
The Serum Concentrations of Hedgehog-Interacting Protein, a Novel Biomarker, Were Decreased in Overweight or Obese Subjects

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials/Subjects and Methods
2.1. Participants
2.2. Data Collection
2.3. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BMI | body mass index |
| ALT | alanine aminotransferase |
| AST | aspartate aminotransferase |
| DBP | diastolic blood pressure |
| eGFR | estimated glomeruli filtration rate |
| FPG | fasting plasma glucose |
| HbA1c | glycated hemoglobin |
| HDL-C | high-density lipoprotein cholesterol |
| Hhip | Hedgehog-interacting protein |
| HOMA-IR | homeostasis model assessment of insulin resistance |
| hsCRP | high-sensitivity C-reactive protein |
| LDL-C | low-density lipoprotein cholesterol |
| SBP | systolic blood pressure |
References
- Al-Goblan, A.S.; Al-Alfi, M.A.; Khan, M.Z. Mechanism linking diabetes mellitus and obesity. Diabetes Metab. Syndr. Obes. 2014, 7, 587–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavie, C.J.; Milani, R.V.; Ventura, H.O. Obesity and Cardiovascular Disease: Risk Factor, Paradox, and Impact of Weight Loss. J. Am. Coll Cardiol. 2009, 53, 1925–1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC Press Releases. Available online: https://www.cdc.gov/media/releases/2017/p1003-vs-cancer-obesity.html (accessed on 25 November 2019).
- Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 25 November 2019).
- Müller, M.J.; Geisler, C. Defining obesity as a disease. Eur J. Clin. Nutr. 2017, 71, 1256–1258. [Google Scholar] [CrossRef]
- De Ferranti, S.; Mozaffarian, D. The perfect storm: Obesity, adipocyte dysfunction, and metabolic consequences. Clin. Chem. 2008, 54, 945–955. [Google Scholar] [CrossRef] [Green Version]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Smelkinson, M.G. The Hedgehog Signaling Pathway Emerges as a Pathogenic Target. J. Dev. Biol. 2017, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Cousin, W.; Fontaine, C.; Dani, C.; Peraldi, P. Hedgehog and adipogenesis: Fat and fiction. Biochimie 2007, 89, 1447–1453. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Long, F. Hedgehog signaling via Gli2 prevents obesity induced by high-fat diet in adult mice. eLife 2017, 6, e31649. [Google Scholar] [CrossRef] [Green Version]
- Suh, J.M.; Gao, X.; McKay, J.; McKay, R.; Salo, Z.; Graff, J.M. Hedgehog signaling plays a conserved role in inhibiting fat formation. Cell Metab. 2006, 3, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, C.; Cousin, W.; Plaisant, M.; Dani, C.; Peraldi, P. Hedgehog Signaling Alters Adipocyte Maturation of Human Mesenchymal Stem Cells. Stem Cells. 2008, 26, 1037–1046. [Google Scholar] [CrossRef]
- Wei, H.; Li, J.; Shi, S.; Zhang, L.; Xiang, A.; Yang, G.; Chu, G. Hhip inhibits proliferation and promotes differentiation of adipocytes through suppressing hedgehog signaling pathway. Biochem Biophys Res. Commun. 2019, 514, 148–156. [Google Scholar] [CrossRef]
- Chuang, P.T.; McMahon, A.P. Vertebrate Hedgehog signalling modulated by induction of a Hedgehog-binding protein. Nature 1999, 397, 617–621. [Google Scholar] [CrossRef]
- Lee, B.N.R.; Son, Y.S.; Lee, D.; Choi, Y.J.; Kwon, S.M.; Chang, H.K.; Kim, P.H.; Cho, J.-Y. Hedgehog-Interacting Protein (HIP) Regulates Apoptosis Evasion and Angiogenic Function of Late Endothelial Progenitor Cells. Sci Rep. 2017, 12449. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.C.; Hung, H.C.; Chen, Y.W.; Cheng, K.P.; Li, C.H.; Lin, C.H.; Chang, C.-J.; Wu, H.-T.; Ou, H.Y. Elevated Hedgehog-Interacting Protein Levels in Subjects with Prediabetes and Type 2 Diabetes. J. Clin. Med. 2019, 8, 1635. [Google Scholar] [CrossRef] [Green Version]
- Health Promotion Administration, Ministry of Health and Welfare, Taiwan. Available online: https://www.hpa.gov.tw/Pages/Detail.aspx?nodeid=542&pid=9737 (accessed on 24 August 2020).
