
Cigarette Smoking, miR-27b Downregulation, and Peripheral Artery Disease: Insights into the Mechanisms of Smoking Toxicity

 ,
,  ,
,  ,
,  ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Recruitment of Participants
2.2. Data Collection
2.3. Candidate miRNAs
2.4. Quantification of Expression Levels of Candidate miRNAs
2.5. Statistical Analysis
3. Results
3.1. Characteristics of Participants According to the Presence of PAD and Cigarette-Smoking Status
3.2. Cigarette Smoking Was Independently Associated with the Presence of PAD
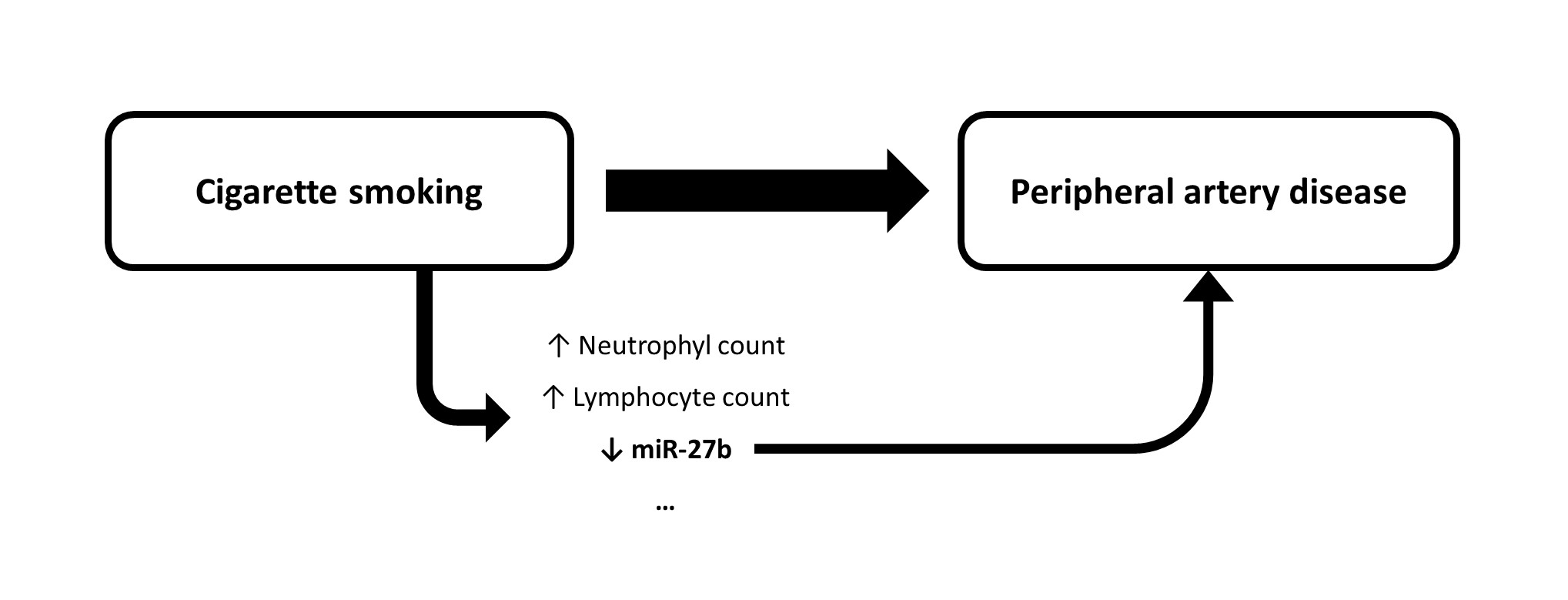
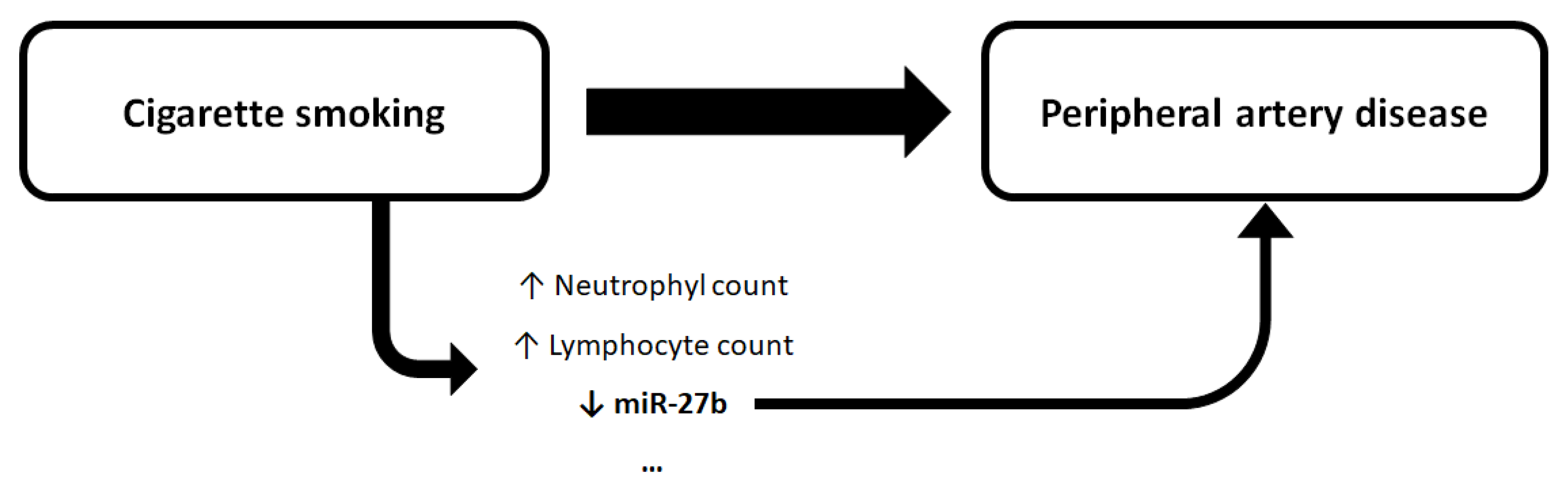
3.3. MiR-27b Was Dysregulated in Active Smokers Independently of Other Metabolic and Inflammatory Parameters
3.4. MiR-27b Was Independently Associated with the Presence of PAD
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siasos, G.; Tsigkou, V.; Kokkou, E.; Oikonomou, E.; Vavuranakis, M.; Vlachopoulos, C.; Verveniotis, A.; Limperi, M.; Genimata, V.; Papavassiliou, A.G.; et al. Smoking and Atherosclerosis: Mechanisms of Disease and New Therapeutic Approaches. Curr. Med. Chem. 2014, 21, 3936–3948. [Google Scholar] [CrossRef]
- Lu, J.T.; Creager, M.A. The relationship of cigarette smoking to peripheral arterial disease. Rev. Cardiovasc. Med. 2004, 5, 189–193. [Google Scholar]
- Feinberg, M.W.; Moore, K.J. MicroRNA Regulation of Atherosclerosis. Circ. Res. 2016, 118, 703–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willinger, C.M.; Rong, J.; Tanriverdi, K.; Courchesne, P.L.; Huan, T.; Wasserman, G.A.; Lin, H.; Dupuis, J.; Joehanes, R.; Jones, M.R.; et al. MicroRNA Signature of Cigarette Smoking and Evidence for a Putative Causal Role of MicroRNAs in Smoking-Related Inflammation and Target Organ Damage. Circ. Cardiovasc. Genet. 2017, 10. [Google Scholar] [CrossRef]
- Kaur, G.; Begum, R.; Thota, S.; Batra, S. A systematic review of smoking-related epigenetic alterations. Arch. Toxicol. 2019, 93, 2715–2740. [Google Scholar] [CrossRef]
- Pereira-da-Silva, T.; Coutinho Cruz, M.; Carrusca, C.; Cruz Ferreira, R.; Napoleão, P.; Mota Carmo, M. Circulating mi-croRNA profiles in different arterial territories of stable atherosclerotic disease: A systematic review. Am. J. Cardiovasc. Dis. 2018, 8, 1–13. [Google Scholar]
- Andreou, I.; Sun, X.; Stone, P.H.; Edelman, E.R.; Feinberg, M.W. miRNAs in atherosclerotic plaque initiation, progression, and rupture. Trends Mol. Med. 2015, 21, 307–318. [Google Scholar] [CrossRef]
- Chen, L.-J.; Lim, S.H.; Yeh, Y.-T.; Lien, S.-C.; Chiu, J.-J. Roles of microRNAs in atherosclerosis and restenosis. J. Biomed. Sci. 2012, 19, 79. [Google Scholar] [CrossRef] [Green Version]
- Navickas, R.; Gal, D.; Laucevičius, A.; Taparauskaitė, A.; Zdanytė, M.; Holvoet, P. Identifying circulating microRNAs as biomarkers of cardiovascular disease: A systematic review. Cardiovasc. Res. 2016, 111, 322–337. [Google Scholar] [CrossRef]
