
Aminosteroid RM-581 Decreases Cell Proliferation of All Breast Cancer Molecular Subtypes, Alone and in Combination with Breast Cancer Treatments

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Proliferation and Drug Combination Assays
2.3. Spheroids Assay (3D Culture)
2.4. RNA Isolation and Quantitative Real-Time PCR (qPCR)
3. Results
3.1. RM-581 Has an Antiproliferative Effect on Breast Cancer Cell Lines
3.1.1. RM-581 Antiproliferative Effect Found in 2D Culture
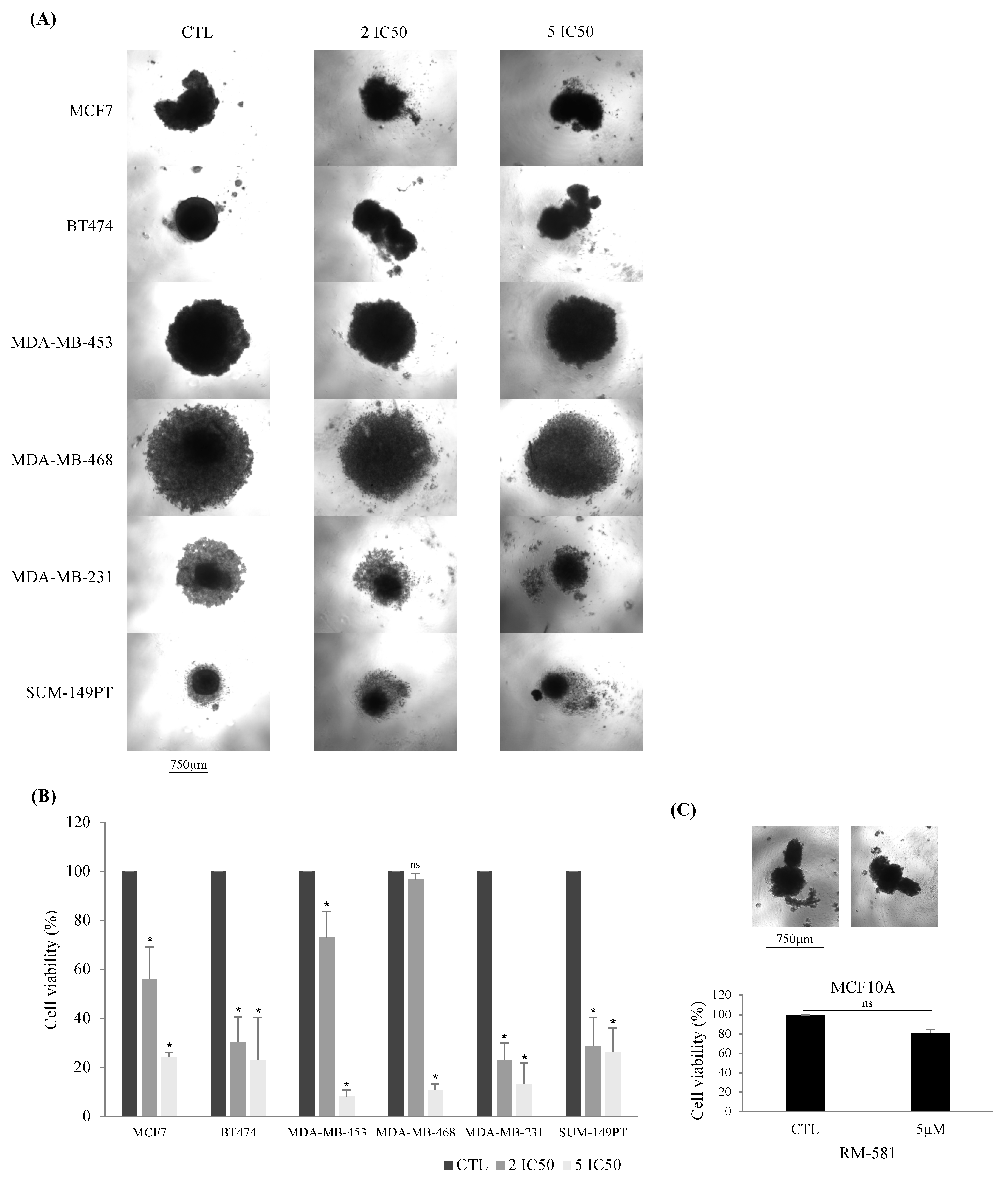
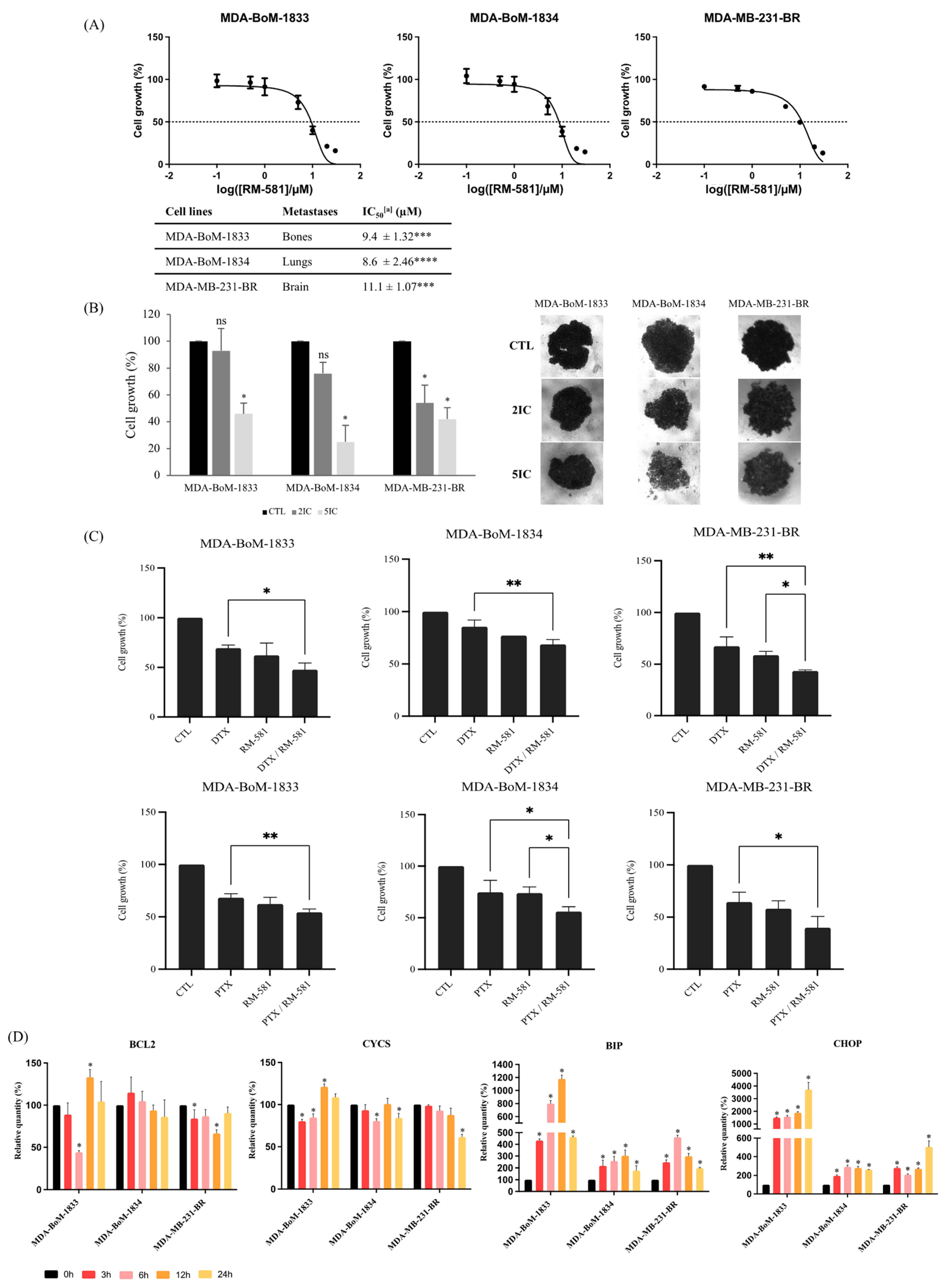
3.1.2. RM-581 Antiproliferative Effect Found in 3D Culture
3.2. RM-581 Is More Effective in Combination with Other Breast Cancer Treatments
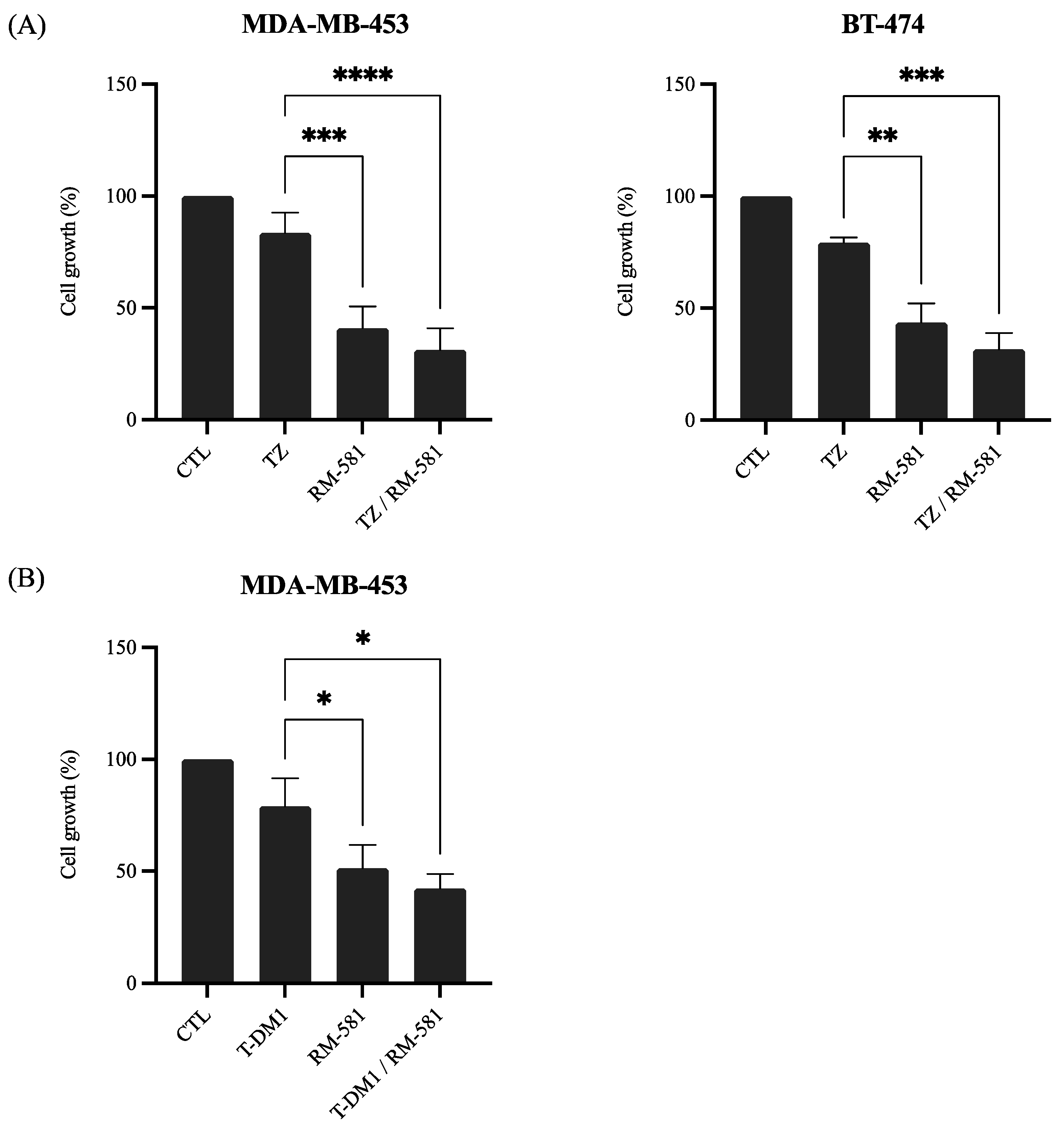
3.2.1. RM-581 in Combination with Anti-HER2 Therapies
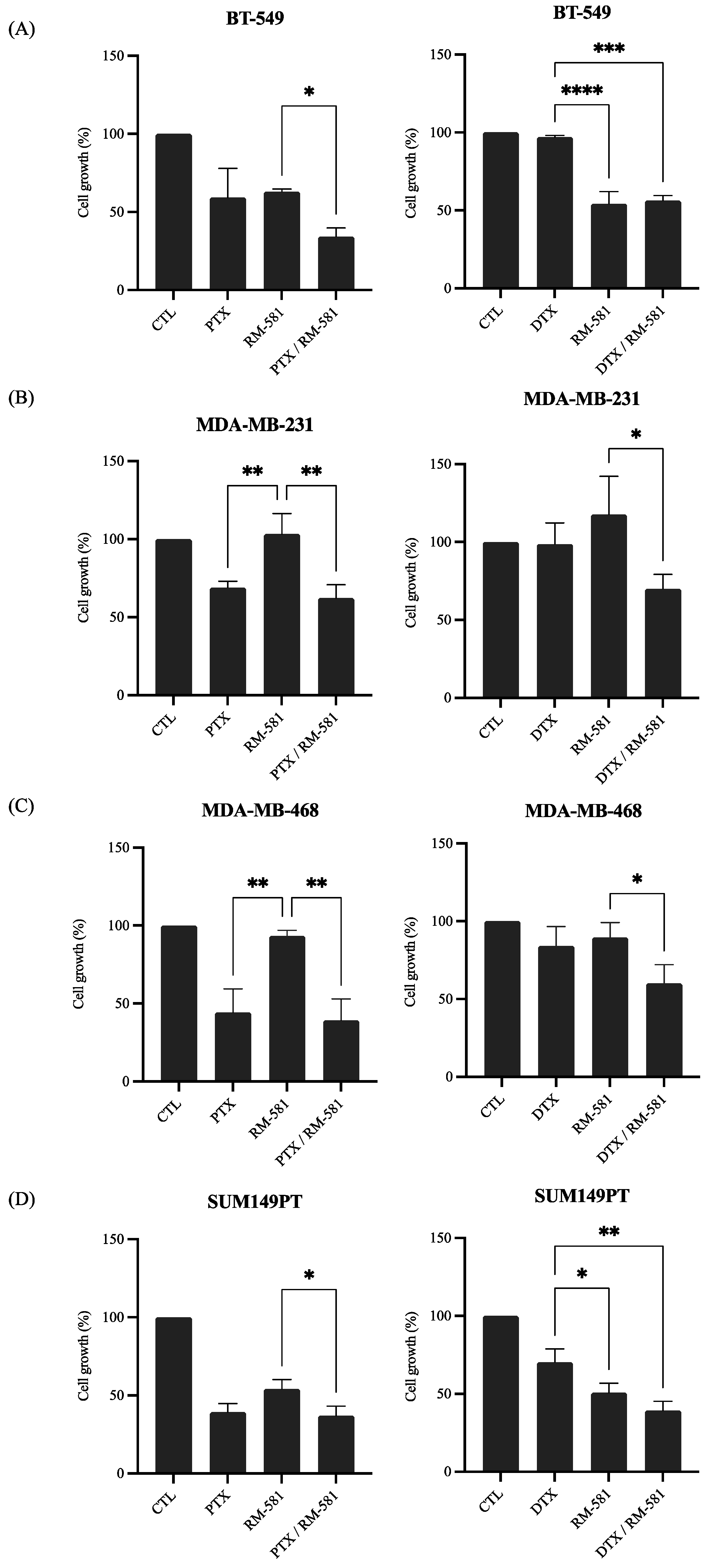
3.2.2. RM-581 in Combination with Chemotherapy
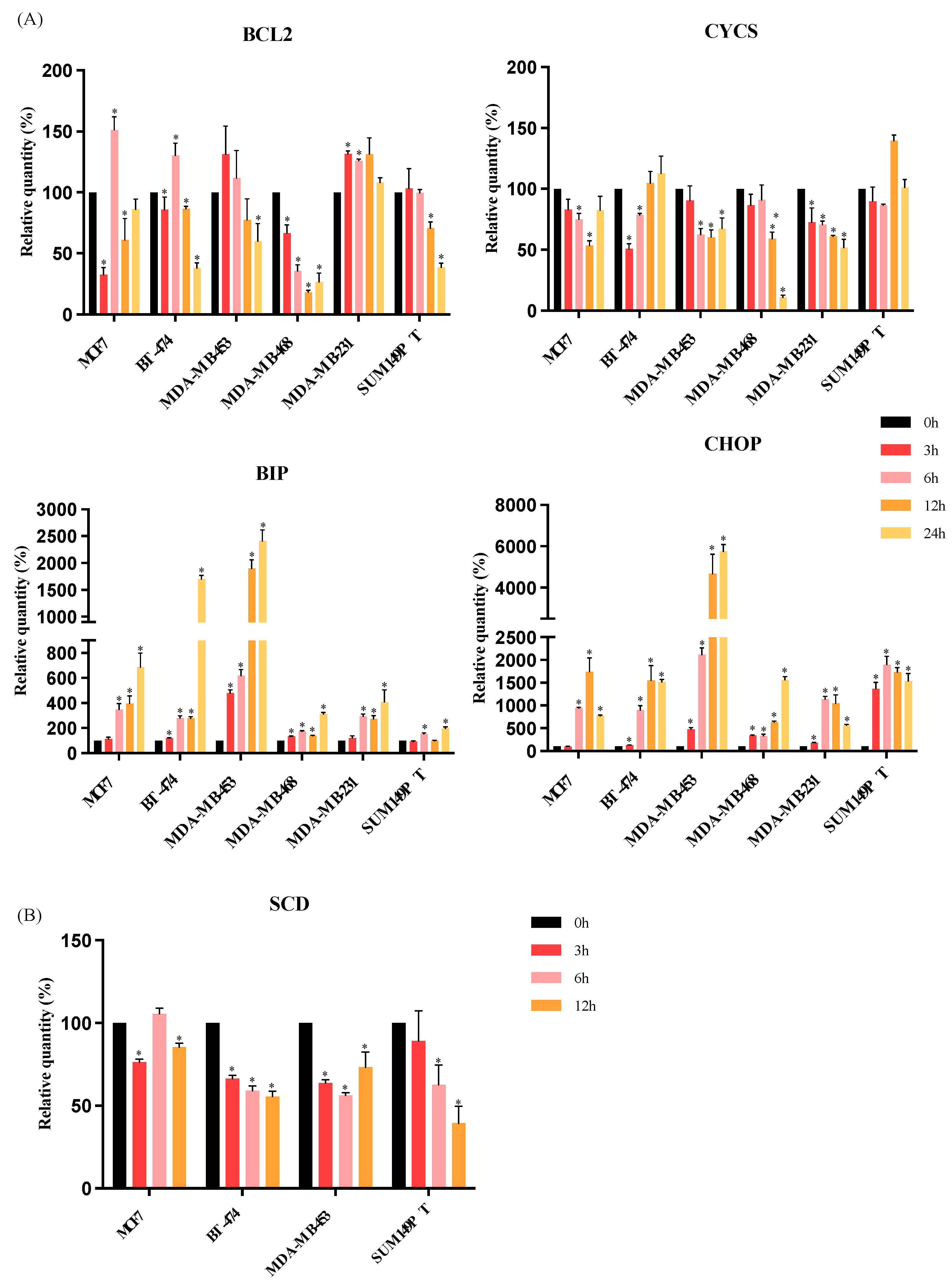
3.3. RM-581 Increases the Expression of Endoplasmic Reticulum Stress Apoptosis Markers
3.4. RM-581 Is Effective against TNBC Derivative Metastasis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Harbeck, N.; Gnant, M. Breast Cancer. Lancet 2017, 389, 1134–1150. [Google Scholar] [CrossRef] [PubMed]
- Howlader, N.; Cronin, K.A.; Kurian, A.W.; Andridge, R. Differences in Breast Cancer Survival by Molecular Subtypes in the United States. Cancer Epidemiol. Biomark. Prev. 2018, 27, 619–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchini, G.; Balko, J.M.; Mayer, I.A.; Sanders, M.E.; Gianni, L. Triple-Negative Breast Cancer: Challenges and Opportunities of a Heterogeneous Disease. Nat. Rev. Clin. Oncol. 