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braune, J.; Weyer, U.; Matz-Soja, M.; Hobusch, C.; Kern, M.; Kunath, A.; Klöting, N.; Kralisch, S.; Blüher, M.; Gebhardt, R.; et al. Hedgehog signalling in myeloid cells impacts on body weight, adipose tissue inflammation and glucose metabolism. Diabetologia 2017, 60, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ge, J.; Cao, H.; Zhang, X.; Guo, Y.; Li, X.; Xia, B.; Yang, G.; Shi, X.E. Leptin Promotes White Adipocyte Browning by Inhibiting the Hh Signaling Pathway. Cells 2019, 8, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todoric, J.; Strobl, B.; Jais, A.; Boucheron, N.; Bayer, M.; Amann, S.; Lindroos, J.; Teperino, R.; Prager, G.; Bilban, M. Cross-Talk Between Interferon-γ and Hedgehog Signaling Regulates Adipogenesis. Diabetes 2011, 60, 1668–1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowatsch, C.; Woolley, R.E.; Kinnebrew, M.; Rohatgi, R.; Siebold, C. Structures of vertebrate Patched and Smoothened reveal intimate links between cholesterol and Hedgehog signalling. Curr Opin Struct Biol. 2019, 57, 204–214. [Google Scholar] [CrossRef]
- Huang, P.; Nedelcu, D.; Watanabe, M.; Jao, C.; Kim, Y.; Liu, J.; Salic, A. Cellular Cholesterol Directly Activates Smoothened in Hedgehog Signaling. Cell 2016, 166, 1176–1187. [Google Scholar] [CrossRef] [Green Version]
- Luchetti, G.; Sircar, R.; Kong, J.H.; Nachtergaele, S.; Sagner, A.; Byrne, E.F.; Covey, D.F.; Siebold, C.; Rohatgi, R. Cholesterol activates the G-protein coupled receptor Smoothened to promote Hedgehog signaling. eLife 2016, 5, e20304. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Baron, R.M.; Hardin, M.; Cho, M.H.; Zielinski, J.; Hawrylkiewicz, I.; Sliwinski, P.; Hersh, C.P.; Mancini, J.D.; Lu, K.; et al. Identification of a chronic obstructive pulmonary disease genetic determinant that regulates HHIP. Hum. Mol. Genet. 2012, 21, 1325–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]


