Planting Density Effects on Grow Rate, Biometric Parameters, and Biomass Calorific Value of Selected Trees Cultivated as SRC


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Characteristics and Experimental Design
- Poplar (Populs L.), AF2 variety;
- Siberian elm (Ulmus pumila);
- Black alder (Alnus glutinosa);
- White birch (Betula pubecsens);
- Boxelder maple (Acer negundo);
- Silver maple (Acer saccharinum L.).
2.2. Biomass Analyses
2.3. Statistical Analyses
3. Results and Discussion
3.1. Green Mass
3.2. Dry Mass
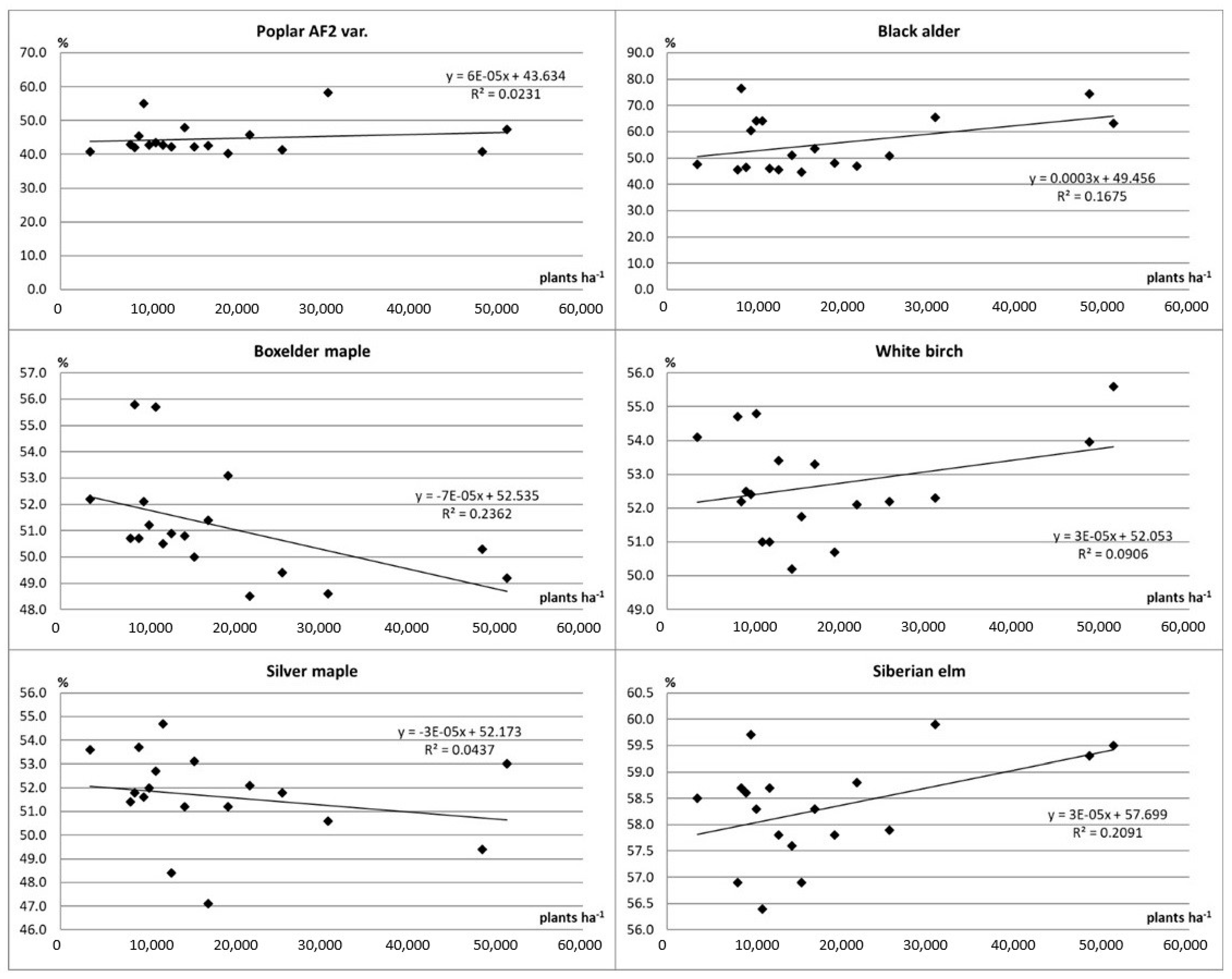
3.3. Share of Dry Mass
3.4. Potential Yield of Dry Mass
3.5. Height of Plants
3.6. Shoots Diameter
3.7. Higher Heating Value
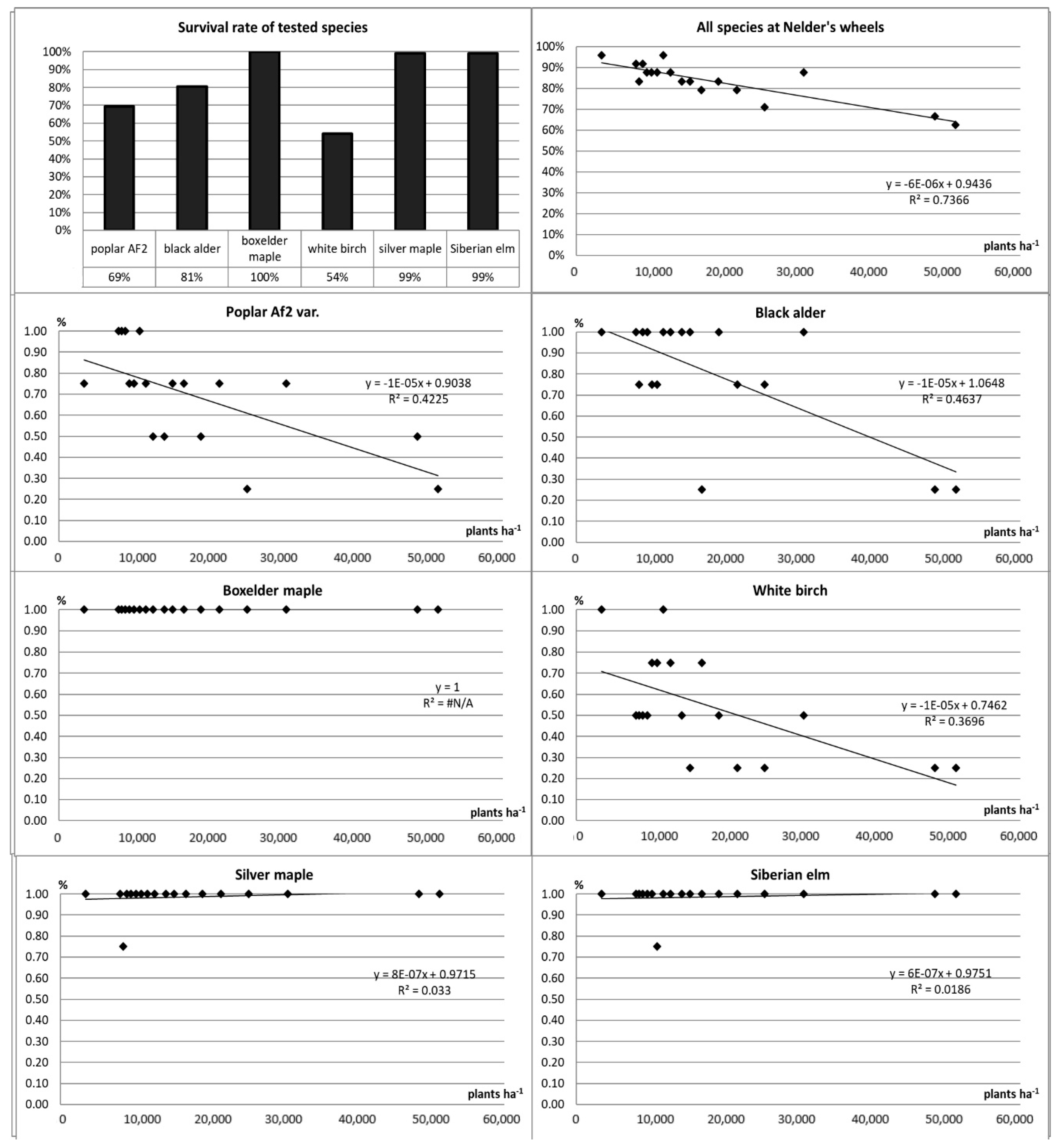
3.8. Survival Rate
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Diamantidis, N.D.; Koukis, E.G. Agricultural crops and residues as feedstock for non-food products in Western Europe. Ind. Crops Prod. 2000, 11, 97–106. [Google Scholar] [CrossRef]
- El Kasmioui, O.; Ceulemans, R. Financial analysis of the cultivation of poplar and willow for bioenergy. Biomass Bioenergy 2012, 43, 52–64. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Śnieg, M.; Krzyżaniak, M.; Tworkowski, J.; Szczukowski, S. Short rotation coppices, grasses and other herbaceous crops: Productivity and yield energy value versus 26 genotypes (2018). Biomass Bioenergy 2018, 119, 109–120. [Google Scholar] [CrossRef]
- Nabel, M.; Barbosa, D.B.P.; Horsch, D.; Jablonowski, N.D. Energy crop (Sida hermaphrodita) fertilization using digestate under marginal conditions: A dose-response experiment. Energy Procedia 2014, 54, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Matyka, M.; Kuś, J. Influence of soil quality for yield and biometric features of Sida hermaphrodita L. Rusby. Pol. J. Environ. Stud. 2018, 27, 2669–2675. [Google Scholar] [CrossRef]
- Navarro, A.; Facciotto, G.; Campi, P.; Mastrorilli, M. Physiological adaptations of five poplar genotypes grown under SRC in the semi-arid Mediterranean environment. Trees 2014, 28, 983–994. [Google Scholar] [CrossRef]
- Benomar, L.; DesRochers, A.; Larocque, G. The effects of spacing on growth, morphology and biomass production and allocation in two hybrid poplar clones growing in the boreal region of Canada. Trees 2012, 26, 939–949. [Google Scholar] [CrossRef]