2016, 13, 674–690. [Google Scholar] [CrossRef]
- Maltais, R.; Hospital, A.; Delhomme, A.; Roy, J.; Poirier, D. Chemical Synthesis, NMR Analysis and Evaluation on a Cancer Xenograft Model (HL-60) of the Aminosteroid Derivative RM-133. Steroids 2014, 82, 68–76. [Google Scholar] [CrossRef]
- Talbot, A.; Maltais, R.; Poirier, D. New Diethylsilylacetylenic Linker for Parallel Solid-Phase Synthesis of Libraries of Hydroxy Acetylenic Steroid Derivatives with Improved Metabolic Stability. ACS Comb. Sci. 2012, 14, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Jegham, H.; Maltais, R.; Roy, J.; Doillon, C.; Poirier, D. Biological Evaluation of a New Family of Aminosteroids That Display a Selective Toxicity for Various Malignant Cell Lines. Anticancer Drugs 2012, 23, 803–814. [Google Scholar] [CrossRef]
- Kenmogne, L.C.; Ayan, D.; Roy, J.; Maltais, R.; Poirier, D. The Aminosteroid Derivative RM-133 Shows In Vitro and In Vivo Antitumor Activity in Human Ovarian and Pancreatic Cancers. PLoS ONE 2015, 10, e0144890. [Google Scholar] [CrossRef] [Green Version]
- Maltais, R.; Perreault, M.; Roy, J.; Poirier, D. Minor Chemical Modifications of the Aminosteroid Derivative RM-581 Lead to Major Impact on Its Anticancer Activity, Metabolic Stability and Aqueous Solubility. Eur. J. Med. Chem. 2020, 188, 111990. [Google Scholar] [CrossRef]
