
A Manifesto in Defense of Pain Complexity: A Critical Review of Essential Insights in Pain Neuroscience

{kind=link}
Abstract
:1. Introduction
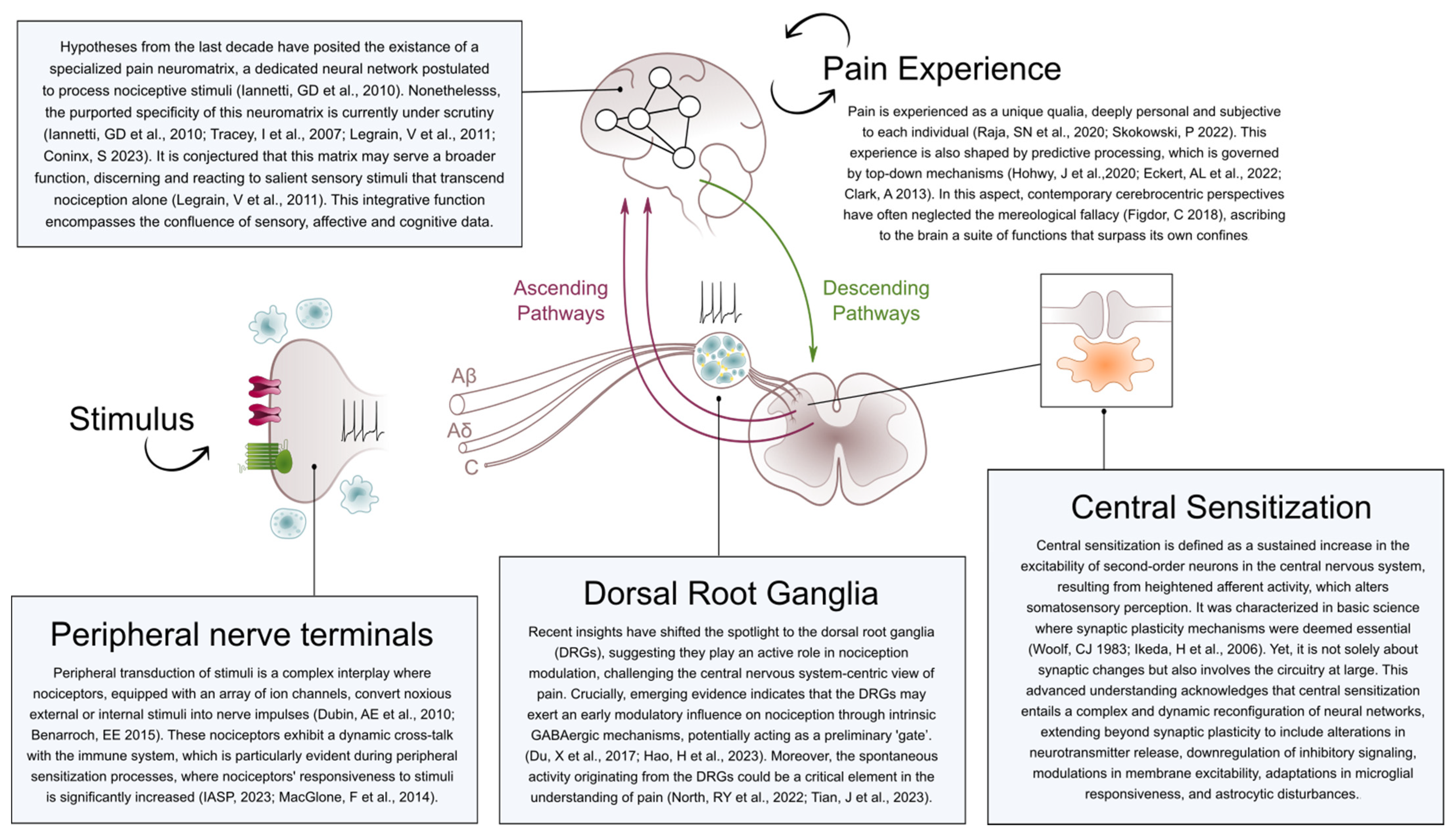
2. Pain and Brain: A Mereological Fallacy We Should Abandon
3. Discerning the Ambiguity between Pain and Nociception: A Crucial Source of Misnomers
4. Starting from the Periphery: Does Pain Modulation Begin in the Central Nervous System?
5. Central Sensitization as a Focus of Confusion: Weaving Threads of Uncertainty
6. Cortical Processing: Does the Pain Neuromatrix Really Exist? A Controversial Simplification
7. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Todd, K.H.; Ducharme, J.; Choiniere, M.; Crandall, C.S.; Fosnocht, D.E.; Homel, P.; Tanabe, P.; PEMI Study Group. Pain in the emergency department: Results of the pain and emergency medicine initiative (PEMI) multicenter study. J. Pain. 2007, 8, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Hübscher, M.; Moseley, G.L.; Kamper, S.J.; Traeger, A.C.; Mansell, G.; McAuley, J.H. How does pain lead to disability? A systematic review and meta-analysis of mediation studies in people with back and neck pain. Pain 2015, 156, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Vos, T.; Flaxman, A.D.; Naghavi, M.; Lozano, R.; Michaud, C.; Ezzati, M.; Shibuya, K.; Salomon, J.A.; Abdalla, S.; Aboyans, V.; et al. Years lived with disability (YLDs) for 1160 sequelae of 289 diseases and injuries 1990–2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2163–2196. [Google Scholar] [CrossRef] [PubMed]
- Raffaeli, W.; Tenti, M.; Corraro, A.; Malafoglia, V.; Ilari, S.; Balzani, E.; Bonci, A. Chronic Pain: What Does It Mean? A Review on the Use of the Term Chronic Pain in Clinical Practice. J. Pain Res. 2021, 14, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Treede, R.D.; Rief, W.; Barke, A.; Aziz, Q.; Bennett, M.I.; Benoliel, R.; Cohen, M.; Evers, S.; Finnerup, N.B.; First, M.B.; et al. A classification of chronic pain for ICD-11. Pain 2015, 156, 1003–1007. [Google Scholar] [CrossRef]
- Deyo, R.A.; Mirza, S.K.; Turner, J.A.; Martin, B.I. Overtreating chronic back pain: Time to back off? J. Am. Board Fam. Med. JABFM 2009, 22, 62–68. [Google Scholar] [CrossRef]
- Bushnell, M.C.; Frangos, E.; Madian, N. Non-pharmacological Treatment of Pain: Grand Challenge and Future Opportunities. Front. Pain Res. 2021, 2, 696783. [Google Scholar] [CrossRef]
- Tang, S.K.; Tse, M.M.Y.; Leung, S.F.; Fotis, T. The effectiveness, suitability, and sustainability of non-pharmacological methods of managing pain in community-dwelling older adults: A systematic review. BMC Public Health 2019, 19, 1488. [Google Scholar] [CrossRef]
- Foster, N.E.; Anema, J.R.; Cherkin, D.; Chou, R.; Cohen, S.P.; Gross, D.P.; Ferreira, P.H.; Fritz, J.M.; Koes, B.W.; Peul, W.; et al. Prevention and treatment of low back pain: Evidence, challenges, and promising directions. Lancet 2018, 391, 2368–2383. [Google Scholar] [CrossRef]
- Macfarlane, G.J.; Kronisch, C.; Dean, L.E.; Atzeni, F.; Häuser, W.; Fluß, E.; Choy, E.; Kosek, E.; Amris, K.; Branco, J.; et al. EULAR revised recommendations for the management of fibromyalgia. Ann. Rheum. Dis. 2017, 76, 318–328. [Google Scholar] [CrossRef]
- National Institute for Health and Care Excellence (NICE). Chronic Pain (Primary and Secondary) in Over 16s: Assessment of All Chronic Pain and Management of Chronic Primary Pain; National Institute for Health and Care Excellence (NICE): London, UK, 2021; NICE Evidence Reviews Collection; ISBN 978-1-4731-4067-7. Available online: https://www.nice.org.uk/guidance/ng193 (accessed on 25 May 2023).
- Watson, J.A.; Ryan, C.G.; Cooper, L.; Ellington, D.; Whittle, R.; Lavender, M.; Dixon, J.; Atkinson, G.; Cooper, K.; Martin, D.J. Pain Neuroscience Education for Adults With Chronic Musculoskeletal Pain: A Mixed-Methods Systematic Review and Meta-Analysis. J. Pain 2019, 20, 1140.e1–1140.e22. [Google Scholar] [CrossRef] [PubMed]
- Butler, D.S.; Moseley, G.L. Explain Pain, 2nd ed.; Noigroup Publications: Adelaide, Australia, 2013. [Google Scholar]
- Engel, G.L. The need for a new medical model: A challenge for biomedicine. Science 1977, 196, 129–136. [Google Scholar] [CrossRef] [PubMed]