- Stather, P.W.; Sylvius, N.; Wild, J.B.; Choke, E.; Sayers, R.D.; Bown, M.J. Differential MicroRNA Expression Profiles in Peripheral Arterial Disease. Circ. Cardiovasc. Genet. 2013, 6, 490–497. [Google Scholar] [CrossRef] [Green Version]
- Signorelli, S.S.; Volsi, G.L.; Pitruzzella, A.; Fiore, V.; Mangiafico, M.; Vanella, L.; Parenti, R.; Rizzo, M.; Volti, G.L. Circulating miR-130a, miR-27b, and miR-210 in Patients With Peripheral Artery Disease and Their Potential Relationship With Oxidative Stress. Angiology 2016, 67, 945–950. [Google Scholar] [CrossRef]
- Wang, J.; Song, Y.; Zhang, Y.; Xiao, H.; Sun, Q.; Hou, N.; Guo, S.; Wang, Y.; Fan, K.; Zhan, D.; et al. Cardiomyocyte overexpression of miR-27b induces cardiac hypertrophy and dysfunction in mice. Cell Res. 2011, 22, 516–527. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Chen, W.; Cao, G.; Dong, Z.; Xu, J.; Luo, T.; Zhang, S. MicroRNA-27b inhibits cell proliferation in oral squamous cell carcinoma by targeting FZD7 and Wnt signaling pathway. Arch. Oral Biol. 2017, 83, 92–96. [Google Scholar] [CrossRef]
- Murata, Y.; Yamashiro, T.; Kessoku, T.; Jahan, I.; Usuda, H.; Tanaka, T.; Okamoto, T.; Nakajima, A.; Wada, K. Up-Regulated MicroRNA-27b Promotes Adipocyte Differentiation via Induction of Acyl-CoA Thioesterase 2 Expression. BioMed Res. Int. 2019, 2019, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Cazalla, D.; Steitz, J.A. Down-Regulation of a Host microRNA by a Viral Noncoding RNA. Cold Spring Harb. Symp. Quant. Biol. 2010, 75, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.E.; Riley, K.J.; Iwasaki, A.; Steitz, J.A. Alternative Capture of Noncoding RNAs or Protein-Coding Genes by Herpesviruses to Alter Host T Cell Function. Mol. Cell 2014, 54, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Marcinowski, L.; Tanguy, M.; Krmpotic, A.; Rädle, B.; Lisnić, V.J.; Tuddenham, L.; Chane-Woon-Ming, B.; Ruzsics, Z.; Erhard, F.; Benkartek, C.; et al. Degradation of Cellular miR-27 by a Novel, Highly Abundant Viral Transcript Is Important for Efficient Virus Replication In Vivo. PLoS Pathog. 2012, 8, e1002510. [Google Scholar] [CrossRef] [Green Version]
- Machitani, M.; Sakurai, F.; Wakabayashi, K.; Nakatani, K.; Tachibana, M.; Mizuguchi, H. MicroRNA miR-27 Inhibits Adenovirus Infection by Suppressing the Expression of SNAP25 and TXN2. J. Virol. 2017, 91, e00159–e00217. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Li, L.; Zhang, M.; Cheng, H.-P.; Gong, D.; Lv, Y.-C.; Yao, F.; He, P.-P.; Ouyang, X.-P.; Lan, G.; et al. MicroRNA-27 Prevents Atherosclerosis by Suppressing Lipoprotein Lipase-Induced Lipid Accumulation and Inflammatory Response in Apolipoprotein E Knockout Mice. PLoS ONE 2016, 11, e0157085. [Google Scholar] [CrossRef] [Green Version]