- Perreault, M.; Maltais, R.; Roy, J.; Dutour, R.; Poirier, D. Design of a Mestranol 2-N-Piperazino-Substituted Derivative Showing Potent and Selective in Vitro and in Vivo Activities in MCF-7 Breast Cancer Models. ChemMedChem 2017, 12, 177–182. [Google Scholar] [CrossRef]
- Perreault, M.; Maltais, R.; Roy, J.; Picard, S.; Popa, I.; Bertrand, N.; Poirier, D. Induction of Endoplasmic Reticulum Stress by Aminosteroid Derivative RM-581 Leads to Tumor Regression in PANC-1 Xenograft Model. Investig. New Drugs 2019, 37, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.; Maltais, R.; Jegham, H.; Poirier, D. Libraries of 2β-(N-Substituted Piperazino)-5α-Androstane-3α, 17β-Diols: Chemical Synthesis and Cytotoxic Effects on Human Leukemia HL-60 Cells and on Normal Lymphocytes. Mol. Divers. 2011, 15, 317–339. [Google Scholar] [CrossRef] [PubMed]
- Ayan, D.; Maltais, R.; Hospital, A.; Poirier, D. Chemical Synthesis, Cytotoxicity, Selectivity and Bioavailability of 5α-Androstane-3α,17β-Diol Derivatives. Bioorg. Med. Chem. 2014, 22, 5847–5859. [Google Scholar] [CrossRef] [PubMed]
- Maltais, R.; Roy, J.; Perreault, M.; Sato, S.; Lévesque, J.-C.; Poirier, D. Induction of Endoplasmic Reticulum Stress-Mediated Apoptosis by Aminosteroid RM-581 Efficiently Blocks the Growth of PC-3 Cancer Cells and Tumors Resistant or Not to Docetaxel. Int. J. Mol. Sci. 2021, 22, 11181. [Google Scholar] [CrossRef]