- Toillon, J.; Fichot, R.; Dallé, E.; Berthelot, A.; Brignolas, F.; Marron, N. Planting density affects growth and water-use efficiency depending on site in Populus deltoids x P. nigra. For. Ecol. Manag. 2013, 304, 345–354. [Google Scholar] [CrossRef]
- Nelder, J.A. New kinds of systematic design for spacing experiments. Biometrics 1962, 18, 283–307. [Google Scholar] [CrossRef]
- Wilkinson, J.M.; Evans, E.J.; Bilsborrow, P.E.; Wright, C.; Hewison, W.O.; Pilbeam, D.J. Yield of willow cultivars at different planting densities in a commercial short rotation coppice in the north of England. Biomass Bioenergy 2007, 31, 469–474. [Google Scholar] [CrossRef]
- Walle, I.V.; Van Camp, N.; Van de Casteele, L.; Verheyen, K.; Lemeur, R. Short-rotation forestry of birch, maple, poplar and willow in Flanders (Belgium) I—Biomass production after 4 years of tree growth. Biomass Bioenergy 2007, 31, 267–275. [Google Scholar] [CrossRef]
- Bullard, M.J.; Mustill, S.J.; McMillan, S.D.; Nixon, P.M.; Carver, P.; Britt, C.P. Yield improvements through modification of planting density and harvest frequency in short rotation coppice Salix spp.—1. Yield response in two morphologically diverse varieties. Biomass Bioenergy 2002, 22, 15–25. [Google Scholar] [CrossRef]
- Dahlhausen, J.; Uhl, E.; Heym, M.; Biber, P.; Ventura, M.; Panzacchi, P.; Tonon, G.; Horváth, T.; Pretzsch, H. Stand density sensitive biomass functions for young oak trees at four different European sites. Trees 2017, 31, 1811–1826. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Szczukowski, S.; Tworkowski, J.; Wróblewska, H.; Krzyżaniak, M. Short rotation willow coppice biomass as an industrial and energy feedstock. Ind. Crops Prod. 2011, 33, 217–223. [Google Scholar] [CrossRef]
- Kulig, B.; Gacek, E.; Wojciechowski, R.; Oleksy, A.; Kołodziejczyk, M.; Szewczyk, W.; Klimek-Kopyra, A. Biomass yield and energy efficiency of willow depending on cultivar, harvesting frequency and planting density. Plant. Soil Environ. 2019, 65, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Elfeel, A.A.; Elmagboul, A.H. Effect of planting density on eucaena leucocephala forage and Woody stems production under arid dry climate. Int. J. Agric. Res. 2016, 2, 7–11. [Google Scholar]
- Achinelli, F.G.; Doffo, G.; Barotto, A.J.; Luquez, V.; Monteoliva, S. Effects of irrigation, plantation density and clonal composition on woody biomass quality for bioenergy in a short rotation culture system with willows (Salix spp.). Rev. Árvore 2018, 42, 1–8. [Google Scholar] [CrossRef]
