
Establishing Irreversible Electroporation Electric Field Potential Threshold in A Suspension In Vitro Model for Cardiac and Neuronal Cells

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Cell Culture and Culture Conditions
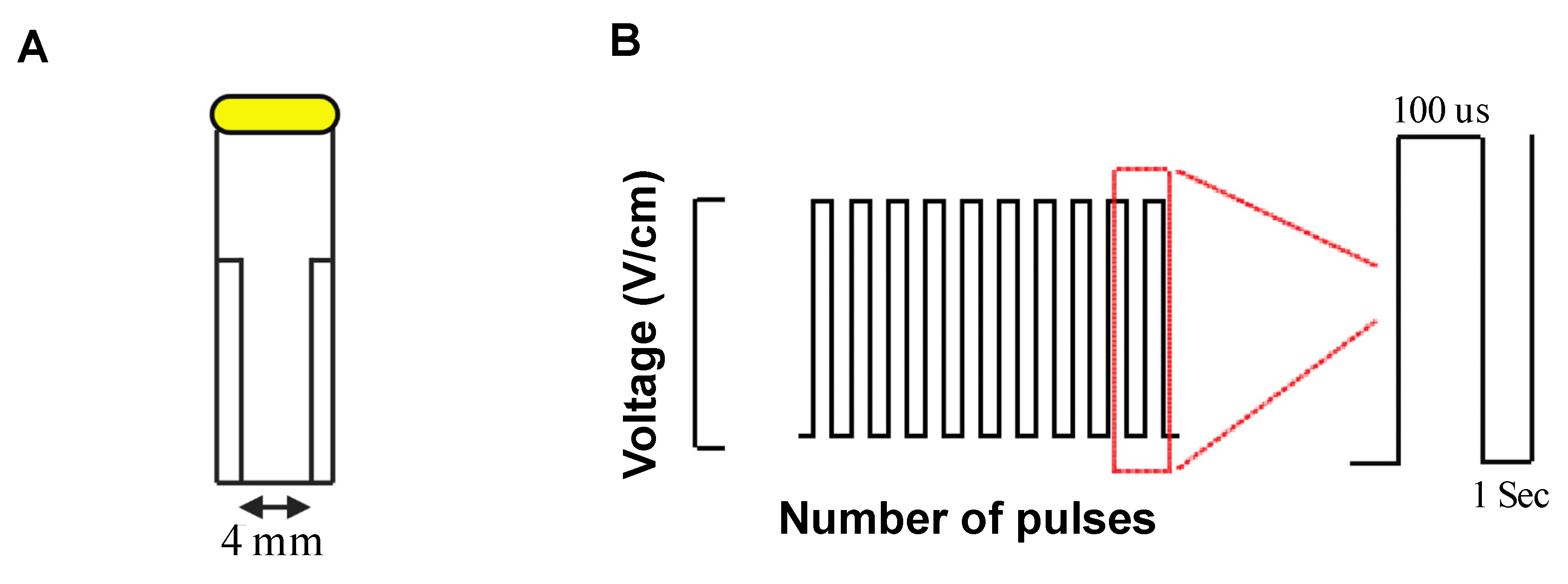
2.2. Electric Field Generation
2.3. Cell Viability Assay
2.4. Live–Dead Assay
2.5. Immunocytochemistry Staining
2.6. Statistical Analyses
3. Results
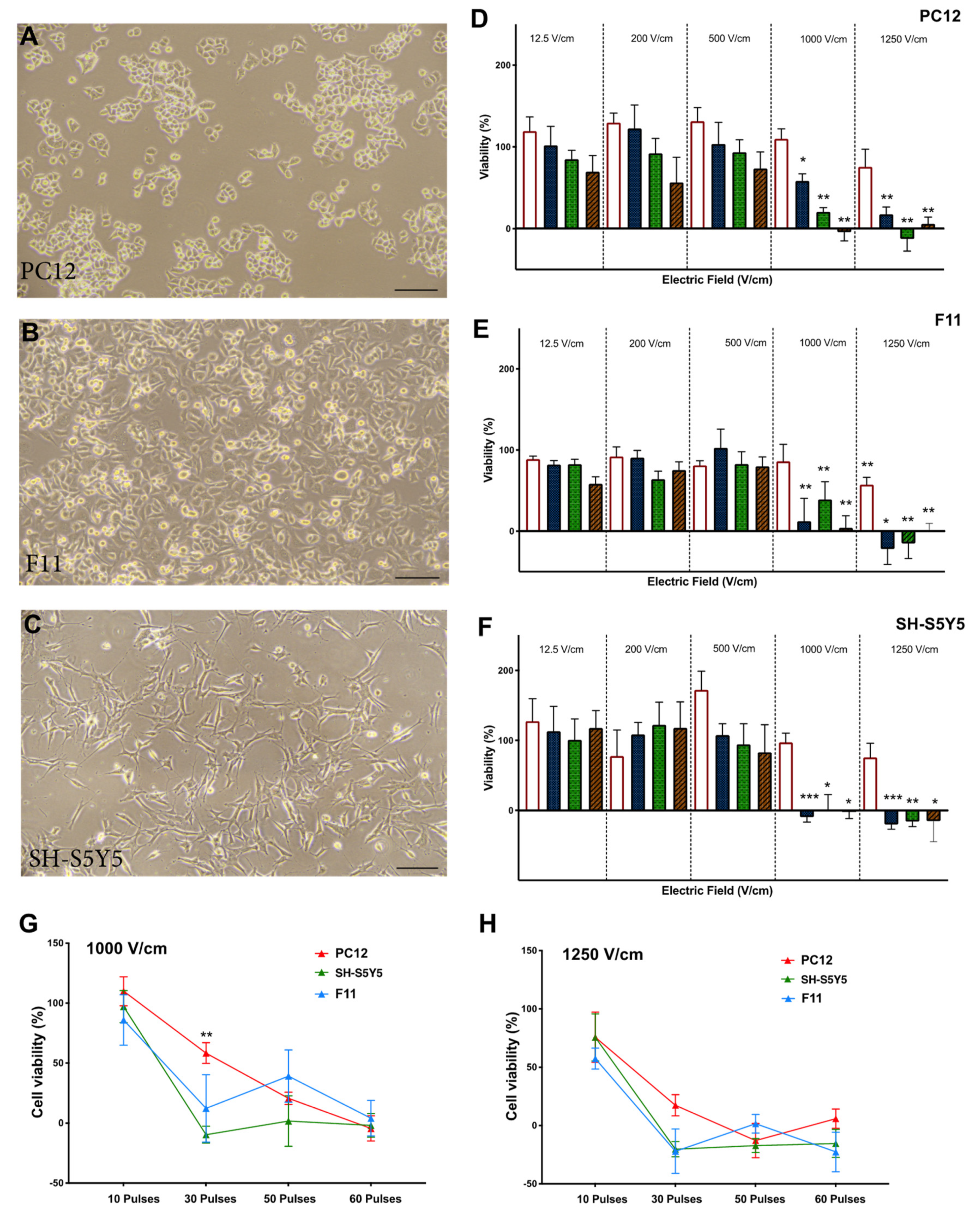
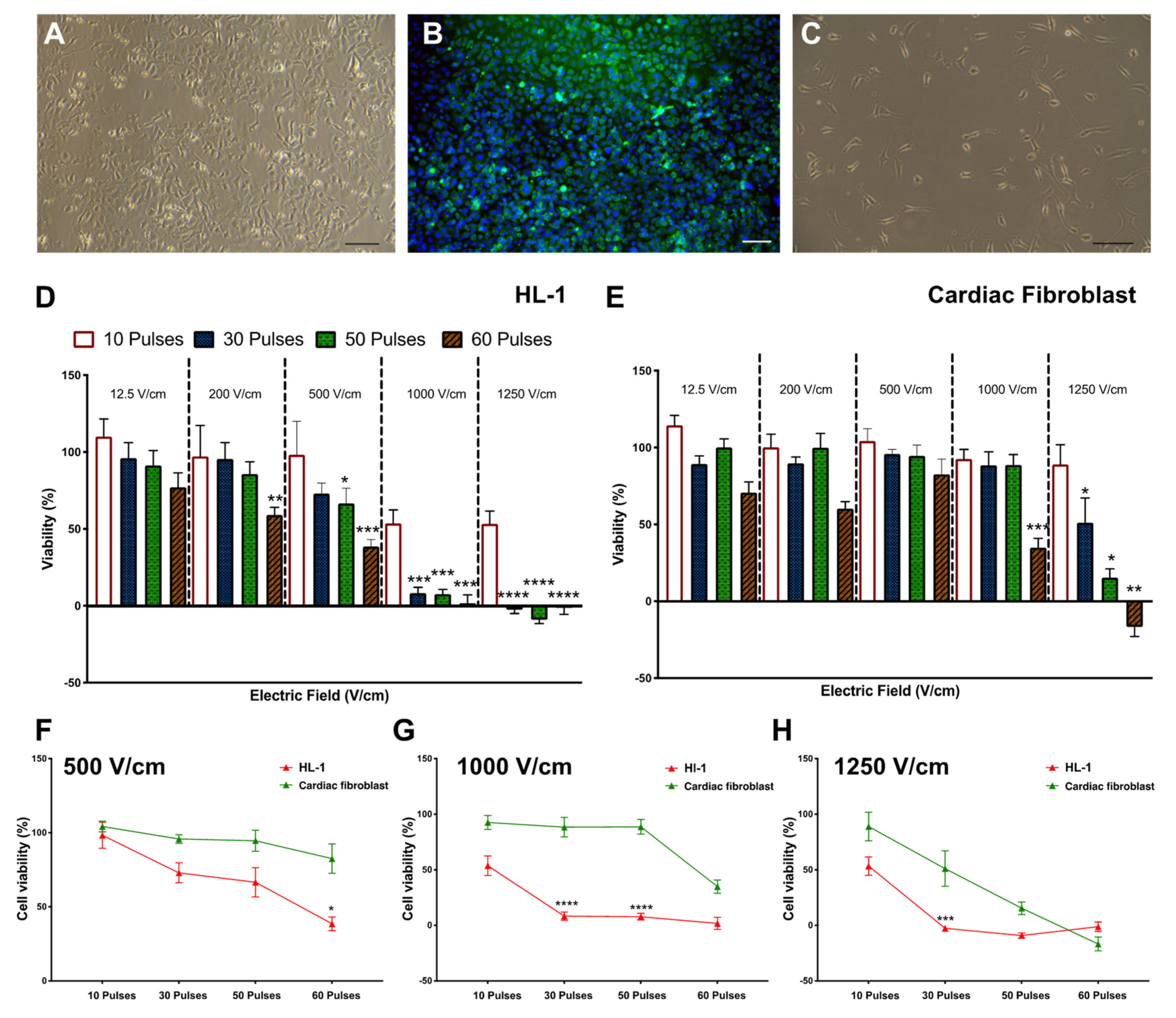
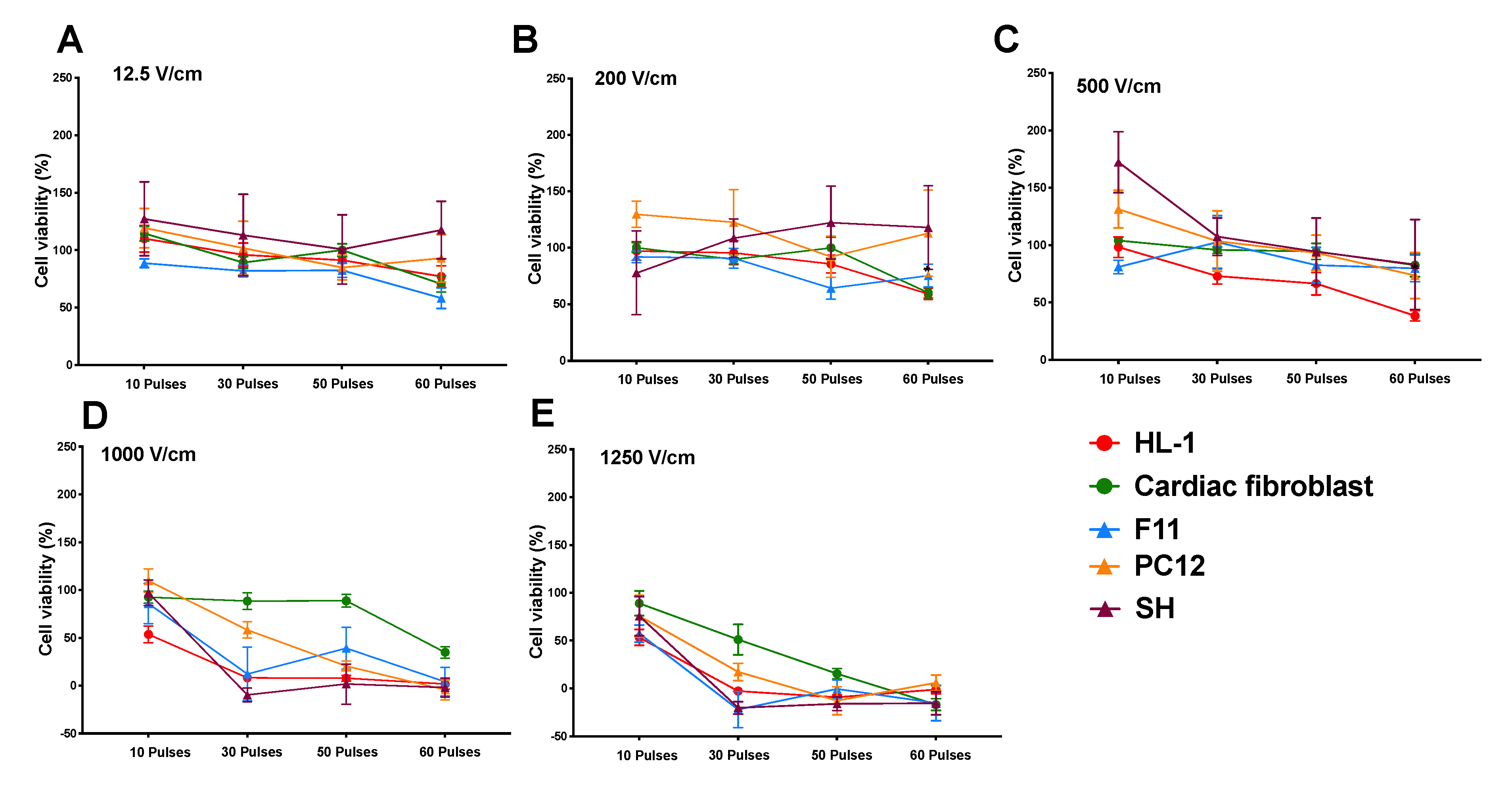
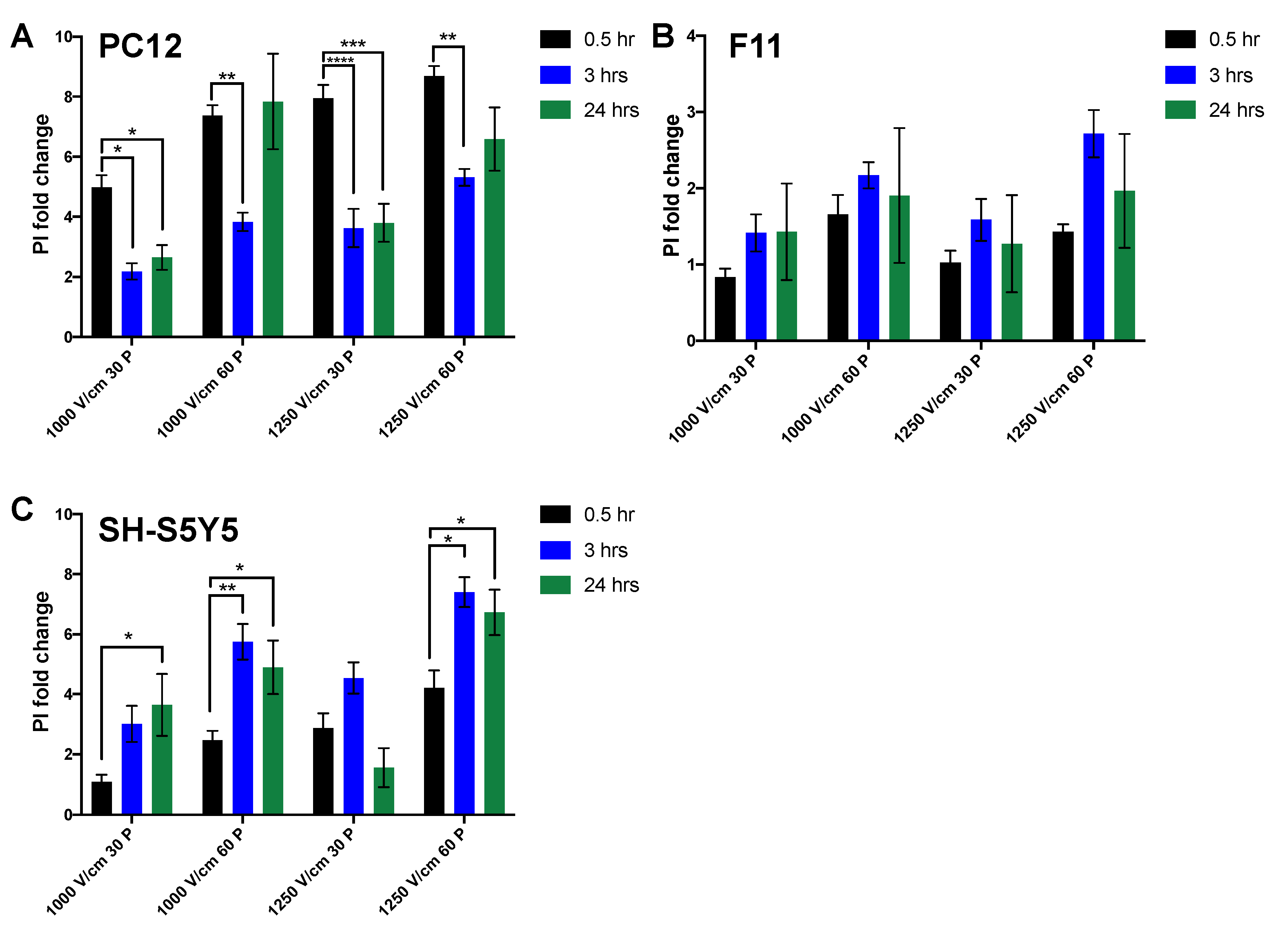
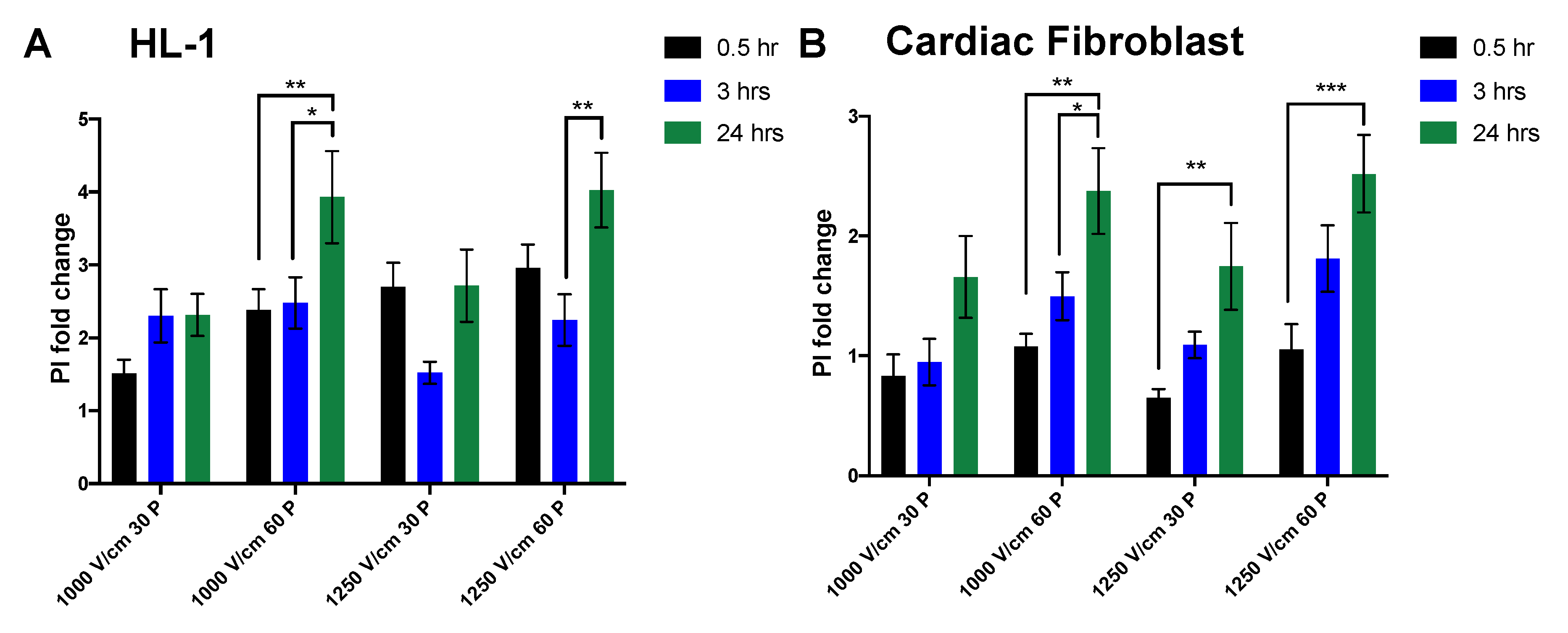
3.1. Reduction in Cell Viability Is Related to Electric Field Strength
3.2. Effect of Time on Cell Death Post Electroporation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tini, G.; Scagliola, R.; Monacelli, F.; La Malfa, G.; Porto, I.; Brunelli, C.; Rosa, G.M. Alzheimer’s Disease and Cardiovascular Disease: A Particular Association. Cardiol. Res. Pract. 2020, 2020, 2617970. [Google Scholar] [CrossRef] [PubMed]
- Frost, L.; Vestergaard, P.; Mosekilde, L. Hyperthyroidism and risk of atrial fibrillation or flutter: A population-based study. ACC Curr. J. Rev. 2004, 164, 1675–1678. [Google Scholar] [CrossRef] [Green Version]