- Schröder, M.; Kaufman, R.J. The Mammalian Unfolded Protein Response. Annu. Rev. Biochem. 2005, 74, 739–789. [Google Scholar] [CrossRef]
- Riha, R.; Gupta-Saraf, P.; Bhanja, P.; Badkul, S.; Saha, S. Stressed Out—Therapeutic Implications of ER Stress Related Cancer Research. Oncomedicine 2017, 2, 156–167. [Google Scholar] [CrossRef] [Green Version]
- Cragg, G.M.; Grothaus, P.G.; Newman, D.J. Impact of Natural Products on Developing New Anti-Cancer Agents. Chem. Rev. 2009, 109, 3012–3043. [Google Scholar] [CrossRef]
- Kelloff, G.J.; Crowell, J.A.; Steele, V.E.; Lubet, R.A.; Malone, W.A.; Boone, C.W.; Kopelovich, L.; Hawk, E.T.; Lieberman, R.; Lawrence, J.A.; et al. Progress in Cancer Chemoprevention: Development of Diet-Derived Chemopreventive Agents. J. Nutr. 2000, 130, 467S–471S. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Song, H.-S.; Park, H.; Kim, B. Activation of ER Stress-Dependent MiR-216b Has a Critical Role in Salvia Miltiorrhiza Ethanol-Extract-Induced Apoptosis in U266 and U937 Cells. Int. J. Mol. Sci. 2018, 19, 1240. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.; Song, H.-S.; Kang, B.; Park, M.; Park, K.; Kim, S.-H.; Shim, B.-S.; Kim, B. MiR-211 Plays a Critical Role in Cnidium Officinale Makino Extract-Induced, ROS/ER Stress-Mediated Apoptosis in U937 and U266 Cells. Int. J. Mol. Sci. 2018, 19, 865. [Google Scholar] [CrossRef] [Green Version]