- Geyer, W.A.; Argent, R.M.; Walawender, W.P. Biomass properties and gasification behavior of 7-year-old Siberian elm. Wood Fiber Sci. 1987, 19, 176–182. [Google Scholar]
- Geyer, W.A.; Walawender, W.P. Biomass properties and gasification behavior of young silver maple trees. Wood Fiber Sci. 1997, 29, 85–90. [Google Scholar]
- Geyer, W.A.; Walawender, W.P. Biomass properties and gasification behavior of young black locust. Wood Fiber Sci. 1994, 26, 354–359. [Google Scholar]
- Niemczyk, M.; Kaliszewski, A.; Jewiarz, M.; Wróbel, M.; Mudryk, K. Productivity and biomass characteristics of selected poplar (Populus spp.) cultivars under the climatic conditions of northern Poland. Biomass Bioenergy 2018, 111, 46–51. [Google Scholar] [CrossRef]
- Truax, B.; Fortier, J.; Gagnon, D.; Lambert, F. Planting density and site effects on stem dimensions, stand productivity, biomass partitioning, carbon stocks and soil nutrient supply in hybrid poplar plantations. Forests 2018, 9, 293. [Google Scholar] [CrossRef] [Green Version]
- Stolarski, M.J.; Szczukowski, S.; Tworkowski, J.; Krzyżaniak, M.; Załuski, D. Willow production during 12 consecutive years—The effects of harvest rotation, planting density and cultivar on biomass yield. Glob. Chang. Biol. Bioenergy 2019, 11, 635–656. [Google Scholar] [CrossRef]
- Geyer, W.A.; Barden, C.; Preece, J.E. Biomass and wood properties of young silver maple clones. Wood Fiber Sci. 2008, 40, 23–28. [Google Scholar]
- Perez, I.; Perez, J.; Carrasco, J.; Ciria, P. Siberian elm responses to different culture conditions under short rotation forestry in Mediterranean areas. Turk. J. Agric. For. 2014, 38, 652–662. [Google Scholar] [CrossRef]
- Di Matteo, G.; Sperandio, G.; Verani, S. Field performance of poplar for bioenergy in southern Europe after two coppicing rotations: Effects of clone and planting density. iForest Biogeosci. For. 2012, 5, 224. [Google Scholar] [CrossRef] [Green Version]
- Hummel, S. Height, diameter and crown dimensions of Cordia alliodora associated with tree density. For. Ecol. Manag. 2000, 127, 31–40. [Google Scholar] [CrossRef]
- Mehtätalo, L.; de-Miguel, S.; Gregoire, T.G. Modeling height-diameter curves for prediction. Can. J. For. Res. 2015, 45, 826–837. [Google Scholar] [CrossRef] [Green Version]