- Chamberlain, A.M.; Agarwal, S.K.; Folsom, A.R.; Duval, S.; Soliman, E.Z.; Ambrose, M.; Eberly, L.; Alonso, A. Smoking and incidence of atrial fibrillation: Results from the Atherosclerosis Risk in Communities (ARIC) Study. Hear. Rhythm. 2011, 8, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlachos, K.; Mascia, G.; Martin, C.A.; Bazoukis, G.; Frontera, A.; Cheniti, G.; Letsas, K.P.; Efremidis, M.; Georgopoulos, S.; Gkalapis, C.; et al. Atrial fibrillation in Brugada syndrome: Current perspectives. J. Cardiovasc. Electrophysiol. 2020, 31, 975–984. [Google Scholar] [CrossRef] [PubMed]
- McBride, S.; Avazzadeh, S.; Wheatley, A.; O’Brien, B.; Coffey, K.; Elahi, A.; O’Halloran, M.; Quinlan, L. Ablation Modalities for Therapeutic Intervention in Arrhythmia-Related Cardiovascular Disease: Focus on Electroporation. J. Clin. Med. 2021, 10, 2657. [Google Scholar] [CrossRef] [PubMed]
- Guenther, E.; Klein, N.; Zapf, S.; Weil, S.; Schlosser, C.; Rubinsky, B.; Stehling, M.K. Prostate cancer treatment with Irreversible Electroporation (IRE): Safety, efficacy and clinical experience in 471 treatments. PLoS ONE 2019, 14, e0215093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charpentier, K.P.; Wolf, F.; Noble, L.; Winn, B.; Resnick, M.; Dupuy, D. Irreversible electroporation of the pancreas in swine: A pilot study. HPB 2010, 12, 348–351. [Google Scholar] [CrossRef] [Green Version]