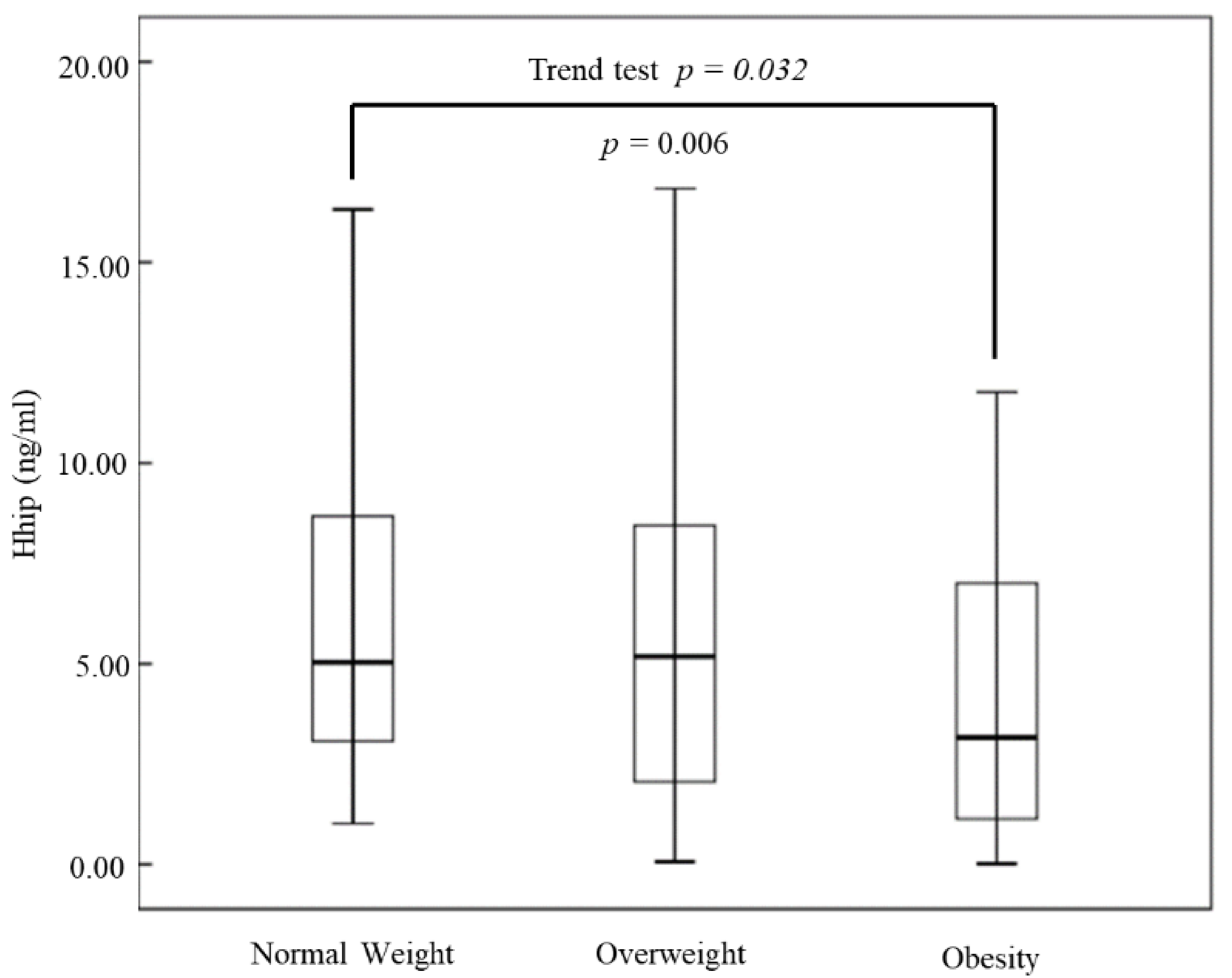{kind=link}
| Clinical Parameters | Normal Weight | Overweight | Obese | p |
|---|---|---|---|---|
| n | 166 | 90 | 38 | |
| Female (%) | 42.8 | 31.1 | 44.7 | 0.147 |
| Hypertension (%) | 17.1 | 23.3 | 27 | 0.272 |
| Diabetes (%) | 24.7 | 31.1 | 31.6 | 0.459 |
| Age (years) | 61.7 ± 11.9 | 60.5 ± 11.8 | 61.4 ± 12.3 | 0.736 |
| Body weight (kg) | 56.4 ± 7 | 67.59 ± 7.47 | 75.79 ± 10.8 | <0.001 |
| Waist circumference (cm) | 78.37 ± 6.82 | 87.51 ± 5.77 | 94.43 ± 8.44 | <0.001 |
| Body-mass index (kg/m2) | 21.65 ± 1.87 | 25.44 ± 0.88 | 28.49 ± 1.75 | <0.001 |
| SBP (mmHg) | 125.3 ± 18.1 | 128.5 ± 16.1 | 131.5 ± 16.7 | 0.098 |
| DBP (mmHg) | 72.2 ± 10.4 | 74.9 ± 9.7 | 76.8 ± 12.0 | 0.022 |
| FPG (mg/dL) | 101.78 ± 43.28 | 107.36 ± 35.33 | 116.47 ± 49.68 | 0.131 |
| Post-load 2-h glucose (mg/dL) | 155.34 ± 80.21 | 147.54 ± 68.90 | 175.41 ± 97.48 | 0.234 |
| HbA1c (%) | 6.13 ± 1.28 | 6.24 ± 1.05 | 6.79 ± 2.09 | 0.026 |
| ALT (U/L) | 25.51 ± 19.50 | 35.19 ± 53.50 | 24.89 ± 10.43 | 0.068 |
| AST (U/L) | 27.23 ± 13.70 | 30.56 ± 41.84 | 24.68 ± 25.45 | 0.429 |
| Creatinine (mg/dL) | 0.87 ± 0.20 | 0.87 ± 0.18 | 0.88 ± 0.20 | 0.886 |
| eGFR | 90.63 ± 19.76 | 91.72 ± 15.65 | 88.22 ± 19.24 | 0.62 |
| hsCRP (mg/L) | 3.68 ± 7.52 | 3.73 ± 6.27 | 4.48 ± 4.71 | 0.809 |
| HDL-C (mg/dL) | 55.73 ± 15.39 | 49.98 ± 13.55 | 51.74 ± 12.49 | 0.008 |
| LDL-C (mg/dL) | 126.33 ± 33.89 | 129.02 ± 35.91 | 131.28 ± 44.65 | 0.694 |
| Triglycerides (mg/dL) | 110.47 ± 68.96 | 127.56 ± 60.71 | 144.94 ± 70.84 | 0.008 |
| Triglycerides (mg/dL) * | 1.99 ± 0.21 | 2.06 ± 0.20 | 2.12 ± 0.20 | <0.001 |
| Cholesterol (mg/dL) | 204.16 ± 39.555 | 204.51 ± 41.545 | 212.00 ± 47.594 | 0.56 |
| HOMA-IR | 0.50 ± 0.47 | 0.97 ± 1.50 | 1.26 ± 1.011 | <0.001 |
| Variable | Model 1 | Model 2 | Model 3 | |||
|---|---|---|---|---|---|---|
| β (95% CI) | p | β (95% CI) | p | β (95% CI) | p | |
| Age (years) | −0.029 (−0.056~0.033) | 0.612 | −0.034 (−0.058~0.032) | 0.565 | −0.006 (−0.062~0.057) | 0.938 |