- McClelland, J.L.; Cleeremans, A. Connectionist models. In The Oxford Companion to Consciousness; Bayne, T., Cleeremans, A., Wilken, P., Eds.; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Moseley, G.L.; Brhyn, L.; Ilowiecki, M.; Solstad, K.; Hodges, P.W. The threat of predictable and unpredictable pain: Differential effects on central nervous system processing? Aust. J. Physiother. 2003, 49, 263–267. [Google Scholar] [CrossRef]
- Bain, D. Evaluativist Accounts of Pain’s Unpleasantness. In The Routledge Handbook of the Philosophy of Pain; Corns, J., Ed.; Routledge: London, UK, 2017; pp. 40–50. [Google Scholar]
- Ploner, M.; Lee, M.C.; Wiech, K.; Bingel, U.; Tracey, I. Prestimulus functional connectivity determines pain perception in humans. Proc. Natl. Acad. Sci. USA 2010, 107, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Mitsi, V.; Zachariou, V. Modulation of pain, nociception, and analgesia by the brain reward center. Neuroscience 2016, 338, 81–92. [Google Scholar] [CrossRef]
- Labrakakis, C. The Role of the Insular Cortex in Pain. Int. J. Mol. Sci. 2023, 24, 5736. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, Z.; Zhong, J.; Zhang, Y.; Lin, X.; Cai, H.; Kong, Y. Thalamocortical Mechanisms for Nostalgia-Induced Analgesia. J. Neurosci. 2022, 42, 2963–2972. [Google Scholar] [CrossRef]
- Kim, J.; Kang, I.; Chung, Y.-A.; Kim, T.-S.; Namgung, E.; Lee, S.; Oh, J.K.; Jeong, H.S.; Cho, H.; Kim, M.J.; et al. Altered attentional control over the salience network in complex regional pain syndrome. Sci. Rep. 2018, 8, 7466. [Google Scholar] [CrossRef]
- Figdor, C. The Nonsense View. In Pieces of Mind: The Proper Domain of Psychological Predicates; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Harré, R. Behind the Mereological Fallacy. Philosophy 2012, 87, 329–352. [Google Scholar] [CrossRef]
- Bich, L. Autonomous Systems and the Place of Biology Among Sciences. Perspectives for an Epistemology of Complex Systems. In Multiplicity and Interdisciplinarity. Contemporary Systems Thinking; Minati, G., Ed.; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Ma’ayan, A. Complex systems biology. J. R. Soc. Interface 2017, 14, 20170391. [Google Scholar] [CrossRef]
- Flor, H.; Elbert, T.; Mühlnickel, W.; Pantev, C.; Wienbruch, C.; Taub, E. Cortical reorganization and phantom phenomena in congenital and traumatic upper-extremity amputees. Exp. Brain Res. 1998, 119, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Collins, K.L.; Russell, H.G.; Schumacher, P.J.; Robinson-Freeman, K.E.; O’Conor, E.C.; Gibney, K.D.; Yambem, O.; Dykes, R.W.; Waters, R.S.; Tsao, J.W. A review of current theories and treatments for phantom limb pain. J. Clin. Investig. 2018, 128, 2168–2176. [Google Scholar] [CrossRef] [PubMed]
- Vaso, A.; Adahan, H.M.; Gjika, A.; Zahaj, S.; Zhurda, T.; Vyshka, G.; Devor, M. Peripheral nervous system origin of phantom limb pain. Pain 2014, 155, 1384–1391. [Google Scholar] [CrossRef]
- Borghi, B.; D’Addabbo, M.; White, P.F.; Gallerani, P.; Toccaceli, L.; Raffaeli, W.; Tognù, A.; Fabbri, N.; Mercuri, M. The use of prolonged peripheral neural blockade after lower extremity amputation: The effect on symptoms associated with phantom limb syndrome. Anesth. Analg. 2010, 111, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Quintner, J. Why Are Women with Fibromyalgia so Stigmatized? Pain Med. 2020, 21, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Nijs, J.; George, S.Z.; Clauw, D.J.; Fernández-de-las-Peñas, C.; Kosek, E.; Ickmans, K.; Fernández-Carnero, J.; Polli, A.; Kapreli, E.; Huysmans, E.; et al. Central sensitisation in chronic pain conditions: Latest discoveries and their potential for precision medicine. Lancet Rheumatol. 2021, 3, e383–e392. [Google Scholar] [CrossRef]
- Martínez-Lavín, M. Centralized nociplastic pain causing fibromyalgia: An emperor with no cloths? Clin. Rheumatol. 2022, 41, 3915–3917. [Google Scholar] [CrossRef]
- Grayston, R.; Czanner, G.; Elhadd, K.; Goebel, A.; Frank, B.; Üçeyler, N.; Malik, R.A.; Alam, U. A systematic review and meta-analysis of the prevalence of small fiber pathology in fibromyalgia: Implications for a new paradigm in fibromyalgia etiopathogenesis. Semin. Arthritis Rheum. 2019, 48, 933–940. [Google Scholar] [CrossRef]