- Rubinsky, B.; Onik, G.; Mikus, P. Irreversible Electroporation: A New Ablation Modality—Clinical Implications. Technol. Cancer Res. Treat. 2007, 6, 37–48. [Google Scholar] [CrossRef]
- Edd, J.F.; Horowitz, L.; Dávalos, R.; Mir, L.; Rubinsky, B. In Vivo Results of a New Focal Tissue Ablation Technique: Irreversible Electroporation. IEEE Trans. Biomed. Eng. 2006, 53, 1409–1415. [Google Scholar] [CrossRef]
- Lavee, J.; Onik, G.; Mikus, P.; Rubinsky, B. A Novel Nonthermal Energy Source for Surgical Epicardial Atrial Ablation: Irreversible Electroporation. Hear. Surg. Forum 2007, 10, E162–E167. [Google Scholar] [CrossRef]
- Reddy, V.Y.; Neuzil, P.; Koruth, J.; Petru, J.; Funosako, M.; Cochet, H.; Sediva, L.; Chovanec, M.; Dukkipati, S.R.; Jais, P. Pulsed Field Ablation for Pulmonary Vein Isolation in Atrial Fibrillation. J. Am. Coll. Cardiol. 2019, 74, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J. Electroporation of cells and tissues. IEEE Trans. Plasma Sci. 2000, 28, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Kaminska, I.; Kotulska, M.; Stecka, A.; Saczko, J.; Drag-Zalesinska, M.; Wysocka, T.; Choromanska, A.; Skolucka, N.; Nowicki, R.; Marczak, J.; et al. Electroporation-induced changes in normal immature rat myoblasts (H9C2). Gen. Physiol. Biophys. 