- Clarke, R.; Cook, K.L.; Hu, R.; Facey, C.O.B.; Tavassoly, I.; Schwartz, J.L.; Baumann, W.T.; Tyson, J.J.; Xuan, J.; Wang, Y.; et al. Endoplasmic Reticulum Stress, the Unfolded Protein Response, Autophagy, and the Integrated Regulation of Breast Cancer Cell Fate. Cancer Res. 2012, 72, 1321–1331. [Google Scholar] [CrossRef] [Green Version]
- Ko, E.-Y.; Moon, A. Natural Products for Chemoprevention of Breast Cancer. J. Cancer Prev. 2015, 20, 223–231. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, T.; Nishihara, H.; Hara, H.; Adachi, T. Ethanol Extract of Brazilian Red Propolis Induces Apoptosis in Human Breast Cancer MCF-7 Cells through Endoplasmic Reticulum Stress. J. Agric. Food Chem. 2012, 60, 11065–11070. [Google Scholar] [CrossRef]
- Shi, J.-M.; Bai, L.-L.; Zhang, D.-M.; Yiu, A.; Yin, Z.-Q.; Han, W.-L.; Liu, J.-S.; Li, Y.; Fu, D.-Y.; Ye, W.-C. Saxifragifolin D Induces the Interplay between Apoptosis and Autophagy in Breast Cancer Cells through ROS-Dependent Endoplasmic Reticulum Stress. Biochem. Pharmacol. 2013, 85, 913–926. [Google Scholar] [CrossRef] [PubMed]
- Burguin, A.; Furrer, D.; Ouellette, G.; Jacob, S.; Diorio, C.; Durocher, F. Trastuzumab Effects Depend on HER2 Phosphorylation in HER2-Negative Breast Cancer Cell Lines. PLoS ONE 2020, 15, e0234991. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Paik, S.; Kim, C.; Wolmark, N. HER2 Status and Benefit from Adjuvant Trastuzumab in Breast Cancer. N. Engl. J. Med. 2008, 358, 1409–1411. [Google Scholar] [CrossRef]
