
The Role of Extracellular Matrix Components in Inflammatory Bowel Diseases


Abstract
:1. Introduction
2. The Role of MMPs in Extracellular Matrix Remodeling in the Progression of Inflammatory Bowel Diseases
3. The Role of MMP in the Immunological Response
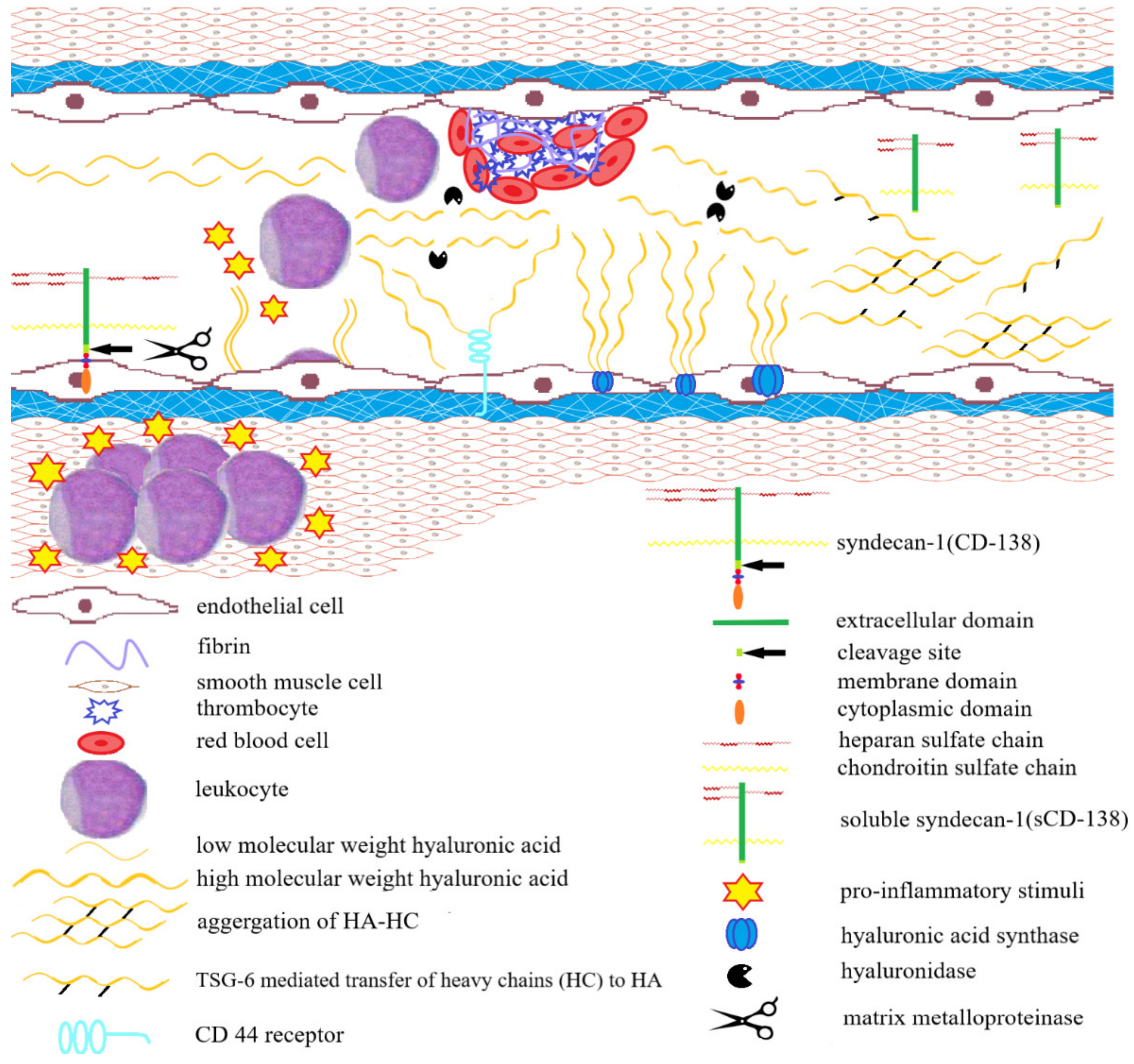
4. The Role of Hyaluronan in the Progression of Inflammatory Bowel Diseases
5. Other Extracellular Matrix Components Involved in the Inflammatory Process during IBD
5.1. Laminin (LN)
5.2. Heparan Sulfate Proteoglycans (HSPG)
5.3. Fibronectin (FN)
5.4. Sulfated GAGs
6. Extracellular Matrix Components as Biomarkers in IBD Diagnosis
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rieder, F.; Fiocchi, C. Intestinal fibrosis in IBD—A dynamic, multifactorial process. Nat. Rev. Gastroenterol. Hepatol. 2009, 6, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Latella, G.; Rogler, G.; Bamias, G.; Breynaert, C.; Florholmen, J.; Pellino, G.; Reif, S.; Speca, S.; Lawrance, I.C. Results of the 4th scientific workshop of the ECCO (I): Pathophysiology of intestinal fibrosis in IBD. J. Crohn’s Colitis 2014, 8, 1147–1165. [Google Scholar] [CrossRef] [Green Version]
- Rieder, F.; de Bruyn, J.R.; Pham, B.T.; Katsanos, K.; Annese, V.; Higgins, P.D.; Magro, F.; Dotan, I. Results of the 4th Scientific Workshop of the ECCO (Group II): Markers of intestinal fibrosis in inflammatory bowel disease. J. Crohn’s Colitis 2014, 8, 1166–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Steen, P.E.V.D.; Sang, Q.-X.A.; Opdenakker, G. Matrix metalloproteinase inhibitors as therapy for inflammatory and vascular diseases. Nat. Rev. Drug Discov. 2007, 6, 480–498. [Google Scholar] [CrossRef]
- Bailey, J.R.; Bland, P.W.; Tarlton, J.F.; Peters, I.; Moorghen, M.; Sylvester, P.A.; Probert, C.S.J.; Whiting, C.V. IL-13 Promotes Collagen Accumulation in Crohn’s Disease Fibrosis by Down-Regulation of Fibroblast MMP Synthesis: A Role for Innate Lymphoid Cells? PLoS ONE 2012, 7, e52332. [Google Scholar] [CrossRef] [Green Version]
- Koelink, P.J.; Overbeek, S.A.; Braber, S.; Morgan, M.E.; Henricks, P.A.J.; Roda, M.A.; Verspaget, H.W.; Wolfkamp, S.C.; Velde, A.A.T.; Jones, C.W.; et al. Collagen degradation and neutrophilic infiltration: A vicious circle in inflammatory bowel disease. Gut 2013, 63, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Heljasvaara, R.; Nyberg, P.; Luostarinen, J.; Parikka, M.; Heikkilä, P.; Rehn, M.; Sorsa, T.; Salo, T.; Pihlajaniemi, T. Generation of biologically active endostatin fragments from human collagen XVIII by distinct matrix metalloproteases. Exp. Cell Res. 2005, 307, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Tolstanova, G.; Khomenko, T.; Chen, L.; Tarnawski, A.; Szabo, S.; Sandor, Z. Mesalamine restores angiogenic balance in experimental ulcerative colitis by reducing expression of endostatin and angiostatin: Novel molecular mechanism for ther-apeutic action of mesalamine. J. Pharmacol. Exp. Ther. 2009, 331, 1071–1078. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Jilani, S.M.; Nikolova, G.V.; Carpizo, D.; Iruela-Arispe, M.L. Processing of VEGF-A by matrix metalloproteinases regulates bioavailability and vascular patterning in tumors. J. Cell Biol. 2005, 169, 681–691. [Google Scholar] [CrossRef] [Green Version]