2012, 31, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.W.; Kostecki, G.; Fish, J.M.; Jensen, J.A.; Tandri, H. In vitro cell selectivity of reversible and irreversible: Electroporation in cardiac tissue. Circ. Arrhythm. Electrophysiol. 2021, 14, e008817. [Google Scholar] [CrossRef]
- Sugrue, A.; Maor, E.; Ivorra, A.; Vaidya, V.; Witt, C.; Kapa, S.; Asirvatham, S. Irreversible electroporation for the treatment of cardiac arrhythmias. Expert Rev. Cardiovasc. Ther. 2018, 16, 349–360. [Google Scholar] [CrossRef]
- Avazzadeh, S.; McBride, S.; O’Brien, B.; Coffey, K.; Elahi, A.; O’Halloran, M.; Soo, A.; Quinlan, L.R. Ganglionated Plexi Ablation for the Treatment of Atrial Fibrillation. J. Clin. Med. 2020, 9, 3081. [Google Scholar] [CrossRef]
- Pani, A.; Giuseppina, B.; Bonanno, C.; Bongiorni, M.G.; Bottoni, N.; Brambilla, R.; De Ceglia, S.; Della Bella, P.; De Vito, G.; Malaspina, D.; et al. Predictors of Zero X-Ray Ablation for Supraventricular Tachycardias in a Nationwide Multicenter Experience. Circ. Arrhythmia Electrophysiol. 2018, 11, e005592. [Google Scholar] [CrossRef]