- Coskun Benlidayi, I. Role of inflammation in the pathogenesis and treatment of fibromyalgia. Rheumatol. Int. 2019, 39, 781–791. [Google Scholar] [CrossRef]
- Olausson, P.; Ghafouri, B.; Ghafouri, N.; Gerdle, B. Specific proteins of the trapezius muscle correlate with pain intensity and sensitivity-an explorative multivariate proteomic study of the trapezius muscle in women with chronic widespread pain. J. Pain Res. 2016, 9, 345–356. [Google Scholar] [CrossRef]
- Wåhlén, K.; Ernberg, M.; Kosek, E.; Mannerkorpi, K.; Gerdle, B.; Ghafouri, B. Significant correlation between plasma proteome profile and pain intensity, sensitivity, and psychological distress in women with fibromyalgia. Sci. Rep. 2020, 10, 12508. [Google Scholar] [CrossRef] [PubMed]
- Erdrich, S.; Hawrelak, J.A.; Myers, S.P.; Harnett, J.E. Determining the association between fibromyalgia, the gut microbiome and its biomarkers: A systematic review. BMC Musculoskelet. Disord. 2020, 21, 181. [Google Scholar] [CrossRef] [PubMed]
- Coderre, T.J. Contribution of microvascular dysfunction to chronic pain. Front. Pain Res. 2023, 4, 1111559. [Google Scholar] [CrossRef] [PubMed]
- Rittner, H.L.; Brack, A.; Stein, C. Pain and the immune system. Br. J. Anaesth. 2008, 101, 40–44. [Google Scholar] [CrossRef]
- Moncrieff, J.; Cooper, R.E.; Stockmann, T.; Amendola, S.; Hengartner, M.P.; Horowitz, M.A. The serotonin theory of depression: A systematic umbrella review of the evidence. Mol. Psychiatry 2022, 28, 3243–3256. [Google Scholar] [CrossRef]
- Lee, M.; Silverman, S.M.; Hansen, H.; Patel, V.B.; Manchikanti, L. A comprehensive review of opioid-induced hyperalgesia. Pain Physician. 2011, 14, 145–161. [Google Scholar] [CrossRef]
- Ferreira, G.E.; Abdel-Shaheed, C.; Underwood, M.; Finnerup, N.B.; Day, R.O.; McLachlan, A.; Eldabe, S.; Zadro, J.R.; Maher, C.G. Efficacy, safety, and tolerability of antidepressants for pain in adults: Overview of systematic reviews. BMJ Clin. Res. Ed. 2023, 380, e072415. [Google Scholar] [CrossRef]
- Carvalho, A.F.; Sharma, M.S.; Brunoni, A.R.; Vieta, E.; Fava, G.A. The Safety, Tolerability and Risks Associated with the Use of Newer Generation Antidepressant Drugs: A Critical Review of the Literature. Psychother. Psychosom. 2016, 85, 270–288. [Google Scholar] [CrossRef]
- Kaczmarek, E. How to distinguish medicalization from over-medicalization? Med. Health Care Philos. 2019, 22, 119–128. [Google Scholar] [CrossRef]
- Paice, J.A.; Von Roenn, J.H. Under- or overtreatment of pain in the patient with cancer: How to achieve proper balance. J. Clin. Oncol. 2014, 32, 1721–1726. [Google Scholar] [CrossRef]
- Raja, S.N.; Carr, D.B.; Cohen, M.; Finnerup, N.B.; Flor, H.; Gibson, S.; Keefe, F.J.; Mogil, J.S.; Ringkamp, M.; Sluka, K.A.; et al. The Revised International Association for the Study of Pain Definition of Pain: Concepts, Challenges, and Compromises. Pain 2020, 161, 1976–1982. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Weisman, A.; Quintner, J. Pain is Not a “Thing”: How That Error Affects Language and Logic in Pain Medicine. J. Pain 2022, 23, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Louw, A.; Nijs, J.; Puentedura, E.J. A Clinical Perspective on a Pain Neuroscience Education Approach to Manual Therapy. J. Man. Manip. Ther. 2017, 25, 160–168. [Google Scholar] [CrossRef]
- Saposnik, G.; Redelmeier, D.; Ruff, C.C.; Tobler, P.N. Cognitive Biases Associated with Medical Decisions: A Systematic Review. BMC Med. Inform. Decis. Mak. 2016, 16, 138. [Google Scholar] [CrossRef] [PubMed]
- Feldman, J. The Simplicity Principle in Perception and Cognition. Wiley Interdiscip. Rev. Cogn. Sci. 2016, 7, 330–340. [Google Scholar] [CrossRef]
- Fares, W.H. The ‘Availability’ Bias: Underappreciated but with Major Potential Implications. Crit. Care 2014, 18, 118. [Google Scholar] [CrossRef] [PubMed]
- Skokowski, P. Sensing Qualia. Front. Syst. Neurosci. 2022, 16, 795405. [Google Scholar] [CrossRef]
- Chalmers, D.J. Facing Up to the Problem of Consciousness. J. Conscious. Stud. 1995, 2, 200–219. [Google Scholar]
- Moreira-Almeida, A.; Araujo, S.F.; Cloninger, C.R. The Presentation of the Mind-Brain Problem in Leading Psychiatry Journals. Rev. Bras. Psiquiatr. 2018, 40, 335–342. [Google Scholar] [CrossRef]