- Lewis Phillips, G.D.; Li, G.; Dugger, D.L.; Crocker, L.M.; Parsons, K.L.; Mai, E.; Blättler, W.A.; Lambert, J.M.; Chari, R.V.J.; Lutz, R.J.; et al. Targeting HER2-Positive Breast Cancer with Trastuzumab-DM1, an Antibody-Cytotoxic Drug Conjugate. Cancer Res. 2008, 68, 9280–9290. [Google Scholar] [CrossRef] [Green Version]
- Burguin, A.; Diorio, C.; Durocher, F. Breast Cancer Treatments: Updates and New Challenges. J. Pers. Med. 2021, 11, 808. [Google Scholar] [CrossRef]
- Koeberle, A.; Löser, K.; Thürmer, M. Stearoyl-CoA Desaturase-1 and Adaptive Stress Signaling. Biochim. Biophys. Acta 2016, 1861, 1719–1726. [Google Scholar] [CrossRef]
- Bergin, A.R.T.; Loi, S. Triple-Negative Breast Cancer: Recent Treatment Advances. F1000Research 2019, 8, 1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.; Siegel, P.M.; Shu, W.; Drobnjak, M.; Kakonen, S.M.; Cordón-Cardo, C.; Guise, T.A.; Massagué, J. A Multigenic Program Mediating Breast Cancer Metastasis to Bone. Cancer Cell 2003, 3, 537–549. [Google Scholar] [CrossRef] [Green Version]
- Dun, M.D.; Chalkley, R.J.; Faulkner, S.; Keene, S.; Avery-Kiejda, K.A.; Scott, R.J.; Falkenby, L.G.; Cairns, M.J.; Larsen, M.R.; Bradshaw, R.A.; et al. Proteotranscriptomic Profiling of 231-BR Breast Cancer Cells: Identification of Potential Biomarkers and Therapeutic Targets for Brain Metastasis. Mol. Cell. Proteom. MCP 2015, 14, 2316–2330. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of Human Triple-Negative Breast Cancer Subtypes and Preclinical Models for Selection of Targeted Therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef] [Green Version]