- Vickers, K.C.; Shoucri, B.M.; Levin, M.G.; Wu, H.; Pearson, D.S.; Osei-Hwedieh, D.; Collins, F.S.; Remaley, A.T.; Sethupathy, P. MicroRNA-27b is a regulatory hub in lipid metabolism and is altered in dyslipidemia. Hepatology 2013, 57, 533–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, S.; Song, Z.; Wu, Y.; Gao, Y.; Gao, M.; Liu, F.; Wang, F.; Zhang, Y. MicroRNA-27b Modulates Inflammatory Response and Apoptosis during Mycobacterium tuberculosis Infection. J. Immunol. 2018, 200, 3506–3518. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.; Shen, Y.; Wei, X.; Zhang, F.; Liu, Y.; Ma, L. Inhibitory effect of microRNA-27b on interleukin 17 (IL-17)-induced monocyte chemoattractant protein-1 (MCP1) expression. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Veliceasa, D.; Biyashev, D.; Qin, G.; Misener, S.; Mackie, A.R.; Kishore, R.; Volpert, O.V. Therapeutic manipulation of angiogenesis with miR-27b. Vasc. Cell 2015, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Boon, R.A.; Hergenreider, E.; Dimmeler, S. Atheroprotective mechanisms of shear stress-regulated microRNAs. Thromb. Haemost. 2012, 108, 616–620. [Google Scholar] [CrossRef]
- Kuehbacher, A.; Urbich, C.; Zeiher, A.M.; Dimmeler, S. Role of Dicer and Drosha for Endothelial MicroRNA Expression and Angiogenesis. Circ. Res. 2007, 101, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Urbich, C.; Kaluza, D.; Frömel, T.; Knau, A.; Bennewitz, K.; Boon, R.A.; Bonauer, A.; Doebele, C.; Boeckel, J.-N.; Hergenreider, E.; et al. MicroRNA-27a/b controls endothelial cell repulsion and angiogenesis by targeting semaphorin 6A. Blood 2012, 119, 1607–1616. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.Y. Doppler ultrasonography of the lower extremity arteries: Anatomy and scanning guidelines. Ultrasonography 2017, 36, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Aboyans, V.; Ricco, J.-B.; Bartelink, M.-L.E.; Björck, M.; Brodmann, M.; Cohnert, T.; Collet, J.-P.; Czerny, M.; De Carlo, M.; Debus, S.; et al. Editor’s Choice—2017 ESC Guidelines on the Diagnosis and Treatment of Peripheral Arterial Diseases, in collaboration with the European Society for Vascular Surgery (ESVS). Eur. J. Vasc. Endovasc. Surg. 2018, 55, 305–368. [Google Scholar] [CrossRef] [Green Version]
- Collins, R.; Burch, J.; Cranny, G.; Aguiar-Ibáñez, R.; Craig, D.; Wright, K.; Berry, E.; Gough, M.; Kleijnen, J.; Westwood, M. Duplex ultrasonography, magnetic resonance angiography, and computed tomography angiography for diagnosis and assessment of symptomatic, lower limb peripheral arterial disease: Systematic review. BMJ 2007, 334, 1257. [Google Scholar] [CrossRef] [Green Version]