- International Association for the Study of Pain. Terminology. Available online: https://www.iasp-pain.org/resources/terminology/ (accessed on 25 May 2023).
- Savitha, S.A.; Sacchidanand, S.A.; Gowda, S.K. Misnomers in Dermatology: An Update. Indian J. Dermatol. 2013, 58, 467–474. [Google Scholar] [CrossRef]
- Fisher, J.P.; Hassan, D.T.; Connor, N.O. Minerva. BMJ 1995, 310, 70. [Google Scholar] [CrossRef]
- Reichling, D.B.; Green, P.G.; Levine, J.D. The Fundamental Unit of Pain is the Cell. Pain 2013, 154 (Suppl. S1), S2–S9. [Google Scholar] [CrossRef]
- Sneddon, L.U. Comparative Physiology of Nociception and Pain. Physiology 2018, 33, 63–73. [Google Scholar] [CrossRef]
- Tobimatsu, S. Understanding Cortical Pain Perception in Humans. Neurol. Clin. Neurosci. 2021, 9, 24–29. [Google Scholar] [CrossRef]
- Roy, M.; Piché, M.; Chen, J.I.; Peretz, I.; Rainville, P. Cerebral and Spinal Modulation of Pain by Emotions. Proc. Natl. Acad. Sci. USA 2009, 106, 20900–20905. [Google Scholar] [CrossRef] [PubMed]
- Tracy, L.M.; Gibson, S.J.; Georgiou-Karistianis, N.; Giummarra, M.J. Effects of Explicit Cueing and Ambiguity on the Anticipation and Experience of a Painful Thermal Stimulus. PLoS ONE 2017, 12, e0183650. [Google Scholar] [CrossRef]
- Boadas-Vaello, P.; Castany, S.; Homs, J.; Álvarez-Pérez, B.; Deulofeu, M.; Verdú, E. Neuroplasticity of Ascending and Descending Pathways after Somatosensory System Injury: Reviewing Knowledge to Identify Neuropathic Pain Therapeutic Targets. Spinal Cord 2016, 54, 330–340. [Google Scholar] [CrossRef]
- Melzack, R.; Wall, P.D. Pain Mechanisms: A New Theory. Science 1965, 150, 971–979. [Google Scholar] [CrossRef]
- Moseley, G.L.; Pearson, N.; Reezigt, R.; Madden, V.J.; Hutchinson, M.R.; Dunbar, M.; Beetsma, A.J.; Leake, H.B.; Moore, P.; Simons, L.; et al. Considering Precision and Utility When we Talk About Pain. Comment on Cohen et al. J. Pain. 2023, 24, 178–181. [Google Scholar] [CrossRef]
- Cohen, M.; Weisman, A.; Quintner, J. Response to van Rysewyk S and Moseley GL et al.’s Comments on Cohen et al. J Pain 2022; 23(8):1283–1293. J. Pain. 2023, 24, 184–185. [Google Scholar] [CrossRef]
- Descartes, R.; Quintás, G. Traité de l’homme (El Tratado del Hombre); Alianza Editorial: Madrid, Spain, 1990. [Google Scholar]
- Dubin, A.E.; Patapoutian, A. Nociceptors: The Sensors of the Pain Pathway. J. Clin. Investig. 2010, 120, 3760–3772. [Google Scholar] [CrossRef] [PubMed]
- Benarroch, E.E. Ion Channels in Nociceptors: Recent Developments. Neurology 2015, 84, 1153–1164. [Google Scholar] [CrossRef] [PubMed]
- Djouhri, L.; Lawson, S.N. Abeta-fiber Nociceptive Primary Afferent Neurons: A Review of Incidence and Properties in Relation to Other Afferent A-fiber Neurons in Mammals. Brain Res. Rev. 2004, 46, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Lechner, S. An Update on the Spinal and Peripheral Pathways of Pain Signalling. e-Neuroforum 2017, 23, 131–136. [Google Scholar] [CrossRef]
- Prato, V.; Taberner, F.J.; Hockley, J.R.F.; Callejo, G.; Arcourt, A.; Tazir, B.; Hammer, L.; Schad, P.; Heppenstall, P.A.; Smith, E.S.; et al. Functional and Molecular Characterization of Mechanoinsensitive “Silent” Nociceptors. Cell Rep. 2017, 21, 3102–3115. [Google Scholar] [CrossRef]
- McGlone, F.; Wessberg, J.; Olausson, H. Discriminative and Affective Touch: Sensing and Feeling. Neuron 2014, 82, 737–755. [Google Scholar] [CrossRef]
- Hucho, T.; Levine, J.D. Signaling Pathways in Sensitization: Toward a Nociceptor Cell Biology. Neuron 2007, 55, 365–376. [Google Scholar] [CrossRef]
- Du, X.; Hao, H.; Yang, Y.; Huang, S.; Wang, C.; Gigout, S.; Ramli, R.; Li, X.; Jaworska, E.; Edwards, I.; et al. Local GABAergic Signaling within Sensory Ganglia Controls Peripheral Nociceptive Transmission. J. Clin. Investig. 2017, 127, 1741–1756. [Google Scholar] [CrossRef]
- Hao, H.; Ramli, R.; Wang, C.; Liu, C.; Shah, S.; Mullen, P.; Lall, V.; Jones, F.; Shao, J.; Zhang, H.; et al. Dorsal Root Ganglia Control Nociceptive Input to the Central Nervous System. PLoS Biol. 2023, 21, e3001958. [Google Scholar] [CrossRef]
- Spigelman, I. Therapeutic Targeting of Peripheral Cannabinoid Receptors in Inflammatory and Neuropathic Pain States. In Translational Pain Research: From Mouse to Man; Kruger, L., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2010. [Google Scholar]