- Koller, F.L.; Dozier, E.A.; Nam, K.T.; Swee, M.; Birkland, T.P.; Parks, W.C.; Fingleton, B. Lack of MMP10 exacerbates experi-mental colitis and promotes development of inflammation-associated colonic dysplasia. Lab. Investig. 2012, 92, 1749–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Aono, S.; Lu, W.; Ouellette, A.J.; Hu, X.; Ji, Y.; Wang, L.; Lenz, S.; van Ginkel, F.W.; Liles, M.; et al. A Novel Role for Defensins in Intestinal Homeostasis: Regulation of IL-1β Secretion. J. Immunol. 2007, 179, 1245–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swee, M.; Wilson, C.L.; Wang, Y.; McGuire, J.K.; Parks, W.C. Matrix metalloproteinase-7 (matrilysin) controls neutrophil egress by generating chemokine gradients. J. Leukoc. Biol. 2008, 83, 1404–1412. [Google Scholar] [CrossRef] [Green Version]
- Mäkitalo, L.; Piekkala, M.; Ashorn, M.; Pakarinen, M.; Koivusalo, A.; Karikoski, R.; Natunen, J.; Saarialho-Kere, U.; Rintala, R.; Kolho, K.-L. Matrix metalloproteinases in the restorative proctocolectomy pouch of pediatric ulcerative colitis. World J. Gastroenterol. 2012, 18, 4028–4036. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, N.R.; Smith, A.M. Matrix metalloproteases role in bowel inflammation and inflammatory bowel disease: An up to date review. Inflamm. Bowel Dis. 2014, 20, 2379–2393. [Google Scholar] [CrossRef]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochem-istry. Circ Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontani, F.; Domazetovic, V.; Marcucci, T.; Vincenzini, M.T.; Iantomasi, T. MMPs, ADAMs and their natural inhibitors in inflammatory bowel disease: Involvement of oxidative stress. J. Clin. Gastroenterol. Treat. 2017, 3, 039. [Google Scholar] [CrossRef]
- Andoh, A.; Bamba, S.; Brittan, M.; Fujiyama, Y.; Wright, N.A. Role of intestinal subepithelial myofibroblasts in inflammation and regenerative response in the gut. Pharmacol. Ther. 2007, 114, 94–106. [Google Scholar] [CrossRef]
- Gordon, J.N.; Pickard, K.M.; di Sabatino, A.; Prothero, J.D.; Pender, S.L.F.; Goggin, P.M.; MacDonald, T.T. Matrix metallo-proteinase-3 production by gut IgG plasma cells in chronic inflammatory bowel disease. Inflamm. Bowel Dis. 2008, 14, 195–203. [Google Scholar] [CrossRef]
- Wilson, C.L.; Schmidt, A.P.; Pirilä, E.; Valore, E.V.; Ferri, N.; Sorsa, T.; Ganz, T.; Parks, W.C. Differential Processing of α- and β-Defensin Precursors by Matrix Metalloproteinase-7 (MMP-7). J. Biol. Chem. 2009, 284, 8301–8311. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-D.; Mao, J.W. Expression of matrix metalloproteinase-1 and tumor necrosis factor- in ulcerative colitis. World J. Gas-troenterol. 2007, 13, 5926–5932. [Google Scholar] [CrossRef]
- Drygiannakis, I.; Valatas, V.; Sfakianaki, O.; Bourikas, L.; Manousou, P.; Kambas, K.; Ritis, K.; Kolios, G.; Kouroumalis, E. Proinflammatory cytokines induce crosstalk between colonic epithelial cells and subepithelial myofibroblasts: Implication in intestinal fibrosis. J. Crohn’s Colitis 2013, 7, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Yoo, J.; Perez, C.E.R.; Nie, W.; Sinnett-Smith, J.; Rozengurt, E. Protein kinase D1 mediates synergistic MMP-3 expression induced by TNF-α and bradykinin in human colonic myofibroblasts. Biochem. Biophys. Res. Commun. 2011, 413, 30–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- León, A.J.; Gómez, E.; Garrote, J.A.; Bernardo, D.; Barrera, A.; Marcos, J.L.; Fernández-Salazar, L.; Velayos, B.; Blanco-Quirós, A.; Arranz, E. High levels of proinflammatory cytokines, but not markers of tissue injury, in unaffected intestinal areas frompatients with IBD. Mediat. Inflamm. 2009, 2009, 1–10. [Google Scholar] [CrossRef]