| Sex | 0.143 (0.278~2.412) | 0.014 | 0.134 (0.182~2.350) | 0.022 | 0.158 (0.204~2.857) | 0.024 |
| Body-mass index | −0.144 (−0.397~−0.046) | 0.013 | −0.147 (−0.411~−0.041) | 0.017 | ||
| OW vs. NW | −0.181 (−3.311~−0.400) | 0.013 | ||||
| OB vs. NW | −0.311 (−6.393~−2.384) | <0.001 | ||||
| Fasting glucose (mg/dL) | 0.024 (−0.010~0.015) | 0.694 | 0.007 (−0.015~0.016) | 0.925 | ||
| Insulin (mIU/L) | −0.011 (−0.210~0.175) | 0.859 | 0.049 (−0.175~0.354) | 0.503 | ||
| hsCRP (mg/L) | 0.016 (−0.078~0.100) | 0.809 | ||||
| SBP (mmHg) | 0.06 (−0.021~0.054) | 0.397 | ||||
| eGFR | −0.026 (−0.042~0.029) | 0.715 | ||||
| ALT (U/L) | −0.07 (−0.027~0.008) | 0.296 | ||||
| CHOL (mg/dL) | −0.063 (−0.147~0.133) | 0.923 | ||||
| TGs (mg/dL) * | 0.029 (−8.388~9.701) | 0.886 | ||||
| HDL-C (mg/dL) | −0.101 (−0.182~0.116) | 0.663 | ||||
| LDL-C (mg/dL) | 0.129 (−0.126~0.158) | 0.821 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chou, H.-W.; Hung, H.-C.; Lin, C.-H.; Lin, A.-C.; Du, Y.-F.; Cheng, K.-P.; Li, C.-H.; Chang, C.-J.; Wu, H.-T.; Ou, H.-Y. The Serum Concentrations of Hedgehog-Interacting Protein, a Novel Biomarker, Were Decreased in Overweight or Obese Subjects. J. Clin. Med. 2021, 10, 742. https://doi.org/10.3390/jcm10040742
Chou H-W, Hung H-C, Lin C-H, Lin A-C, Du Y-F, Cheng K-P, Li C-H, Chang C-J, Wu H-T, Ou H-Y. The Serum Concentrations of Hedgehog-Interacting Protein, a Novel Biomarker, Were Decreased in Overweight or Obese Subjects. Journal of Clinical Medicine. 2021; 10(4):742. https://doi.org/10.3390/jcm10040742
Chicago/Turabian StyleChou, Hsuan-Wen, Hao-Chang Hung, Ching-Han Lin, An-Chi Lin, Ye-Fong Du, Kai-Pi Cheng, Chung-Hao Li, Chih-Jen Chang, Hung-Tsung Wu, and Horng-Yih Ou. 2021. "The Serum Concentrations of Hedgehog-Interacting Protein, a Novel Biomarker, Were Decreased in Overweight or Obese Subjects" Journal of Clinical Medicine 10, no. 4: 742. https://doi.org/10.3390/jcm10040742