- Sehgal, N.; Smith, H.S.; Manchikanti, L. Peripherally Acting Opioids and Clinical Implications for Pain Control. Pain Physician 2011, 14, 249–258. [Google Scholar] [CrossRef]
- Agarwal, N.; Pacher, P.; Tegeder, I.; Amaya, F.; Constantin, C.E.; Brenner, G.J.; Rubino, T.; Michalski, C.W.; Marsicano, G.; Monory, K.; et al. Cannabinoids Mediate Analgesia Largely Via Peripheral Type 1 Cannabinoid Receptors in Nociceptors. Nat. Neurosci. 2007, 10, 870–879. [Google Scholar] [CrossRef] [PubMed]
- Guindon, J.; Hohmann, A.G. Cannabinoid CB2 Receptors: A Therapeutic Target for the Treatment of Inflammatory and Neuropathic Pain. Br. J. Pharmacol. 2008, 153, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Baron, R.; Hans, G.; Dickenson, A.H. Peripheral Input and its Importance for Central Sensitization. Ann. Neurol. 2013, 74, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wu, Z.; Hadden, J.K.; Odem, M.A.; Zuo, Y.; Crook, R.J.; Frost, J.A.; Walters, E.T. Persistent Pain After Spinal Cord Injury is Maintained by Primary Afferent Activity. J. Neurosci. 2014, 34, 10765–10769. [Google Scholar] [CrossRef]
- North, R.Y.; Odem, M.A.; Li, Y.; Tatsui, C.E.; Cassidy, R.M.; Dougherty, P.M.; Walters, E.T. Electrophysiological Alterations Driving Pain-Associated Spontaneous Activity in Human Sensory Neuron Somata Parallel Alterations Described in Spontaneously Active Rodent Nociceptors. J. Pain. 2022, 23, 1343–1357. [Google Scholar] [CrossRef]
- Velasco, E.; Alvarez, J.L.; Meseguer, V.M.; Gallar, J.; Talavera, K. Membrane Potential Instabilities in Sensory Neurons: Mechanisms and Pathophysiological Relevance. Pain 2022, 163, 64–74. [Google Scholar] [CrossRef]
- Velasco, E.; Alvarez, J.; Meseguer Vigueras, V.M.; Gallar, J.; Talavera, K. Action Potential Firing and Sensory Transduction are Sustained by Membrane Potential Instabilities in Peripheral Sensory Neurons. Biophys. J. 2023, 122, 417a. [Google Scholar] [CrossRef]
- Tian, J.; Bavencoffe, A.G.; Zhu, M.X.; Walters, E.T. Readiness of nociceptor cell bodies to generate spontaneous activity results from background activity of diverse ion channels and high input resistance. Pain 2023. [Google Scholar] [CrossRef]
- van der Vlist, M.; Raoof, R.; Willemen, H.L.D.M.; Prado, J.; Versteeg, S.; Martin Gil, C.; Vos, M.; Lokhorst, R.E.; Pasterkamp, R.J.; Kojima, T.; et al. Macrophages Transfer Mitochondria to Sensory Neurons to Resolve Inflammatory Pain. Neuron 2022, 110, 613–626.e9. [Google Scholar] [CrossRef]
- Bavencoffe, A.G.; Spence, E.A.; Zhu, M.Y.; Garza-Carbajal, A.; Chu, K.E.; Bloom, O.E.; Dessauer, C.W.; Walters, E.T. Macrophage Migration Inhibitory Factor (MIF) Makes Complex Contributions to Pain-Related Hyperactivity of Nociceptors after Spinal Cord Injury. J. Neurosci. 2022, 42, 5463–5480. [Google Scholar] [CrossRef]
- Hanani, M.; Spray, D.C. Emerging Importance of Satellite Glia in Nervous System Function and Dysfunction. Nat. Rev. Neurosci. 2020, 21, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Lim, J.Y.; Seong, J.Y.; Hwang, S.W. The Role of Corticotropin-Releasing Hormone at Peripheral Nociceptors: Implications for Pain Modulation. Biomedicines 2020, 8, 623. [Google Scholar] [CrossRef]
- D’Mello, R.; Dickenson, A.H. Spinal Cord Mechanisms of Pain. Br. J. Anaesth. 2008, 101, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Heinricher, M.M.; Tavares, I.; Leith, J.L.; Lumb, B.M. Descending Control of Nociception: Specificity, Recruitment and Plasticity. Brain Res. Rev. 2009, 60, 214–225. [Google Scholar] [CrossRef]
- Rexed, B. The Cytoarchitectonic Organization of the Spinal Cord in the Cat. J. Comp. Neurol. 1952, 96, 414–495. [Google Scholar] [CrossRef]
- Todd, A. Neuronal Circuitry for Pain Processing in the Dorsal Horn. Nat. Rev. Neurosci. 2010, 11, 823–836. [Google Scholar] [CrossRef] [PubMed]
- Sandkühler, J. Models and Mechanisms of Hyperalgesia and Allodynia. Physiol. Rev. 2009, 89, 707–758. [Google Scholar] [CrossRef]
- Woolf, C.J. Evidence for a Central Component of Post-Injury Pain Hypersensitivity. Nature 1983, 306, 686–688. [Google Scholar] [CrossRef]
- Bliss, T.V.; Lomo, T. Long-lasting Potentiation of Synaptic Transmission in the Dentate Area of the Anaesthetized Rabbit Following Stimulation of the Perforant Path. J. Physiol. 1973, 232, 331–356. [Google Scholar] [CrossRef]