- Pedersen, G.; Saermark, T.; Kirkegaard, T.; Brynskov, J. Spontaneous and cytokine induced expression and activity of matrix metalloproteinases in human colonic epithelium. Clin. Exp. Immunol. 2009, 155, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Medina, C.; Santana, A.; Paz-Cabrera, M.C.; Parra-Blanco, A.; Nicolás, D.; Gimeno-García, A.Z.; Quintero, E. Increased Activity and Expression of Gelatinases in Ischemic Colitis. Dig. Dis. Sci. 2006, 51, 2393–2399. [Google Scholar] [CrossRef]
- Lakatos, G.; Hritz, I.; Varga, M.Z.; Juhász, M.; Miheller, P.; Cierny, G.; Tulassay, Z.; Herszényi, L. The impact of matrix met-alloproteinases and their tissue inhibitors in inflammatory bowel diseases. Dig. Dis. 2012, 30, 289–295. [Google Scholar] [CrossRef]
- Adhikary, S.; Kocieda, V.P.; Yen, J.-H.; Tuma, R.F.; Ganea, D. Signaling through cannabinoid receptor 2 suppresses murine dendritic cell migration by inhibiting matrix metalloproteinase 9 expression. Blood 2012, 120, 3741–3749. [Google Scholar] [CrossRef] [Green Version]
- Castaneda, F.E.; Walia, B.; Vijay–Kumar, M.; Patel, N.R.; Roser, S.; Kolachala, V.L.; Rojas, M.; Wang, L.; Oprea, G.; Garg, P.; et al. Targeted Deletion of Metalloproteinase 9 Attenuates Experimental Colitis in Mice: Central Role of Epithelial-Derived MMP. Gastroenterology 2005, 129, 1991–2008. [Google Scholar] [CrossRef] [PubMed]
- Reinisch, W.; Panés, J.; Khurana, S.; Toth, G.; Hua, F.; Comer, G.M.; Hinz, M.; Page, K.; O’Toole, M.; Moorehead, T.M.; et al. Anrukinzumab, an anti-interleukin 13 monoclonal antibody, in active UC: Efficacy and safety from a phase IIa randomised multicentre study. Gut 2015, 64, 894–900. [Google Scholar] [CrossRef] [PubMed]
- Marshall, D.C.; Lyman, S.K.; McCauley, S.; Kovalenko, M.; Splangler, R.; Liu, C.; Lee, M.; O’Sullivan, C.; Barry-Hamilton, V.; Ghermazien, H.; et al. Selective allosteric inhibition of MMP9 is efficacious in preclinical models of ulcerative colitis and colo-rectal cancer. PLoS ONE 2015, 10, e0127063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bruyn, M.; Breynaert, C.; Arijs, I.; de Hertogh, G.; Geboes, K.; Thijs, G.; Matteoli, G.; Hu, J.; Van Damme, J.; Arnold, B.; et al. Inhibition of gelatinase B/MMP-9 does not attenuate colitis in murine models of inflammatory bowel disease. Nat. Commun. 2017, 8, 15384. [Google Scholar] [CrossRef]
- Shang, K.; Bai, Y.-P.; Wang, C.; Wang, Z.; Gu, H.-Y.; Du, X.; Zhou, X.-Y.; Zheng, C.-L.; Chi, Y.-Y.; Mukaida, N.; et al. Crucial Involvement of Tumor-Associated Neutrophils in the Regulation of Chronic Colitis-Associated Carcinogenesis in Mice. PLoS ONE 2012, 7, e51848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milner, C.; Tongsoongnoen, W.; Rugg, M.; Day, A. The molecular basis of inter-α-inhibitor heavy chain transfer on to hyaluronan: Figure 1. Biochem. Soc. Trans. 2007, 35, 672–676. [Google Scholar] [CrossRef]
