390/agriculture10020028
Chicago/Turabian StyleAzad, Md Obyedul Kalam, Katrine Heinsvig Kjaer, Md Adnan, Most Tahera Naznin, Jung Dae Lim, In Je Sung, Cheol Ho Park, and Young Seok Lim. 2020. "The Evaluation of Growth Performance, Photosynthetic Capacity, and Primary and Secondary Metabolite Content of Leaf Lettuce Grown under Limited Irradiation of Blue and Red LED Light in an Urban Plant Factory" Agriculture 10, no. 2: 28. https://doi.org/10.3390/agriculture10020028