- Castaño-Díaz, M.; Barrio-Anta, M.; Afif-Khouri, E.; Cámara-Obregón, A. Willow short rotation coppice trial in a former mining area in northern Spain: Effects of clone, fertilization and planting density on yield after five years. Forests 2018, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Sabatti, M.; Fabbrini, F.; Harfouche, A.; Beritognolo, I.; Mareschi, L.; Carlini, M.; Paris, P.; Scarascia-Mugnozza, G. Evaluation of biomass production potential and heating value of hybrid poplar genotypes in a short-rotation culture in Italy. Ind. Crops Prod. 2014, 61, 62–73. [Google Scholar] [CrossRef]
- Navarro, A.; Stellacci, A.M.; Campi, P.; Vitti, C.; Modugno, F.; Mastrorilli, M. Feasibility of SRC species for growing in Mediterranean conditions. Bioenergy Res. 2016, 9, 208–223. [Google Scholar] [CrossRef]
- Nebeská, D.; Trögl, J.; Žofková, D.; Voslařová, A.; Štojdl, J.; Pidlisnyuk, V. Calorific values of miscanthus x giganteus biomass cultivated under suboptimal conditions in marginal soils. Stud. Oecol. 2019, 13, 61–67. [Google Scholar] [CrossRef]
- Kumar, R.; Pandey, K.K.; Chandrashekar, N.; Mohan, S. Study of age and height wise variability on calorific value and other fuel properties of Eucalyptus hybrid, Acacia auriculaeformis and Casuarina equisetifolia. Biomass Bioenergy 2011, 35, 1339–1344. [Google Scholar] [CrossRef]
- Trnka, M.; Trnka, M.; Fialová, J.; Koutecky, V.; Fajman, M.; Zalud, Z.; Hejduk, S. Biomass production and survival rates of selected poplar clones grown under a short-rotation system on arable land. Plant Soil Environ. 2008, 54, 78–88. [Google Scholar] [CrossRef] [Green Version]
- Geyer, W.A. Biomass production in the central great plains USA under various coppice regimes. Biomass Bioenergy 2006, 30, 778–783. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ring No. | Distance to the Center [m] | Distance to Previous Ring [m] | Density [Plants ha−1] |
|---|---|---|---|
| 1 | 1.0 | Guard ring | |
| 2 | 1.5 | 0.5 | 51,282 |
| 3 | 2.0 | 0.5 | 48,462 |
| 4 | 2.5 | 0.5 | 30,769 |
| 5 | 3.0 | 0.5 | 25,477 |
| 6 | 3.5 | 0.5 | 21,739 |
| 7 | 4.0 | 0.5 | 19,231 |
| 8 | 4.5 | 0.5 | 16,949 |
| 9 | 5.0 | 0.5 | 15,384 |
| 10 | 5.5 | 0.5 | 14,286 |
| 11 | 6.0 | 0.5 | 12,739 |
| 12 | 6.5 | 0.5 | 11,764 |
| 13 | 7.0 | 0.5 | 10,929 |
| 14 | 7.5 | 0.5 | 10,204 |