- Morgan, A.R.; Han, D.-Y.; Lam, W.-J.; Triggs, C.M.; Fraser, A.G.; Barclay, M.; Gearry, R.B.; Meisner, S.; Stokkers, P.; Boeckxstaens, G.E.; et al. Genetic variations in matrix metalloproteinases may be associated with increased risk of ulcerative colitis. Hum. Immunol. 2011, 72, 1117–1127. [Google Scholar] [CrossRef] [PubMed]
- Meijer, M.J.W.; Mieremet- Ooms, M.A.C.; van Hogezand, R.A.; Lamers, C.B.H.W.; Hommes, D.W.; Verspaget, H.W. Role of matrix metalloproteinase, tissue inhibitor of metalloproteinase and tumor necrosis factor-α single nucleotide gene polymor-phisms in inflammatory bowel disease. World J. Gastroenterol. 2007, 13, 2960–2966. [Google Scholar] [CrossRef]
- López-Hernández, R.; Valdés, M.; Campillo, J.A.; Martinez-Garcia, P.; Salama, H.; Bolarin, J.M.; Banaclocha, H.M.; Moya-quiles, M.R.; Puras, A.M.; Sanches-Torres, A.; et al. Pro- and anti-inflammatory cytokine gene single-nucleotide poly-morphisms in inflammatory bowel disease. Int. J. Immunogenet. 2015, 42, 38–45. [Google Scholar] [CrossRef]
- O’Sullivan, S.; Gilmer, J.F.; Medina, C. Matrix Metalloproteinases in Inflammatory Bowel Disease: An Update. Mediat. Inflamm. 2015, 2015, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.; Jang, J.; Choi, C.; Kang, S.; Bang, K.; Kim, T.; Seo, G.; Cha, J.; Chun, J.; Jung, Y.; et al. Development of a Clinical and Genetic Prediction Model for Early Intestinal Resection in Patients with Crohn’s Disease: Results from the IMPACT Study. J. Clin. Med. 2021, 10, 633. [Google Scholar] [CrossRef]
- Wang, X.; Li, D.; Zhang, Y.; Wu, S.; Tang, F. Costus root granules improve ulcerative colitis through regulation of TGF-β mediation of the PI3K/AKT signaling pathway. Exp. Ther. Med. 2018, 15, 4477–4484. [Google Scholar] [CrossRef]
- Mäkitalo, L.; Sipponen, T.; Kärkkäinen, P.; Kolho, K.-L.; Saarialho-Kere, U. Changes in matrix metalloproteinase (MMP) and tissue inhibitors of metalloproteinases (TIMP) expression profile in Crohn’s disease after immunosuppressive treatment correlate with histological score and calprotectin values. Int. J. Color. Dis. 2009, 24, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- de la Motte, C.A.; Hascall, V.C.; Drazba, J.; Bandyopadhyay, S.K.; Strong, S.A. Mononuclear leukocytes bind to specific hya-luronan structures on colon mucosal smooth muscle cells treated with polyinosinic acid: Polycytidylic acid: Inter-alpha-trypsin inhibitor is crucial to structure and function. Am. J. Pathol. 2003, 163, 121–133. [Google Scholar] [CrossRef]
- Culty, M.; O’Mara, T.E.; Underhill, C.B.; Yeager, H., Jr.; Swartz, R.P. Hyaluronan receptor (CD44) expression and function in human peripheral blood monocytes and alveolar macrophages. J. Leukoc. Biol. 1994, 56, 605–611. [Google Scholar] [CrossRef]
- Kessler, S.; Rho, H.; West, G.; Fiocchi, C.; Drazba, J.; de la Motte, C. Hyaluronan (HA) Deposition Precedes and Promotes Leukocyte Recruitment in Intestinal Inflammation. Clin. Transl. Sci. 2008, 1, 57–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Motte, C.; Nigro, J.; Vasanji, A.; Rho, H.; Kessler, S.; Bandyopadhyay, S.; Danese, S.; Fiocchi, C.; Stern, R. Platelet-Derived Hyaluronidase 2 Cleaves Hyaluronan into Fragments that Trigger Monocyte-Mediated Production of Proinflammatory Cytokines. Am. J. Pathol. 2009, 174, 2254–2264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Docampo, M.J.; Cabrera, J.; Bassols, A. Hyaluronan mediates the adhesion of porcine peripheral blood mononuclear cells to poly (I:C)-treated intestinal cells and modulates their cytokine production. Vet. Immunol. Immunopathol. 2017, 184, 8–17. [Google Scholar] [CrossRef]