- Fisusi, F.A.; Akala, E.O. Drug Combinations in Breast Cancer Therapy. Pharm. Nanotechnol. 2019, 7, 3–23. [Google Scholar] [CrossRef]
- Wuerstlein, R.; Harbeck, N. Neoadjuvant Therapy for HER2-Positive Breast Cancer. Rev. Recent Clin. Trials 2017, 12, 81–92. [Google Scholar] [CrossRef]
- Minckwitz, G.; Procter, M.; Azambuja, E.; Zardavas, D.; Benyunes, M.; Viale, G.; Suter, T.; Arahmani, A.; Rouchet, N.; Clark, E.; et al. Adjuvant Pertuzumab and Trastuzumab in Early HER2-Positive Breast Cancer. N. Engl. J. Med. 2017, 377, 122–131. [Google Scholar] [CrossRef]
- Amiri-Kordestani, L.; Blumenthal, G.M.; Xu, Q.C.; Zhang, L.; Tang, S.W.; Ha, L.; Weinberg, W.C.; Chi, B.; Candau-Chacon, R.; Hughes, P.; et al. FDA Approval: Ado-Trastuzumab Emtansine for the Treatment of Patients with HER2-Positive Metastatic Breast Cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2014, 20, 4436–4441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahba, H.A.; El-Hadaad, H.A. Current Approaches in Treatment of Triple-Negative Breast Cancer. Cancer Biol. Med. 2015, 12, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Female Breast Cancer Subtypes—Cancer Stat Facts. Available online: https://seer.cancer.gov/statfacts/html/breast-subtypes.html (accessed on 5 May 2022).