- Randic, M.; Jiang, M.C.; Cerne, R. Long-term Potentiation and Long-term Depression of Primary Afferent Neurotransmission in the Rat Spinal Cord. J. Neurosci. 1993, 13, 5228–5241. [Google Scholar] [CrossRef]
- Ikeda, H.; Stark, J.; Fischer, H.; Wagner, M.; Drdla, R.; Jager, T.; Sandkuhler, J. Synaptic Amplifier of Inflammatory Pain in the Spinal Dorsal Horn. Science 2006, 312, 1659–1662. [Google Scholar] [CrossRef] [PubMed]
- Woolf, C.J. Central Sensitization: Implications for the Diagnosis and Treatment of Pain. Pain 2011, 152, S2–S15. [Google Scholar] [CrossRef] [PubMed]
- Yunus, M.B. Fibromyalgia and Overlapping Disorders: The Unifying Concept of Central Sensitivity Syndromes. Semin. Arthritis Rheum. 2007, 36, 339–356. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Laird, P.N. Mental Models and Deduction. Trends Cogn. Sci. 2001, 5, 434–442. [Google Scholar] [CrossRef]
- Adams, G.R.; Gandhi, W.; Harrison, R.; van Reekum, C.M.; Wood-Anderson, D.; Gilron, I.; Salomons, T.V. Do “Central Sensitization” Questionnaires Reflect Measures of Nociceptive Sensitization or Psychological Constructs? A Systematic Review and Meta-Analyses. Pain 2023, 164, 1222–1239. [Google Scholar] [CrossRef]
- Walitt, B.; Ceko, M.; Gracely, J.L.; Gracely, R.H. Neuroimaging of Central Sensitivity Syndromes: Key Insights from the Scientific Literature. Curr. Rheumatol. Rev. 2016, 12, 55–87. [Google Scholar] [CrossRef]
- Yunus, M.B. Central Sensitivity Syndromes: A New Paradigm and Group Nosology for Fibromyalgia and Overlapping Conditions, and the Related Issue of Disease Versus Illness. Semin. Arthritis Rheum. 2008, 37, 339–352. [Google Scholar] [CrossRef]
- Clauw, D.J. Fibromyalgia and Related Conditions. Mayo Clin. Proc. 2015, 90, 680–692. [Google Scholar] [CrossRef]
- Weisman, A.; Quintner, J.; Masharawi, Y. Amplified Pain Syndrome—An Insupportable Assumption. JAMA Pediatr. 2021, 175, 557–558. [Google Scholar] [CrossRef]
- Al-Chalabi, M.; Reddy, V.; Gupta, S. Neuroanatomy, Spinothalamic Tract. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Bowsher, D. Termination of the Central Pain Pathway in Man: The Conscious Appreciation of Pain. Brain 1957, 80, 607–622. [Google Scholar] [CrossRef]
- Albe-Fessard, D.; Berkley, K.J.; Kruger, L.; Ralston, H.J., III; Willis, W.D., Jr. Diencephalic Mechanisms of Pain Sensation. Brain Res. 1985, 356, 217–296. [Google Scholar] [CrossRef] [PubMed]
- De Ridder, D.; Adhia, D.; Vanneste, S. The Anatomy of Pain and Suffering in the Brain and Its Clinical Implications. Neurosci. Biobehav. Rev. 2021, 130, 125–146. [Google Scholar] [CrossRef]
- Craig, A.D.; Dostrovsky, J. Medulla to Thalamus. In Textbook of Pain; Wall, P., Melzack, R., Eds.; Churchill-Livingstone: Edinburgh, UK, 1999; pp. 183–214. [Google Scholar]
- Tolomeo, S.; Christmas, D.; Jentzsch, I.; Johnston, B.; Sprengelmeyer, R.; Matthews, K.; Steele, J.D. A Causal Role for the Anterior Mid-Cingulate Cortex in Negative Affect and Cognitive Control. Brain 2016, 139, 1844–1854. [Google Scholar] [CrossRef] [PubMed]
- Hohwy, J.; Seth, A.K. Predictive Processing as a Systematic Basis for Identifying the Neural Correlates of Consciousness. Philos. Mind Sci. 2020, 1, 3. [Google Scholar] [CrossRef]
- Flor, H.; Elbert, T.; Knecht, S.; Wienbruch, C.; Pantev, C.; Birbaumers, N.; Larbig, W.; Taub, E. Phantom-Limb Pain as a Perceptual Correlate of Cortical Reorganization Following Arm Amputation. Nature 1995, 375, 482–484. [Google Scholar] [CrossRef] [PubMed]
- Gerrans, P. Pain Asymbolia as Depersonalization for Pain Experience. An Interoceptive Active Inference Account. Front. Psychol. 2020, 11, 523710. [Google Scholar] [CrossRef]
- Grahek, N. Feeling Pain and Being in Pain, 2nd ed.; Boston Review; Bradford Books; MIT Pess: Cambridge, MA, USA, 2007. [Google Scholar]
- Stilwell, P.; Hudon, A.; Meldrum, K.; Pagé, M.G.; Wideman, T.H. What is Pain-Related Suffering? Conceptual Critiques, Key Attributes, and Outstanding Questions. J. Pain 2022, 23, 729–738. [Google Scholar] [CrossRef]