- Kessler, S.P.; Obery, D.R.; de la Motte, C. Hyaluronan Synthase 3 Null Mice Exhibit Decreased Intestinal Inflammation and Tissue Damage in the DSS-Induced Colitis Model. Int. J. Cell Biol. 2015, 2015, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauer, M.E.; Glant, T.T.; Mikecz, K.; DeAngelis, P.L.; Haller, F.M.; Husni, M.E.; Hascall, V.C.; Calabro, A. Irreversible Heavy Chain Transfer to Hyaluronan Oligosaccharides by Tumor Necrosis Factor-stimulated Gene-6. J. Biol. Chem. 2013, 288, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Chamouard, P.; Grunebaum, L.; Wiesel, M.L.; Frey, P.L.; Wittersheim, C.; Sapin, R.; Baumann, R.; Cazenave, J.P. Prothrombin fragment 1+2 and thrombin???antithrombin III complex as markers of activation of blood coagulation in inflammatory bowel diseases. Eur. J. Gastroenterol. Hepatol. 1995, 12, 1183–1188. [Google Scholar] [CrossRef]
- Kume, K.; Yamasaki, M.; Tashiro, M.; Yoshikawa, I.; Otsuki, M. Activations of Coagulation and Fibrinolysis Secondary to Bowel Inflammation in Patients with Ulcerative Colitis. Intern. Med. 2007, 46, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Slevin, M.; Krupinski, J.; Gaffney, J.; Matou, S.; West, D.; Delisser, H.; Savani, R.C.; Kumar, S. Hyaluronan-mediated angio-genesis in vascular disease: Uncovering RHAMM and CD44 receptor signaling pathways. Matrix Biol. 2007, 26, 58–68. [Google Scholar] [CrossRef]
- Curry, F.E.; Adamson, R.H. Endothelial Glycocalyx: Permeability Barrier and Mechanosensor. Ann. Biomed. Eng. 2011, 40, 828–839. [Google Scholar] [CrossRef] [Green Version]
- van Hinsbergh, V.W. Endothelium–role in regulation of coagulation and inflammation. Semin. Immunopathol. 2012, 34, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Walley, K.R.; Lukacs, N.W.; Standiford, T.J.; Strieter, R.M.; Kunkel, S.L. Balance of inflammatory cytokines related to severity and mortality of murine sepsis. Infect. Immun. 1996, 64, 4733–4738. [Google Scholar] [CrossRef] [Green Version]
- Vigetti, D.; Genasetti, A.; Karousou, E.; Viola, M.; Moretto, P.; Clerici, M.; Deleonibus, S.; de Luca, G.; Hascall, V.C.; Passi, A. Proinflammatory cytokines induce hyaluronan synthesis and monocyte adhesion in human endothelial cells through hyalu-ronan synthase 2 (HAS2) and the nuclear factor- kappaB (NF-kappaB) pathway. J. Biol. Chem. 2010, 285, 24639–24645. [Google Scholar] [CrossRef] [Green Version]
- Kolodgie, F.D.; Burke, A.P.; Wight, T.N.; Virmani, R. The accumulation of specific types of proteoglycans in eroded plaques: A role in coronary thrombosis in the absence of rupture. Curr. Opin. Lipidol. 2004, 15, 575–582. [Google Scholar] [CrossRef]
- Kolodgie, F.D.; Burke, A.P.; Farb, A.; Weber, D.K.; Kutys, R.; Wight, T.N.; Virmani, R. Differential accumulation of proteo-glycans and hyaluronan in culprit lesions: Insights into plaque erosion. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1642–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmeier, W.; Hynes, R.O. Extracellular matrix proteins in hemostasis and thrombosis. Cold Spring Harb. Perspect. Biol. 2012, 4, a005132. [Google Scholar] [CrossRef]