- Hackshaw, A.; Morris, J.K.; Boniface, S.; Tang, J.-L.; Milenković, D. Low cigarette consumption and risk of coronary heart disease and stroke: Meta-analysis of 141 cohort studies in 55 study reports. BMJ 2018, 360, 5855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deo, A.; Carlsson, J.; Lindlöf, A. How to Choose A Normalization Strategy for Mirna Quantitative Real-Time (QPCR) Arrays. J. Bioinform. Comput. Biol. 2011, 9, 795–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfinger, R.D.; Beedanagari, S.; Boitier, E.; Chen, T.; Couttet, P.; Ellinger-Ziegelbauer, H.; Guillemain, G.; Mariet, C.; Mouritzen, P.; O’Lone, R.; et al. Two approaches for estimating the lower limit of quantitation (LLOQ) of microRNA levels assayed as exploratory biomarkers by RT-qPCR. BMC Biotechnol. 2018, 18, 6. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Shao, S.; Geng, H.; Yu, Y.; Wang, C.; Liu, Z.; Yu, C.; Jiang, X.; Deng, Y.; Gao, L.; et al. Expression Profiles of Six Circulating MicroRNAs Critical to Atherosclerosis in Patients With Subclinical Hypothyroidism: A Clinical Study. J. Clin. Endocrinol. Metab. 2014, 99, 766–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, D.; Narang, R.; Sreenivas, V.; Rastogi, V.; Bhatia, J.; Saluja, D.; Srivastava, K. Circulatory miR-133b and miR-21 as Novel Biomarkers in Early Prediction and Diagnosis of Coronary Artery Disease. Genes 2020, 11, 164. [Google Scholar] [CrossRef] [Green Version]
- Vegter, E.L.; Ovchinnikova, E.S.; Van Veldhuisen, D.J.; Jaarsma, T.; Berezikov, E.; Van Der Meer, P.; Voors, A.A. Low circulating microRNA levels in heart failure patients are associated with atherosclerotic disease and cardiovascular-related rehospitalizations. Clin. Res. Cardiol. 2017, 106, 598–609. [Google Scholar] [CrossRef] [Green Version]
- Stather, P.W.; Sylvius, N.; Sidloff, D.A.; Dattani, N.; Verissimo, A.; Wild, J.B.; Butt, H.Z.; Choke, E.; Sayers, R.D.; Bown, M.J. Identification of microRNAs associated with abdominal aortic aneurysms and peripheral arterial disease. BJS 2015, 102, 755–766. [Google Scholar] [CrossRef]
- Huang, Y.-Q.; Ying-Ling, Z.; Chen, J.-Y.; Zhou, Y.-L.; Cai, A.-P.; Huang, C.; Feng, Y.-Q. The Association of Circulating MiR-29b and Interleukin-6 with Subclinical Atherosclerosis. Cell. Physiol. Biochem. 2017, 44, 1537–1544. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Takahashi, K.; Yokota, S.-I.; Tatsumi, N.; Fukami, T.; Yokoi, T.; Nakajima, M. Cigarette smoking substantially alters plasma microRNA profiles in healthy subjects. Toxicol. Appl. Pharmacol. 2013, 272, 154–160. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Yamada, H.; Nagura, A.; Ohashi, K.; Ishikawa, H.; Yamazaki, M.; Ando, Y.; Ichino, N.; Osakabe, K.; Sugimoto, K.; et al. Association of cigarette smoking with serum microRNA expression among middle-aged Japanese adults. Fujita Med J. 2013, 2, 1–5. [Google Scholar] [CrossRef]
- Bhat, M.Y.; Advani, J.; Rajagopalan, P.; Patel, K.; Nanjappa, V.; Solanki, H.S.; Patil, A.H.; Bhat, F.A.; Mathur, P.P.; Nair, B.; et al. Cigarette smoke and chewing tobacco alter expression of different sets of miRNAs in oral keratinocytes. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Fazeli, S.; Motovali-Bashi, M.; Peymani, M.; Hashemi, M.-S.; Etemadifar, M.; Nasr-Esfahani, M.H.; Ghaedi, K. A compound downregulation of SRRM2 and miR-27a-3p with upregulation of miR-27b-3p in PBMCs of Parkinson’s patients is associated with the early stage onset of disease. PLoS ONE 2020, 15, e0240855. [Google Scholar] [CrossRef]