- Koolhaas, J.M.; Bartolomucci, A.; Buwalda, B.; de Boer, S.; Flügge, G.; Korte, S.; Meerlo, P.; Murison, R.; Olivier, B.; Palanza, P.; et al. Stress Revisited: A Critical Evaluation of the Stress Concept. Neurosci. Biobehav. Rev. 2011, 35, 1291–1301. [Google Scholar] [CrossRef]
- Blackburn-Munro, G.; Blackburn-Munro, R.E. Chronic Pain, Chronic Stress and Depression: Coincidence or Consequence? J. Neuroendocrinol. 2001, 13, 1009–1023. [Google Scholar] [CrossRef]
- Abdallah, C.G.; Geha, P. Chronic Pain and Chronic Stress: Two Sides of the Same Coin? Chronic Stress 2017, 1, 2470547017704763. [Google Scholar] [CrossRef]
- Kornelsen, J.; McIver, T.A.; Stroman, P.W. Unique Brain Regions Involved in Positive Versus Negative Emotional Modulation of Pain. Scand. J. Pain 2019, 19, 583–596. [Google Scholar] [CrossRef]
- Mathur, V.A.; Moayedi, M.; Keaser, M.L.; Khan, S.A.; Hubbard, C.S.; Goyal, M.; Seminowicz, D.A. High Frequency Migraine is Associated with Lower Acute Pain Sensitivity and Abnormal Insula Activity Related to Migraine Pain Intensity, Attack Frequency, Pain Catastrophizing. Front. Hum. Neurosci. 2016, 10, 489. [Google Scholar] [CrossRef]
- Yang, S.; Chang, M.C. Chronic Pain: Structural and Functional Changes in Brain Structures and Associated Negative Affective States. Int. J. Mol. Sci. 2019, 20, 3130. [Google Scholar] [CrossRef]
- Wade, J.B.; Riddle, D.L.; Price, D.D.; Dumenci, L. Role of Pain Catastrophizing During Pain Processing in a Cohort of Patients with Chronic and Severe Arthritic Knee Pain. Pain 2011, 152, 314–319. [Google Scholar] [CrossRef]
- Edwards, R.R.; Dworkin, R.H.; Sullivan, M.D.; Turk, D.C.; Wasan, A.D. The Role of Psychosocial Processes in the Development and Maintenance of Chronic Pain. J. Pain 2016, 17, T70–T92. [Google Scholar] [CrossRef]
- Piazza, M. Maine de Biran and Gall’s Phrenology: The Origins of a Debate About the Localization of Mental Faculties. Br. J. Hist. Philos. 2020, 28, 866–884. [Google Scholar] [CrossRef]
- Iannetti, G.D.; Mouraux, A. From the Neuromatrix to the Pain Matrix (and Back). Exp. Brain Res. 2010, 205, 1–12. [Google Scholar] [CrossRef]
- Melzack, R. From the Gate to the Neuromatrix. Pain 1999, 82, S121–S126. [Google Scholar] [CrossRef]
- Tracey, I.; Mantyh, P.W. The Cerebral Signature for Pain Perception and Its Modulation. Neuron 2007, 55, 377–391. [Google Scholar] [CrossRef]
- Büchel, C.; Bornhovd, K.; Quante, M.; Glauche, V.; Bromm, B.; Weiller, C. Dissociable Neural Responses Related to Pain Intensity, Stimulus Intensity, and Stimulus Awareness within the Anterior Cingulate Cortex: A Parametric Single-Trial Laser Functional Magnetic Resonance Imaging Study. J. Neurosci. 2002, 22, 970–976. [Google Scholar] [CrossRef]
- Hofbauer, R.K.; Rainville, P.; Duncan, G.H.; Bushnell, M.C. Cortical Representation of the Sensory Dimension of Pain. J. Neurophysiol. 2001, 86, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Schweinhardt, P.; Bountra, C.; Tracey, I. Pharmacological FMRI in the Development of New Analgesic Compounds. NMR Biomed. 2006, 19, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Boly, M.; Faymonville, M.E.; Schnakers, C.; Peigneux, P.; Lambermont, B.; Phillips, C.; Lancellotti, P.; Luxen, A.; Lamy, M.; Moonen, G.; et al. Perception of Pain in the Minimally Conscious State with PET Activation: An Observational Study. Lancet Neurol. 2008, 7, 1013–1020. [Google Scholar] [CrossRef] [PubMed]
- Legrain, V.; Iannetti, G.D.; Plaghki, L.; Mouraux, A. The Pain Matrix Reloaded: A Salience Detection System for the Body. Prog. Neurobiol. 2011, 93, 111–124. [Google Scholar] [CrossRef]
- Coninx, S. The Notorious Neurophilosophy of Pain: A Family Resemblance Approach to Idiosyncrasy and Generalizability. Mind Lang. 2023, 38, 178–197. [Google Scholar] [CrossRef]
- Wall, P.D. Independent Mechanisms Converge on Pain. Nat. Med. 1995, 1, 740–741. [Google Scholar] [CrossRef]
- Choi, J.M.; Padmala, S.; Spechler, P.; Pessoa, L. Pervasive Competition between Threat and Reward in the Brain. Soc. Cogn. Affect. Neurosci. 2014, 9, 737–750. [Google Scholar] [CrossRef]
- Elman, I.; Borsook, D. Common Brain Mechanisms of Chronic Pain and Addiction. Neuron 2016, 89, 11–36. [Google Scholar] [CrossRef]
- De Ridder, D.; Vanneste, S. The bayesian brain in imbalance: Medial, lateral and descending pathways in tinnitus and pain: A perspective. Prog. Brain Res. 2021, 262, 309–334. [Google Scholar] [CrossRef]