- Weigel, P.H.; Fuller, G.M.; Leboeuf, R.D. A model for the role of hyaluronic acid and fibrin in the early events during the inflammatory response and wound healing. J. Theor. Biol. 1986, 119, 219–234. [Google Scholar] [CrossRef]
- Chester, D.; Brown, A.C. The role of biophysical properties of provisional matrix proteins in wound repair. Matrix Biol. 2017, 60, 124–140. [Google Scholar] [CrossRef] [PubMed]
- Wall, C.A.; Hanlon, C.R. Experimental enzymatic dissolution of soft tissue hematomas. I. Streptokinase streptodornase and hyaluronidase. Surgery 1957, 42, 536–540. [Google Scholar]
- Roth, K.L. Inhibition of hyaluronidase activity by factors associated with blood clotting. Proc. Soc. Exp. Biol. Med. 1954, 85, 533–537. [Google Scholar] [CrossRef]
- Crawley, J.T.B.; Zanardelli, S.; Chion, C.K.N.K.; Lane, D.A. The central role of thrombin in hemostasis. J. Thromb. Haemost. 2007, 5, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Petrey, A.C.; de la Motte, C.A. Thrombin Cleavage of Inter-α-inhibitor Heavy Chain 1 Regulates Leukocyte Binding to an Inflammatory Hyaluronan Matrix. J. Biol. Chem. 2016, 291, 24324–24334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, Y.; Noda, H.; Okaniwa, N.; Adachi, K.; Shinmura, T.; Nakagawa, S.; Ebi, M.; Ogasawara, N.; Funaki, Y.; Zhuo, L.; et al. Serum-Derived Hyaluronan-Associated Protein Is a Novel Biomarker for Inflammatory Bowel Diseases. Digestion 2017, 95, 146–155. [Google Scholar] [CrossRef]
- Mitchell, J.D.; Lee, R.; Hodakowski, G.T.; Neya, K.; Harringer, W.; Valeri, C.R.; Vlahakes, G.J. Prevention of postoperative pericardial adhesions with a hyaluronic acid coating solution. J. Thorac. Cardiovasc. Surg. 1994, 107, 1481–1488. [Google Scholar] [CrossRef]
- Verheye, S.; Markou, C.P.; Salame, M.Y.; Wan, B.; Kingiii, S.B.; Robinson, K.A.; Chronos, N.A.F.; Hanson, S.R. Reduced Thrombus Formation by Hyaluronic Acid Coating of Endovascular Devices. Arter. Thromb. Vasc. Biol. 2000, 20, 1168–1172. [Google Scholar] [CrossRef] [Green Version]
- Stone, R.L.; Nick, A.M.; McNeish, I.A.; Balkwill, F.; Han, H.D.; Bottsford-Miller, J.; Rupairmoole, R.; Armaiz-Pena, G.N.; Pecot, C.V.; Coward, J.; et al. Paraneoplastic Thrombocytosis in Ovarian Cancer. N. Engl. J. Med. 2012, 366, 610–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timpl, R.; Rohole, H.; Robey, R.G.; Rennard, S.I.; Foidart, J.M.; Martin, G.R. Laminin- a glycoprotein from basement mem-branes. J. Biol. Chem. 1979, 254, 9933–9937. [Google Scholar] [CrossRef]
- Schmehl, K.; Florian, S.; Jacobasch, G.; Salomon, A.; Körber, J. Deficiency of epithelial basement membrane laminin in ulcerative colitis affected human colonic mucosa. Int. J. Color. Dis. 2000, 15, 39–48. [Google Scholar] [CrossRef]
- Gelbmann, C.M.; Mestermann, S.; Gross, V.; Kollinger, M.; Scholmerich, J.; Falk, W. Strictures in Crohn’s disease are charac-terised by an accumulation of mast cells colocalised with laminin but not with fibronectin or vitronectin. Gut 1999, 45, 210–217. [Google Scholar] [CrossRef]
- Koutroubakis, I.E.; Petinaki, E.; Dimoulios, P.; Vardas, E.; Roussomoustakaki, M.; Maniatis, A.N.; Kouroumalis, E.A. Serum laminin and collagen IV in inflammatory bowel disease. J. Clin. Pathol. 2003, 56, 817–820. [Google Scholar] [CrossRef] [PubMed]