- Shafiei, J.; Heidari, F.; Khashen, E.; Ghandehari-Alavijeh, R.; Darmishonnejad, Z. Distinctive deregulation of miR-27a and miR-27b in relapsing remitting multiple sclerosis. J. Bas. Res. Med. Sci. 2020, 7, 1–6. [Google Scholar]
- Ma, M.; Yin, Z.; Zhong, H.; Liang, T.; Guo, L. Analysis of the expression, function, and evolution of miR-27 isoforms and their responses in metabolic processes. Genomics 2019, 111, 1249–1257. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Ridker, P.M.; Hansson, G.K. Inflammation in Atherosclerosis. J. Am. Coll. Cardiol. 2009, 54, 2129–2138. [Google Scholar] [CrossRef] [Green Version]
- Ross, R. Atherosclerosis—An Inflammatory Disease. N. Engl. J. Med. 1999, 340, 115–126. [Google Scholar] [CrossRef]
- Kaur, A.; Mackin, S.T.; Schlosser, K.; Wong, F.L.; Elharram, M.; Delles, C.; Stewart, D.J.; Dayan, N.; Landry, T.; Pilote, L. Systematic review of microRNA biomarkers in acute coronary syndrome and stable coronary artery disease. Cardiovasc. Res. 2019, 116, 1113–1124. [Google Scholar] [CrossRef]
- Cheng, H.S.; Sivachandran, N.; Lau, A.; Boudreau, E.; Zhao, J.L.; Baltimore, D.; Delgado-Olguin, P.; Cybulsky, M.I.; Fish, J.E. Micro RNA -146 represses endothelial activation by inhibiting pro-inflammatory pathways. EMBO Mol. Med. 2013, 5, 1017–1034. [Google Scholar] [CrossRef]
- Wang, H.-J.; Huang-Joe, W.; Shih, Y.-Y.; Wu, H.-Y.; Peng, C.-T.; Lo, W.-Y. MicroRNA-146a Decreases High Glucose/Thrombin-Induced Endothelial Inflammation by Inhibiting NAPDH Oxidase 4 Expression. Mediat. Inflamm. 2014, 2014, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Zhan, J.-K.; Wang, Y.-J.; Tan, P.; Chen, Y.-Y.; Deng, H.-Q.; Liu, Y.-S. Function, Role, and Clinical Application of MicroRNAs in Vascular Aging. BioMed Res. Int. 2016, 2016, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhao, L.; He, X.; Yang, T.; Yang, K. MiR-21 inhibits c-Ski signaling to promote the proliferation of rat vascular smooth muscle cells. Cell. Signal. 2014, 26, 724–729. [Google Scholar] [CrossRef]
- Weber, M.; Baker, M.B.; Moore, J.P.; Searles, C.D. MiR-21 is induced in endothelial cells by shear stress and modulates apoptosis and eNOS activity. Biochem. Biophys. Res. Commun. 2010, 393, 643–648. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.; Zhao, Y.; Yu, L.; Xu, S.; Fu, G. MicroRNA-21 mediates the rapamycin-induced suppression of endothelial proliferation and migration. FEBS Lett. 2013, 587, 378–385. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wang, K.-C.; Wu, W.; Subramaniam, S.; Shyy, J.Y.-J.; Chiu, J.-J.; Li, J.Y.-S.; Chien, S. MicroRNA-21 targets peroxisome proliferators-activated receptor- in an autoregulatory loop to modulate flow-induced endothelial inflammation. Proc. Natl. Acad. Sci. USA 2011, 108, 10355–10360. [Google Scholar] [CrossRef] [Green Version]