- Yang, L.; Sun, G.; Chen, X.; Chen, G.; Yang, S.; Guo, P.; Wang, Y.; Wang, D.W. Meta-Analysis of Zero or Near-Zero Fluoroscopy Use During Ablation of Cardiac Arrhythmias. Am. J. Cardiol. 2016, 118, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Aksu, T.; Yalin, K.; Gopinathannair, R. Fractionation mapping software to map ganglionated plexus sites during sinus rhythm. J. Cardiovasc. Electrophysiol. 2020, 31, 3326–3329. [Google Scholar] [CrossRef]
- Maor, E.; Ivorra, A.; Rubinsky, B. Non Thermal Irreversible Electroporation: Novel Technology for Vascular Smooth Muscle Cells Ablation. PLoS ONE 2009, 4, e4757. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Fan, Q.; Ji, Z.; Qiu, X.; Li, Z. The Effects of Irreversible Electroporation (IRE) on Nerves. PLoS ONE 2011, 6, e18831. [Google Scholar] [CrossRef] [PubMed]
- van Driel, V.J.H.M.; Neven, K.; van Wessel, H.; Vink, A.; Doevendans, P.A.F.M.; Wittkampf, F.H.M. Low vulnerability of the right phrenic nerve to electroporation ablation. Heart Rhythm. 2015, 12, 1838–1844. [Google Scholar] [CrossRef] [PubMed]
- Neven, K.; van Driel, V.; van Wessel, H.; van Es, R.; du Pré, B.; Doevendans, P.A.; Wittkampf, F. Safety and feasibility of closed chest epicardial catheter ablation using electroporation. Circ. Arrhythm. Electrophysiol. 2014, 7, 913–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loh, P.; van Es, R.; Groen, M.H.A.; Neven, K.; Kassenberg, W.; Wittkampf, F.H.M.; Doevendans, P.A. Pulmonary vein isolation with single pulse irreversible electroporation: A first in human study in 10 patients with atrial fibrillation. Circ. Arrhythm. Electrophysiol. 2020, 13, e008192. [Google Scholar] [CrossRef]