- Yablecovitch, D.; Stein, A.; Shabat-Simon, M.; Naftali, T.; Gabay, G.; Laish, I.; Oren, A.; Konikoff, F.M. Soluble Syndecan-1 Levels Are Elevated in Patients with Inflammatory Bowel Disease. Dig. Dis. Sci. 2015, 60, 2419–2426. [Google Scholar] [CrossRef]
- Bass, M.D.; Morgan, M.R.; Humphries, M.J. Syndecans shed their reputation as inert molecules. Sci. Signal. 2009, 12, 18–20. [Google Scholar] [CrossRef] [Green Version]
- Tkachenko, E.; Rhodes, J.M.; Simons, M. Syndecans: New kids on the signaling block. Circ. Res. 2005, 96, 488–500. [Google Scholar] [CrossRef] [Green Version]
- Bernfield, M.; Kokenyesi, R.; Kato, M.; Hinkes, M.; Spring, J.; Gallo, R.; Lose, E. Biology of the syndecans: A family of trans-membrane heparan sulfate proteoglycans. Annu. Rev. Cell Biol. 1992, 8, 333–364. [Google Scholar] [CrossRef]
- Bertrand, J.; Bollmann, M. Soluble syndecans: Biomarkers for diseases and therapeutic options. Br. J. Pharmacol. 2018, 176, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Palaiologou, M.; Delladetsima, I.; Tiniakos, D. CD138 (syndecan-1) expression in health and disease. Histol. Histopathol. 2014, 29, 177–189. [Google Scholar] [PubMed]
- Lemańska-Perek, A.; Adamik, B. Fibronectin and its soluble EDA-FN isoform as biomarkers for inflammation and sepsis. Adv. Clin. Exp. Med. 2019, 28, 1561–1567. [Google Scholar] [CrossRef] [Green Version]
- Potts, J.R.; Campbell, I.D. Fibronectin structure and assembly. Curr. Opin. Cell Biol. 1994, 6, 648–655. [Google Scholar] [CrossRef]
- Allan, A.; Wyke, J.; Allan, R.N.; Morel, P.; Robinson, M.; Scott, D.L.; Alexander-Williams, J. Plasma fibronectin in Crohn’s disease. Gut 1989, 30, 627–633. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Zhou, G.; Lin, J.; Li, L.; Zeng, Z.; Chen, M.; Zhang, S. Serum Biomarkers for Inflammatory Bowel Disease. Front. Med. 2020, 7, 123. [Google Scholar] [CrossRef] [Green Version]
- Petrey, A.C.; de la Motte, C.A. The extracellular matrix in IBD: A dynamic mediator of inflammation. Curr. Opin. Gastroenterol. 2013, 33, 234–238. [Google Scholar] [CrossRef]
- Mortensen, J.; Lindholm, M.; Langholm, L.; Kjeldsen, J.; Bay-Jensen, A.; Karsdal, M.; Manon-Jensen, T. The intestinal tissue homeostasis—The role of extracellular matrix remodeling in inflammatory bowel disease. Expert Rev. Gastroenterol. Hepatol. 2019, 13, 977–993. [Google Scholar] [CrossRef] [PubMed]
- Madisch, A.; Hellmig, S.; Schreiber, S.; Bethke, B.; Stolte, M.; Miehlke, S. Allelic variation of the matrix metalloproteinase-9 gene is associated with collagenous colitis. Inflamm. Bowel Dis. 2011, 17, 2295–2298. [Google Scholar] [CrossRef]
- Sutherland, L.; Singleton, J.; Sessions, J.; Hanauer, S.; Krawitt, E.; Rankin, G.; Summers, R.; Mekhjian, H.; Greenberger, N.; Kelly, M. Double blind, placebo controlled trial of metronidazole in Crohn’s disease. Gut 1991, 32, 1071–1075. [Google Scholar] [CrossRef] [Green Version]