| 15 | 8.0 | 0.5 | 9569 |
| 16 | 8.5 | 0.5 | 9009 |
| 17 | 9.0 | 0.5 | 8510 |
| 18 | 9.5 | 0.5 | 8064 |
| 19 | 10.0 | Guard ring | |
| 20 | 11.0 | 1.0 | 3448 |
| 21 | 12.0 | Guard ring |
| Specification | Green Mass [kg] | Share of Dry Mass [%] | Dry Mass [kg] | Plant Height [m] | Shoot Diameter [mm] | Higher Heating Value [J g−1] | Potential Yield of Dry Mass [t ha−1 yr−1] |
|---|---|---|---|---|---|---|---|
| Poplar AF2 var. | 29.1 a * | 42.8 a | 13.1 a | 10.6 a | 113.5 a | 17,908 a | 15.8 a |
| Siberian elm | 2.8 b | 58.5 b | 1.7 b | 4.7 d | 41.8 b | 18,664 bc | 5.2 b |
| Black alder | 5.3 bc | 48.2 c | 2.5 bc | 6.6 bc | 60.4 c | 18,366 ab | 3.9 b |
| White birch | 5.5 bc | 52.4 c | 2.9 bc | 7.0 b | 59.4 bc | 19,509 c | 2.2 b |
| Boxelder maple | 9.3 c | 50.8 c | 4.7 c | 6.8 b | 63.8 c | 19,158 c | 9.8 c |
| Silver maple | 2.6 b | 51.8 c | 1.4 b | 5.2 cd | 39.2 b | 18,726 bc | 4.7 b |
| Plant | Specification | Green Mass [kg Plant −1] | Share of Dry Mass [%] | Dry Mass [kg Plant −1] | Plant Height [m] | Shoot Diameter [mm] | Higher Heating Value [J g−1] | Potential Yield of Dry Mass [t ha−1 yr−1] |
|---|---|---|---|---|---|---|---|---|
| Poplar AF2 var. | Plant density [plants ha−1] | −0.619 * | 0.152 | −0.651 | −0.384 | −0.367 | −0.485 | 0.025 |
| Green mass [kg] | −0.542 | 0.993 | 0.822 | 0.815 | 0.435 | 0.377 | ||
| Share of dry matter [%] | −0.454 | −0.650 | −0.389 | −0.350 | −0.131 | |||
| Dry mass [kg plant −1] | 0.797 | 0.796 | 0.428 | 0.399 | ||||
| Plant height [m] | 0.676 | 0.443 | 0.525 | |||||
| Shoot diameter [mm] | 0.346 | 0.130 | ||||||
| Higher heating value [J g−1] | 0.086 | |||||||
| Siberian elm | Plant density [plants ha−1] | −0.660 | 0.457 | −0.663 | −0.402 | −0.628 | 0.248 | 0.414 |
| Green mass [kg] | −0.504 | 1.000 | 0.774 | 0.923 | −0.065 | 0.309 | ||
| Share of dry matter [%] | −0.479 | −0.376 | −0.439 | 0.356 | −0.087 | |||
| Dry mass [kg plants −1] | 0.777 | 0.925 | −0.061 | 0.306 | ||||
| Plant height [m] | 0.793 | −0.023 | 0.518 | |||||
| Shoot diameter [mm] | −0.124 | 0.234 | ||||||
| Higher heating value [J g−1] | 0.235 | |||||||
| Black alder | Plant density [plants ha−1] | −0.833 | 0.409 | −0.804 | −0.775 | −0.874 | 0.115 | −0.539 |
| Green mass [kg] | −0.357 | 0.929 | 0.616 | 0.884 | −0.166 | 0.437 | ||
| Share of dry matter [%] | −0.025 | −0.257 | −0.293 | −0.317 | −0.257 | |||