- Davalos, R.V.; Mir, I.L.M.; Rubinsky, B. Tissue ablation with irreversible electroporation. Ann. Biomed. Eng. 2005, 33, 223–231. [Google Scholar] [CrossRef]
- Qin, Q.; Xiong, Z.-A.; Liu, Y.; Yao, C.-G.; Zhou, W.; Hua, Y.-Y.; Wang, Z.-L. Effects of irreversible electroporation on cervical cancer cell lines in vitro. Mol. Med. Rep. 2016, 14, 2187–2193. [Google Scholar] [CrossRef] [Green Version]
- Mercadal, B.; Beitel-White, N.; Aycock, K.N.; Castellví, Q.; Davalos, R.V.; Ivorra, A. Dynamics of Cell Death After Conventional IRE and H-FIRE Treatments. Ann. Biomed. Eng. 2020, 48, 1451–1462. [Google Scholar] [CrossRef] [Green Version]
- Dm, D.P.; Naksuk, N.; Killu, A.K.; Kapa, S.; Witt, C.; Sugrue, A.; DeSimone, C.V.; Madhavan, M.; Groot, J.R.; O’Brien, B.; et al. Electroporation of epicardial autonomic ganglia: Safety and efficacy in medium-term canine models. J. Cardiovasc. Electrophysiol. 2019, 30, 607–615. [Google Scholar] [CrossRef]
- van Driel, V.J.; Neven, K.G.; van Wessel, H.; du Pré, B.C.; Vink, A.; Doevendans, P.A.; Wittkampf, F.H. Pulmonary vein stenosis after catheter ablation: Electroporation versus radiofrequency. Circ. Arrhythm. Electrophysiol. 2014, 7, 734–738. [Google Scholar] [CrossRef] [Green Version]