- Tabor, A.; Thacker, M.A.; Moseley, G.L.; Körding, K.P. Pain: A Statistical Account. PLoS Comput. Biol. 2017, 13, e1005142. [Google Scholar] [CrossRef]
- Harris, S.; Sheth, S.A.; Cohen, M.S. Functional Neuroimaging of Belief, Disbelief, and Uncertainty. Ann. Neurol. 2008, 63, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Eckert, A.L.; Pabst, K.; Endres, D.M. A Bayesian Model for Chronic Pain. Front. Pain Res. 2022, 3, 966034. [Google Scholar] [CrossRef] [PubMed]
- Hechler, T.; Endres, D.M.; Thorwart, A. Why Harmless Sensations Might Hurt in Individuals with Chronic Pain: About Heightened Prediction and Perception of Pain in the Mind. Front. Psychol. 2016, 7, 1638. [Google Scholar] [CrossRef] [PubMed]
- McCarberg, B.; Peppin, J. Pain Pathways and Nervous System Plasticity: Learning and Memory in Pain. Pain Med. 2019, 20, 2421–2437. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, X.; Bi, Y.; Hu, L. Pavlov’s Pain: The Effect of Classical Conditioning on Pain Perception and its Clinical Implications. Curr. Pain Headache Rep. 2019, 23, 19. [Google Scholar] [CrossRef]
- Gatzounis, R.; Schrooten, M.G.; Crombez, G.; Vlaeyen, J.W. Operant learning theory in pain and chronic pain rehabilitation. Curr. Pain Headache Rep. 2012, 16, 117–126. [Google Scholar] [CrossRef]
- Stegemann, A.; Liu, S.; Romero, O.A.R.; Oswald, M.J.; Han, Y.; Beretta, C.A.; Gan, Z.; Tan, L.L.; Wisden, W.; Gräff, J.; et al. Prefrontal engrams of long-term fear memory perpetuate pain perception. Nat. Neurosci. 2023, 26, 820–829. [Google Scholar] [CrossRef]
- Blasini, M.; Corsi, N.; Klinger, R.; Colloca, L. Nocebo and Pain: An Overview of the Psychoneurobiological Mechanisms. Pain Rep. 2017, 2, e585. [Google Scholar] [CrossRef]
- Rossettini, G.; Campaci, F.; Bialosky, J.; Huysmans, E.; Vase, L.; Carlino, E. The Biology of Placebo and Nocebo Effects on Experimental and Chronic Pain: State of the Art. J. Clin. Med. 2023, 12, 4113. [Google Scholar] [CrossRef]
- Colloca, L.; Barsky, A.J. Placebo and Nocebo Effects. N. Engl. J. Med. 2020, 382, 554–561. [Google Scholar] [CrossRef]
- Clark, A. Whatever next? Predictive brains, situated agents, and the future of cognitive science. Behav. Brain Sci. 2013, 36, 181–204. [Google Scholar] [CrossRef]
- Seth, A.K.; Bayne, T. Theories of consciousness. Nat. Rev. Neurosci. 2022, 23, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.; Lau, H.; LeDoux, J.E. Understanding the higher-order approach to consciousness. Trends Cogn. Sci. 2019, 23, 754–768. [Google Scholar] [CrossRef]
- Mashour, G.A.; Roelfsema, P.; Changeux, J.P.; Dehaene, S. Conscious processing and the global neuronal workspace hypothesis. Neuron 2020, 105, 776–798. [Google Scholar] [CrossRef]
- Tononi, G.; Boly, M.; Massimini, M.; Koch, C. Integrated information theory: From consciousness to its physical substrate. Nat. Rev. Neurosci. 2016, 17, 450–461. [Google Scholar] [CrossRef]
- Lamme, V.A. Towards a true neural stance on consciousness. Trends Cogn. Sci. 2006, 10, 494–501. [Google Scholar] [CrossRef]
- Piasini, E.; Panzeri, S. Information Theory in Neuroscience. Entropy 2019, 21, 62. [Google Scholar] [CrossRef]
- Borst, A.; Theunissen, F.E. Information theory and neural coding. Nat. Neurosci. 1999, 2, 947–957. [Google Scholar] [CrossRef]
- Shannon, C.E. The mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Eckhorn, R.; Popel, B. Rigorous and extended application of information theory to the afferent visual system of the cat. Biol. Cybern. 1975, 17, 7–17. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Párraga, J.P.; Castellanos, A. A Manifesto in Defense of Pain Complexity: A Critical Review of Essential Insights in Pain Neuroscience. J. Clin. Med. 2023, 12, 7080. https://doi.org/10.3390/jcm12227080
Párraga JP, Castellanos A. A Manifesto in Defense of Pain Complexity: A Critical Review of Essential Insights in Pain Neuroscience. Journal of Clinical Medicine. 2023; 12(22):7080. https://doi.org/10.3390/jcm12227080
Chicago/Turabian StylePárraga, Javier Picañol, and Aida Castellanos. 2023. "A Manifesto in Defense of Pain Complexity: A Critical Review of Essential Insights in Pain Neuroscience" Journal of Clinical Medicine 12, no. 22: 7080. https://doi.org/10.3390/jcm12227080