- Matusiewicz, M.; Neubauer, K.; Mierzchala-Pasierb, M.; Gamian, A.; Krzystek-Korpacka, M. Matrix Metalloproteinase-9: Its Interplay with Angiogenic Factors in Inflammatory Bowel Diseases. Dis. Markers 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Onderdonk, A.B.; A Hermos, J.; Bartlett, J.G. The role of the intestinal microflora in experimental colitis. Am. J. Clin. Nutr. 1977, 30, 1819–1825. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-D.; Tan, X.-Y.; Zhang, K. Correlation of Plasma MMP-1 and TIMP-1 Levels and the Colonic Mucosa Expressions in Patients with Ulcerative Colitis. Mediat. Inflamm. 2009, 2009, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Mizoguchi, E.; Nguyen, D.D.; Low, D. Animal models of ulcerative colitis and their application in drug research. Drug Des. Dev. Ther. 2013, 7, 1341–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrence, A.E.; Brabb, T.; Viney, J.L.; Bielefeldt-Ohmann, H.; Treuting, P.; Seamons, A.; Drivdahl, R.; Zeng, W.; Maggio-Price, L. Serum biomarkers in a mouse model of bacterial-induced inflammatory bowel disease. Inflamm. Bowel Dis. 2008, 14, 480–490. [Google Scholar] [CrossRef]
- Burich, A.; Hershberg, R.; Waggie, K.; Zeng, W.; Brabb, T.; Westrich, G.; Viney, J.L.; Maggio-Price, L. Helicobacter-induced inflammatory bowel disease in IL-10- and T cell-deficient mice. Am. J. Physiol. Liver Physiol. 2001, 281, G764–G778. [Google Scholar] [CrossRef] [PubMed]
- Kirkegaard, T.; Hansen, A.; Bruun, E.; Brynskov, J. Expression and localisation of matrix disease & quot. Gut 2004, 53, 701–709. [Google Scholar] [PubMed] [Green Version]
- Efsen, E.; Saermark, T.; Hansen, A.; Bruun, E.; Brynskov, J. Ramiprilate Inhibits Functional Matrix Metalloproteinase Activity in Crohn’s Disease Fistulas. Basic Clin. Pharmacol. Toxicol. 2011, 109, 208–216. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| MMP | Group | Common Name | Role in IBD |
|---|---|---|---|
| MMP-1 | collagenases | collagenase-1 | • prevents intestinal fibrosis |
| MMP-8 | collagenase-2 | • neutrophil infiltration into the intestinal tissue | |
| MMP-13 | collagenase-3 | • activates TNF-α and endostatin [5,6,7,14,15,16] | |
| MMP-2 | gelatinases | gelatinase A | • prevents intestinal fibrosis • produces antiangiogenic factors • supporting epithelial tissue [17] |
| MMP-9 | gelatinase B | • neutrophil infiltration into the intestinal tissue, • chemokine expression • produces antiangiogenic factors • prevents intestinal fibrosis [6,7,8,9,14] | |
| MMP-3 | stromelysins | stromelysin-1 | • activating endostatin [7,18] |
| MMP-10 | stromelysin-2 | • involved in wound healing [10] | |
| MMP-7 | matrilysins | matrilysin-1 | • activating A-defensins • chemokine expression • endostatin production [7,11,12,18,19] |
| MMP-12 | other metalloproteases | macrophage metalloelastase | • degrades intestinal epithelium [13,14,15] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Derkacz, A.; Olczyk, P.; Olczyk, K.; Komosinska-Vassev, K. The Role of Extracellular Matrix Components in Inflammatory Bowel Diseases. J. Clin. Med. 2021, 10, 1122. https://doi.org/10.3390/jcm10051122
Derkacz A, Olczyk P, Olczyk K, Komosinska-Vassev K. The Role of Extracellular Matrix Components in Inflammatory Bowel Diseases. Journal of Clinical Medicine. 2021; 10(5):1122. https://doi.org/10.3390/jcm10051122
Chicago/Turabian StyleDerkacz, Alicja, Paweł Olczyk, Krystyna Olczyk, and Katarzyna Komosinska-Vassev. 2021. "The Role of Extracellular Matrix Components in Inflammatory Bowel Diseases" Journal of Clinical Medicine 10, no. 5: 1122. https://doi.org/10.3390/jcm10051122