- Ulrich, V.; Rotllan, N.; Araldi, E.; Luciano, A.; Skroblin, P.; Abonnenc, M.; Perrotta, P.; Yin, X.; Bauer, A.; Leslie, K.L.; et al. Chronic miR-29 antagonism promotes favorable plaque remodeling in atherosclerotic mice. EMBO Mol. Med. 2016, 8, 643–653. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Huang, A.; Ferruzzi, J.; Mecham, R.P.; Starcher, B.C.; Tellides, G.; Humphrey, J.D.; Giordano, F.J.; Niklason, L.E.; Sessa, W.C. Inhibition of MicroRNA-29 Enhances Elastin Levels in Cells Haploinsufficient for Elastin and in Bioengineered Vessels—Brief Report. Arter. Thromb. Vasc. Biol. 2012, 32, 756–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schober, A.; Nazari-Jahantigh, M.; Wei, Y.; Bidzhekov, K.; Gremse, F.; Grommes, J.; Megens, R.T.A.; Heyll, K.; Noels, H.; Hristov, M.; et al. MicroRNA-126-5p promotes endothelial proliferation and limits atherosclerosis by suppressing Dlk1. Nat. Med. 2014, 20, 368–376. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Kim, C.W.; Simmons, R.D.; Jo, H. Role of Flow-Sensitive microRNAs in Endothelial Dysfunction and Atherosclerosis. Arter. Thromb. Vasc. Biol. 2014, 34, 2206–2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Hernando, C.; Suárez, Y. MicroRNAs in endothelial cell homeostasis and vascular disease. Curr. Opin. Hematol. 2018, 25, 227–236. [Google Scholar] [CrossRef]
- Small, E.M.; Sutherland, L.B.; Rajagopalan, K.N.; Wang, S.; Olson, E.N. MicroRNA-218 Regulates Vascular Patterning by Modulation of Slit-Robo Signaling. Circ. Res. 2010, 107, 1336–1344. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.S.; Gamazon, E.R.; Ziliak, D.; Wen, Y.; Im, H.K.; Zhang, W.; Wing, C.; Duan, S.; Bleibel, W.K.; Cox, N.J.; et al. Population differences in microRNA expression and biological implications. RNA Biol. 2011, 8, 692–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawlings-Goss, R.A.; Campbell, M.C.; Tishkoff, S.A. Global population-specific variation in miRNA associated with cancer risk and clinical biomarkers. BMC Med. Genom. 2014, 7, 53. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Characteristics of Participants | Without Peripheral Artery Disease | With Peripheral Artery Disease | p-Value |
|---|---|---|---|
| n | 58 | 36 | |
| Clinical characteristics | |||
| Age, years | 61 (53–70) | 68 (60–73) | 0.009 |
| Male, n (%) | 51 (88) | 33 (92) | 0.419 |
| Hypertension, n (%) | 42 (72) | 36 (100) | <0.001 |
| Dyslipidemia, n (%) | 48 (83) | 35 (97) | 0.031 |
| Diabetes mellitus, n (%) | 15 (26) | 17 (47) | 0.029 |
| Cigarette-smoking status, n (%) | 0.001 | ||
| Never smoker | 39 (67) | 12 (33) | |
| Prior smoker | 15 (26) | 13 (36) | |
| Active smoker | 4 (7) | 11 (31) | |
| LVEF > 50%, n (%) | 58 (100) | 36 (100) | – |