- Wittkampf, F.H.; van Es, R.; Neven, K. Electroporation and its Relevance for Cardiac Catheter Ablation. JACC: Clin. Electrophysiol. 2018, 4, 977–986. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1000 V/Cm 30 Pulses | Mean ± SEM 1 | Mean ± SEM 2 | p-Value |
|---|---|---|---|
| F11 vs. PC12 | 0.84 ± 0.11 | 4.98 ± 0.41 | <0.0001 |
| F11 vs. SH-S5Y5 | 0.84 ± 0.11 | 1.08 ± 0.23 | 0.87 (ns) |
| PC12 vs. SH-S5Y5 | 4.98 ± 0.41 | 1.08 ± 0.23 | <0.0001 |
| 1000 V/cm 60 pulses | |||
| F11 vs. PC12 | 1.66 ± 0.25 | 7.37 ± 0.34 | <0.0001 |
| F11 vs. SH-S5Y5 | 1.66 ± 0.25 | 2.47 ± 0.32 | 0.16 (ns) |
| PC12 vs. SH-S5Y5 | 7.37 ± 0.34 | 2.47 ± 0.32 | <0.0001 |
| 1250 V/cm 30 pulses | |||
| F11 vs. PC12 | 1.03 ± 1.16 | 7.95 ± 0.44 | <0.0001 |
| F11 vs. SH-S5Y5 | 1.03 ± 1.16 | 2.88 ± 0.49 | 0.0010 |
| PC12 vs. SH-S5Y5 | 7.95 ± 0.44 | 2.88 ± 0.49 | <0.0001 |
| 1250 V/cm 60 pulses | |||
| F11 vs. PC12 | 1.43 ± 0.10 | 8.69 ± 0.34 | <0.0001 |
| F11 vs. SH-S5Y5 | 1.43 ± 0.10 | 4.22 ± 0.57 | <0.0001 |
| PC12 vs. SH-S5Y5 | 8.69 ± 0.34 | 4.22 ± 0.57 | <0.0001 |
| 1000 V/cm 30 pulses | Mean ± SEM 1 | Mean ± SEM 2 | p Value |
|---|---|---|---|
| HL-1 vs. Cardiac fibroblast | 1.51 ± 0.60 | 0.83 ± 0.44 | 0.70 [ns] |