- Minn, A.J.; Gupta, G.P.; Siegel, P.M.; Bos, P.D.; Shu, W.; Giri, D.D.; Viale, A.; Olshen, A.B.; Gerald, W.L.; Massagué, J. Genes That Mediate Breast Cancer Metastasis to Lung. Nature 2005, 436, 518–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sisinni, L.; Pietrafesa, M.; Lepore, S.; Maddalena, F.; Condelli, V.; Esposito, F.; Landriscina, M. Endoplasmic Reticulum Stress and Unfolded Protein Response in Breast Cancer: The Balance between Apoptosis and Autophagy and Its Role in Drug Resistance. Int. J. Mol. Sci. 2019, 20, 857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.S. GRP78 Induction in Cancer: Therapeutic and Prognostic Implications. Cancer Res. 2007, 67, 3496–3499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Wey, S.; Zhang, Y.; Ye, R.; Lee, A.S. Role of the Unfolded Protein Response Regulator GRP78/BiP in Development, Cancer, and Neurological Disorders. Antioxid. Redox Signal. 2009, 11, 2307–2316. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.S. The ER Chaperone and Signaling Regulator GRP78/BiP as a Monitor of Endoplasmic Reticulum Stress. Methods 2005, 35, 373–381. [Google Scholar] [CrossRef]
- Marciniak, S.J.; Yun, C.Y.; Oyadomari, S.; Novoa, I.; Zhang, Y.; Jungreis, R.; Nagata, K.; Harding, H.P.; Ron, D. CHOP Induces Death by Promoting Protein Synthesis and Oxidation in the Stressed Endoplasmic Reticulum. Genes Dev. 2004, 18, 3066–3077. [Google Scholar] [CrossRef] [Green Version]
- Maltais, R.; Roy, J.; Poirier, D. Turning a Quinoline-Based Steroidal Anticancer Agent into Fluorescent Dye for Its Tracking by Cell Imaging. ACS Med. Chem. Lett. 2021, 12, 822–826. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.R.; Schulze, A. Lipid Metabolism in Cancer. FEBS J. 2012, 279, 2610–2623. [Google Scholar] [CrossRef] [PubMed]
- Cosentino, G.; Plantamura, I.; Tagliabue, E.; Iorio, M.V.; Cataldo, A. Breast Cancer Drug Resistance: Overcoming the Challenge by Capitalizing on MicroRNA and Tumor Microenvironment Interplay. Cancers 2021, 13, 3691. [Google Scholar] [CrossRef] [PubMed]
- Tanner, M.; Kapanen, A.I.; Junttila, T.; Raheem, O.; Grenman, S.; Elo, J.; Elenius, K.; Isola, J. Characterization of a Novel Cell Line Established from a Patient with Herceptin-Resistant Breast Cancer. Mol. Cancer Ther. 2004, 3, 1585–1592. [Google Scholar] [CrossRef]
- Friedrich, J.; Seidel, C.; Ebner, R.; Kunz-Schughart, L.A. Spheroid-Based Drug Screen: Considerations and Practical Approach. Nat. Protoc. 2009, 4, 309–324. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | Molecular Subtype | IC50 (µM) [a] |
|---|---|---|
| MCF10A | Normal | 17.1 ± 0.61 |
| MCF7 | Luminal A | 2.8 ± 1.15 **** |
| BT-474 | Luminal B | 12.0 ± 4.28 ** |
| MDA-MB-453 | HER2 | 13.4 ± 0.97 |
| JIMT-1 | HER2 | 12.3 ± 2.21 * |
| MDA-MB-231 | TNBC | 10.4 ± 2.40 *** |
| BT-549 | TNBC | 8.7 ± 0.37 **** |
| SUM159PT | TNBC | 8.5 ± 1.10 **** |
| MDA-MB-468 | TNBC | 6.9 ± 1.03 **** |
| SUM149PT | TNBC | 5.6 ± 0.96 **** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burguin, A.; Roy, J.; Ouellette, G.; Maltais, R.; Bherer, J.; Diorio, C.; Poirier, D.; Durocher, F. Aminosteroid RM-581 Decreases Cell Proliferation of All Breast Cancer Molecular Subtypes, Alone and in Combination with Breast Cancer Treatments. J. Clin. Med. 2023, 12, 4241. https://doi.org/10.3390/jcm12134241
Burguin A, Roy J, Ouellette G, Maltais R, Bherer J, Diorio C, Poirier D, Durocher F. Aminosteroid RM-581 Decreases Cell Proliferation of All Breast Cancer Molecular Subtypes, Alone and in Combination with Breast Cancer Treatments. Journal of Clinical Medicine. 2023; 12(13):4241. https://doi.org/10.3390/jcm12134241
Chicago/Turabian StyleBurguin, Anna, Jenny Roy, Geneviève Ouellette, René Maltais, Juliette Bherer, Caroline Diorio, Donald Poirier, and Francine Durocher. 2023. "Aminosteroid RM-581 Decreases Cell Proliferation of All Breast Cancer Molecular Subtypes, Alone and in Combination with Breast Cancer Treatments" Journal of Clinical Medicine 12, no. 13: 4241. https://doi.org/10.3390/jcm12134241