| Antiplatelet therapy, n (%) | 37 (64) | 35 (97) | <0.001 |
| Statin therapy, n (%) | 42 (72) | 32 (91) | 0.023 |
| Coronary artery disease, n (%) | 32 (55) | 36 (100) | <0.001 |
| Carotid artery disease, n (%) | 12 (55) | 18 (50) | 0.003 |
| Peripheral artery disease | |||
| Bilateral disease, n (%) | 0 (0) | 25 (69) | <0.001 |
| Prior bypass surgery, n (%) | 0 (0) | 8 (22) | <0.001 |
| Laboratory parameters | |||
| Hemoglobin, g/dL | 13.8 (1.5) | 13.6 (1.6) | 0.586 |
| Leukocyte count, 109/L | 7.0 (1.9) | 7.7 (1.7) | 0.081 |
| Neutrophil count, 109/L | 4.0 (1.7) | 4.5 (1.4) | 0.192 |
| Lymphocyte count, 109/L | 1.9 (1.6–2.3) | 2.2 (1.6–2.7) | 0.100 |
| Neutrophil/lymphocyte ratio | 2.2 (1.0) | 2.2 (1.0) | 0.943 |
| Platelet count, 109/L | 223 (58) | 227 (44) | 0.749 |
| Fasting glycemia, mg/dL | 92 (83–104) | 90 (82–121) | 0.978 |
| Percentage of glycosylated hemoglobin | 5.7 (5.4–6.2) | 5.9 (5.5–7.4) | 0.295 |
| Creatinine, mg/dL | 0.85 (0.77–0.97) | 0.94 (0.80–1.34) | 0.036 |
| Total cholesterol, mg/dL | 164 (130–206) | 166 (147–205) | 0.781 |
| LDL-cholesterol, mg/dL | 92 (72–130) | 109 (83–135) | 0.250 |
| HDL-cholesterol, mg/dL | 43 (34–51) | 36 (31–43) | 0.062 |
| Triglycerides, mg/dL | 115 (71–163) | 117 (88–178) | 0.429 |
| C-reactive protein, mg/L | 4.0 (1.9) | 3.6 (1.5) | 0.529 |
| miRNAs1 | |||
| miR-21 | 14.89 (4.51) | 15.99 (4.65) | 0.282 |
| miR-27b | 18.23 (14.58–21.59) | 22.32 (16.50–24.13) | 0.032 |
| miR-29a | 20.42 (3.57) | 21.80 (3.12) | 0.152 |
| miR-126 | 16.89 (14.89–22.46) | 19.69 (16.28–24.06) | 0.060 |
| miR-146 | 18.70 (3.40) | 20.48 (4.23) | 0.048 |
| miR-218 | 22.69 (22.48–23.33) | 14.49 (−8.6–23.50) | 0.186 |
| Characteristics of Participants | Never Smokers | Prior Smokers | Active Smokers | p-Value |
|---|---|---|---|---|
| n | 51 | 28 | 15 | |
| Clinical characteristics | ||||
| Age, years | 65 (56–73) | 67 (58–71) | 59 (53–68) | 0.415 |
| Male, n (%) | 43 (84) | 28 (100) | 13 (87) | 0.090 |
| Hypertension, n (%) | 36 (71) | 28 (100) | 14 (93) | 0.002 |
| Dyslipidemia, n (%) | 43 (84) | 26 (93) | 14 (93) | 0.424 |
| Diabetes mellitus, n (%) | 17 (33) | 10 (36) | 5 (33) | 0.975 |
| LVEF > 50%, n (%) | 51 (100) | 28 (100) | 15 (100) | – |
| Antiplatelet therapy, n (%) | 36 (71) | 24 (86) | 12 (80) | 0.298 |
| Statin therapy, n (%) | 37 (73) | 24 (86) | 13 (93) | 0.156 |
| Coronary artery disease, n (%) | 31 (61) | 24 (86) | 13 (86) | 0.024 |
| Carotid artery disease, n (%) | 14 (51) | 10 (36) | 6 (40) | 0.576 |
| Peripheral artery disease | ||||
| Number of patients | 12 (24) | 13 (46) | 11 (73) | 0.001 |
| Bilateral disease, n (%) | 8 (16) | 11 (39) | 6 (40) | 0.002 |
| Prior bypass surgery, n (%) | 2 (4) | 5 (18) | 1 (7) | 0.101 |
| Laboratory parameters | ||||
| Hemoglobin, g/dL | 13.4 (1.5) | 14.1 (1.4) | 14.1 (1.4) | 0.063 |
| Leukocyte count, 109/L | 6.6 (1.5) | 7.3 (1.8) | 9.2 (1.8) | <0.001 1,2 |