| Dry mass [kg plant −1] | 0.603 | 0.850 | −0.304 | 0.437 | ||||
| Plant height [m] | 0.841 | −0.129 | 0.346 | |||||
| Shoot diameter [mm] | −0.129 | 0.280 | ||||||
| Higher heating value [J g−1] | −0.149 | |||||||
| White birch | Plant density [plants ha−1] | −0.589 | 0.301 | −0.589 | −0.188 | −0.639 | −0.095 | −0.175 |
| Green mass [kg] | −0.278 | 0.998 | 0.598 | 0.679 | −0.139 | 0.728 | ||
| Share of dry matter [%] | −0.227 | −0.457 | −0.175 | −0.520 | −0.185 | |||
| Dry mass [kg plant −1] | 0.588 | 0.684 | −0.170 | 0.727 | ||||
| Plant height [m] | 0.563 | 0.062 | 0.434 | |||||
| Shoot diameter [mm] | 0.169 | 0.233 | ||||||
| Higher heating value [J g−1] | −0.288 | |||||||
| Boxelder maple | Plant density [plants ha−1] | −0.684 | −0.486 | −0.670 | −0.642 | −0.706 | 0.321 | 0.493 |
| Green mass [kg] | 0.185 | 1.000 | 0.587 | 0.956 | −0.388 | −0.068 | ||
| Share of dry matter [%] | 0.245 | 0.119 | 0.229 | −0.109 | −0.360 | |||
| Dry mass [kg plant −1] | 0.580 | 0.957 | −0.395 | −0.095 | ||||
| Plant height [m] | 0.630 | −0.163 | 0.121 | |||||
| Shoot diameter [mm] | −0.313 | −0.130 | ||||||
| Higher heating value [J g−1] | 0.196 | |||||||
| Silver maple | Plant density [plants ha−1] | −0.590 | −0.209 | −0.601 | −0.124 | −0.621 | −0.064 | 0.378 |
| Green mass [kg] | 0.041 | 0.994 | 0.567 | 0.865 | −0.112 | 0.326 | ||
| Share of dry matter [%] | 0.144 | −0.450 | −0.140 | −0.172 | −0.281 | |||
| Dry mass [kg plant −1] | 0.518 | 0.839 | −0.121 | 0.295 | ||||
| Plant height [m] | 0.668 | −0.087 | 0.549 | |||||
| Shoot diameter [mm] | −0.013 | 0.197 | ||||||
| Higher heating value [J g−1] | −0.539 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berbeć, A.K.; Matyka, M. Planting Density Effects on Grow Rate, Biometric Parameters, and Biomass Calorific Value of Selected Trees Cultivated as SRC. Agriculture 2020, 10, 583. https://doi.org/10.3390/agriculture10120583
Berbeć AK, Matyka M. Planting Density Effects on Grow Rate, Biometric Parameters, and Biomass Calorific Value of Selected Trees Cultivated as SRC. Agriculture. 2020; 10(12):583. https://doi.org/10.3390/agriculture10120583
Chicago/Turabian StyleBerbeć, Adam Kleofas, and Mariusz Matyka. 2020. "Planting Density Effects on Grow Rate, Biometric Parameters, and Biomass Calorific Value of Selected Trees Cultivated as SRC" Agriculture 10, no. 12: 583. https://doi.org/10.3390/agriculture10120583