| 1000 V/cm 60 pulses | |||
| HL-1 vs. Cardiac fibroblast | 2.38 ± 0.89 | 1.08 ± 0.26 | 0.06 [ns] |
| 1250 V/cm 30 pulses | |||
| HL-1 vs. Cardiac fibroblast | 2.70 ± 0.97 | 0.65 ± 0.18 | 0.0004 |
| 1250 V/cm 60 pulses | |||
| HL-1 vs. Cardiac fibroblast | 2.96 ± 0.95 | 1.05 ± 0.52 | 0.0014 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avazzadeh, S.; O’Brien, B.; Coffey, K.; O’Halloran, M.; Keane, D.; Quinlan, L.R. Establishing Irreversible Electroporation Electric Field Potential Threshold in A Suspension In Vitro Model for Cardiac and Neuronal Cells. J. Clin. Med. 2021, 10, 5443. https://doi.org/10.3390/jcm10225443
Avazzadeh S, O’Brien B, Coffey K, O’Halloran M, Keane D, Quinlan LR. Establishing Irreversible Electroporation Electric Field Potential Threshold in A Suspension In Vitro Model for Cardiac and Neuronal Cells. Journal of Clinical Medicine. 2021; 10(22):5443. https://doi.org/10.3390/jcm10225443
Chicago/Turabian StyleAvazzadeh, Sahar, Barry O’Brien, Ken Coffey, Martin O’Halloran, David Keane, and Leo R. Quinlan. 2021. "Establishing Irreversible Electroporation Electric Field Potential Threshold in A Suspension In Vitro Model for Cardiac and Neuronal Cells" Journal of Clinical Medicine 10, no. 22: 5443. https://doi.org/10.3390/jcm10225443