| Neutrophil count, 109/L | 3.8 (3.0–4.4) | 3.9 (3.2–5.3) | 4.6 (3.7–5.8) | <0.001 1,2 |
| Lymphocyte count, 109/L | 1.8 (1.5–2.2) | 2.1 (1.6–2.4) | 2.3 (1.8–3.5) | 0.027 1 |
| Neutrophil/lymphocyte ratio | 2.0 (1.5–3.0) | 2.2 (1.7–2.7) | 2.3 (1.9–2.8) | 0.568 |
| Platelet count, 109/L | 219 (51) | 218 (54) | 255 (51) | 0.058 |
| Fasting glycemia, mg/dL | 92 (83–116) | 91 (85–112) | 89 (72–117) | 0.565 |
| Percentage of glycosylated hemoglobin | 5.7 (5.3–7.1) | 5.8 (5.4–6.2) | 5.9 (5.6–6.1) | 0.787 |
| Creatinine, mg/dL | 0.86 (0.78–1.08) | 0.86 (0.76–1.00) | 1.01 (0.75–1.71) | 0.447 |
| Total cholesterol, mg/dL | 178 (51) | 154 (43) | 172 (48) | 0.060 |
| LDL-cholesterol, mg/dL | 108 (41) | 89 (38) | 116 (32) | 0.059 |
| HDL-cholesterol, mg/dL | 42 (34–51) | 41 (29–46) | 35 (31–42) | 0.236 |
| Triglycerides, mg/dL | 116 (79–156) | 104 (73–178) | 158 (112–243) | 0.139 |
| C-reactive protein, mg/L | 3.5 (1.6) | 3.7 (1.8) | 4.0 (1.9) | 0.339 |
| miRNAs3 | ||||
| miR-21 | 15.44 (4.40) | 14.55 (4.62) | 16.42 (5.09) | 0.454 |
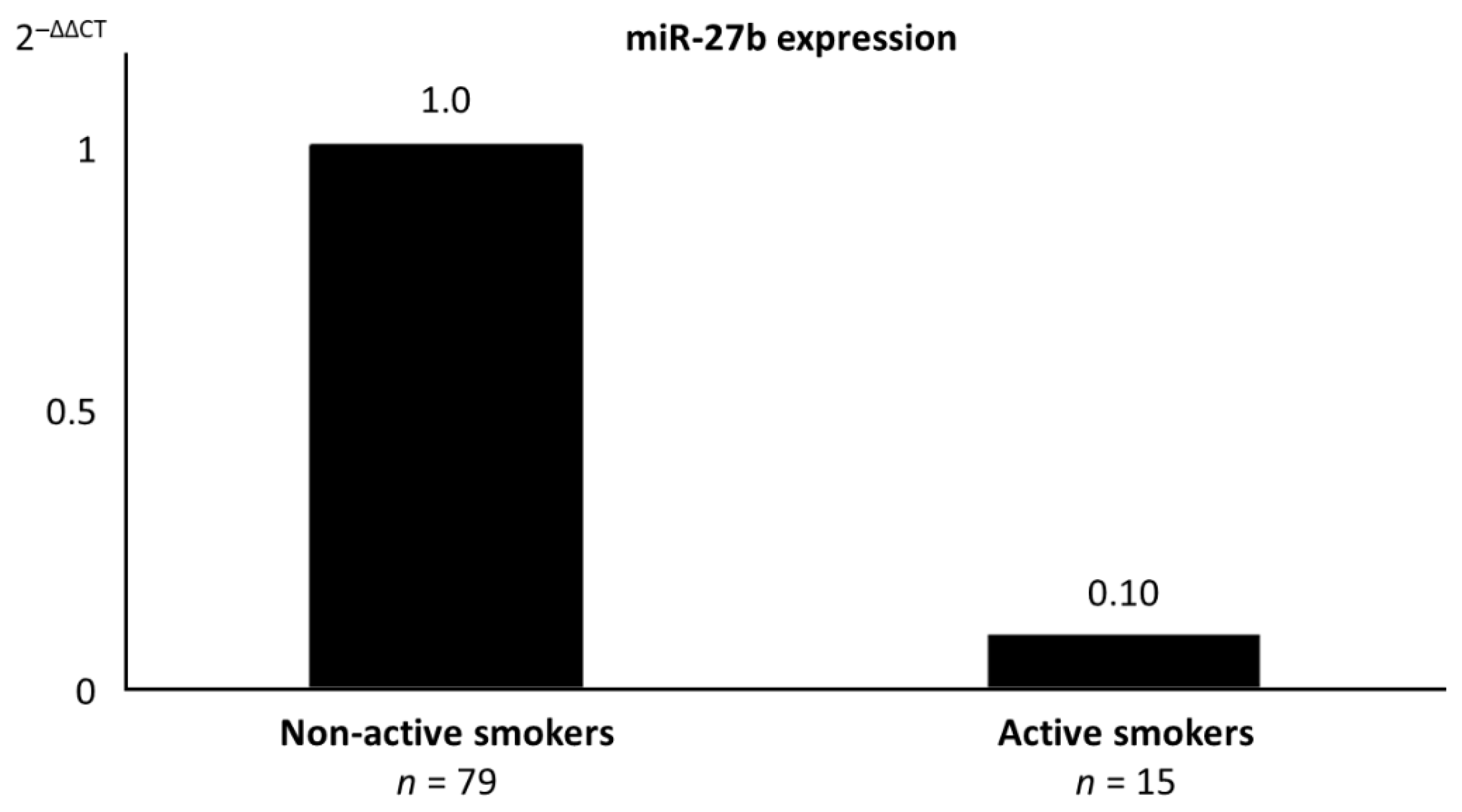
| miR-27b | 19.29 (4.42) | 17.44 (3.98) | 22.00 (4.35) | 0.014 2 |
| miR-29a | 21.01 (3.04) | 19.74 (3.86) | 23.19 (3.12) | 0.059 |
| miR-126 | 17.42 (15.20–23.98) | 16.10 (14.89–19.48) | 22.40 (17.65–24.15) | 0.075 |
| miR-146 | 20.03 (15.74–22.21) | 17.79 (16.26–20.12) | 21.69 (18.23–23.58) | 0.150 |
| miR-218 | 22.31 (15.67–23.02) | 22.31 (15.67–23.02) | 8.45 (−8.46 –21.50) | 0.120 |
| Independent Predictors | β | 95% CI | p-Value |
|---|---|---|---|
| Age, years | 1.11 | 1.04–1.18 | 0.002 |
| Cigarette smoking | 4.11 | 1.89–8.95 | 0.031 |
| Creatinine, mg/dL | 6.29 | 1.12–33.49 | <0.001 |
| Independent Predictors | β | 95% CI | p-Value |
|---|---|---|---|
| Leukocyte count | 2.33 | 1.37–3.94 | 0.002 |
| ΔCt miR-27b | 1.33 | 1.02–1.72 | 0.034 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira-da-Silva, T.; Napoleão, P.; Costa, M.C.; Gabriel, A.F.; Selas, M.; Silva, F.; Enguita, F.J.; Ferreira, R.C.; Carmo, M.M. Cigarette Smoking, miR-27b Downregulation, and Peripheral Artery Disease: Insights into the Mechanisms of Smoking Toxicity. J. Clin. Med. 2021, 10, 890. https://doi.org/10.3390/jcm10040890
Pereira-da-Silva T, Napoleão P, Costa MC, Gabriel AF, Selas M, Silva F, Enguita FJ, Ferreira RC, Carmo MM. Cigarette Smoking, miR-27b Downregulation, and Peripheral Artery Disease: Insights into the Mechanisms of Smoking Toxicity. Journal of Clinical Medicine. 2021; 10(4):890. https://doi.org/10.3390/jcm10040890
Chicago/Turabian StylePereira-da-Silva, Tiago, Patrícia Napoleão, Marina C. Costa, André F. Gabriel, Mafalda Selas, Filipa Silva, Francisco J. Enguita, Rui Cruz Ferreira, and Miguel Mota Carmo. 2021. "Cigarette Smoking, miR-27b Downregulation, and Peripheral Artery Disease: Insights into the Mechanisms of Smoking Toxicity" Journal of Clinical Medicine 10, no. 4: 890. https://doi.org/10.3390/jcm10040890