
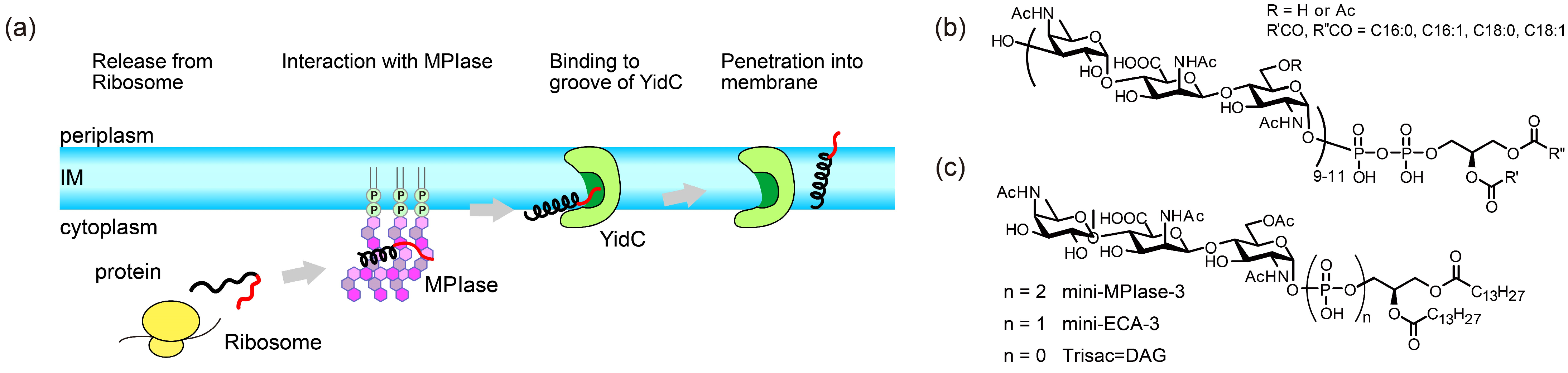
Roles of a Glycolipid MPIase in Sec-Independent Membrane Protein Insertion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Factors Influencing Membrane Protein Insertion
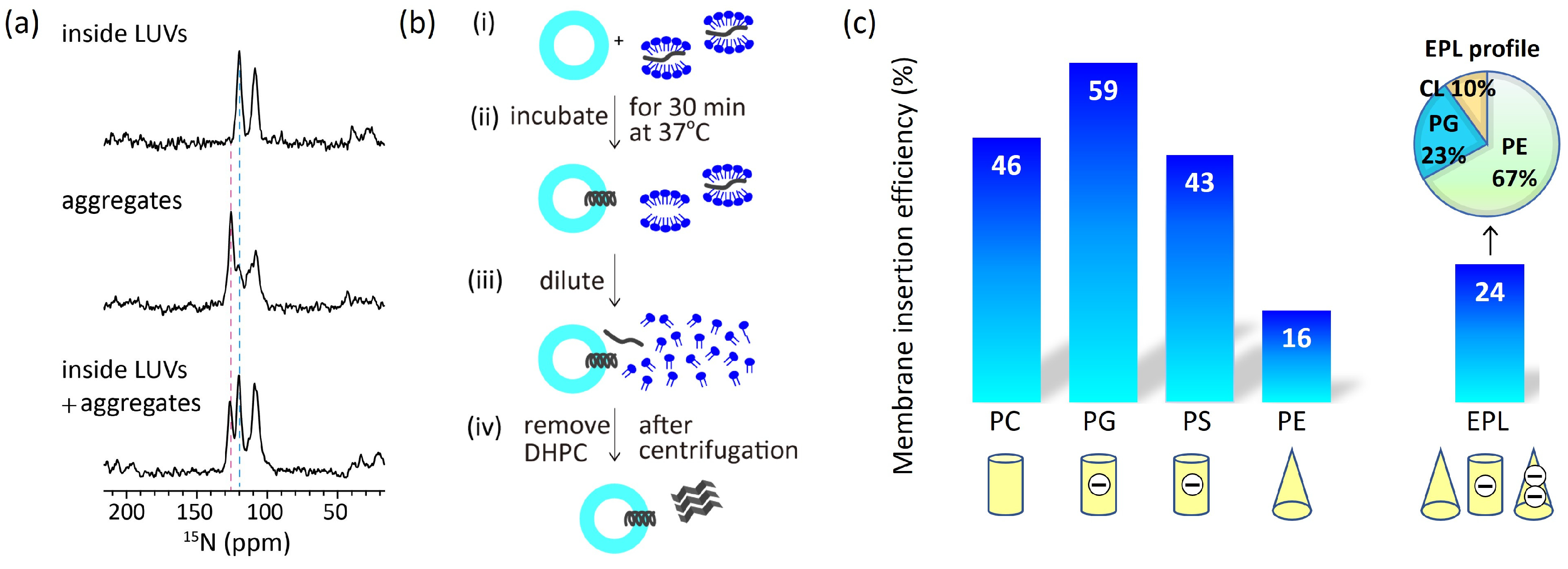
2.1. Charges of Membrane Lipids
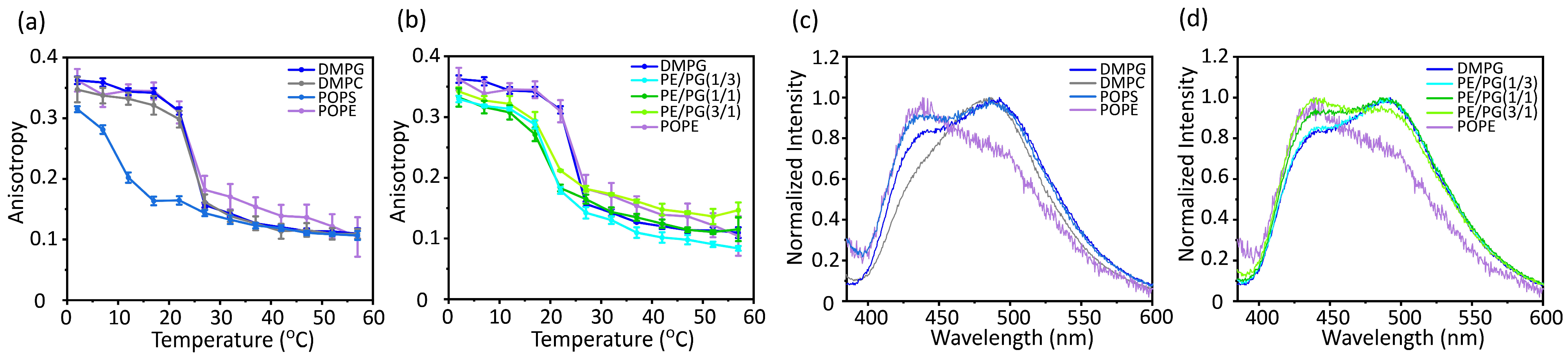
2.2. Acyl Chain Ordering
2.3. Spontaneous Lipid Curvature
2.4. Membrane Defects
2.5. Conformational Changes of Proteins Associated with Membranes
3. Effect of Membrane Properties on Membrane Insertion
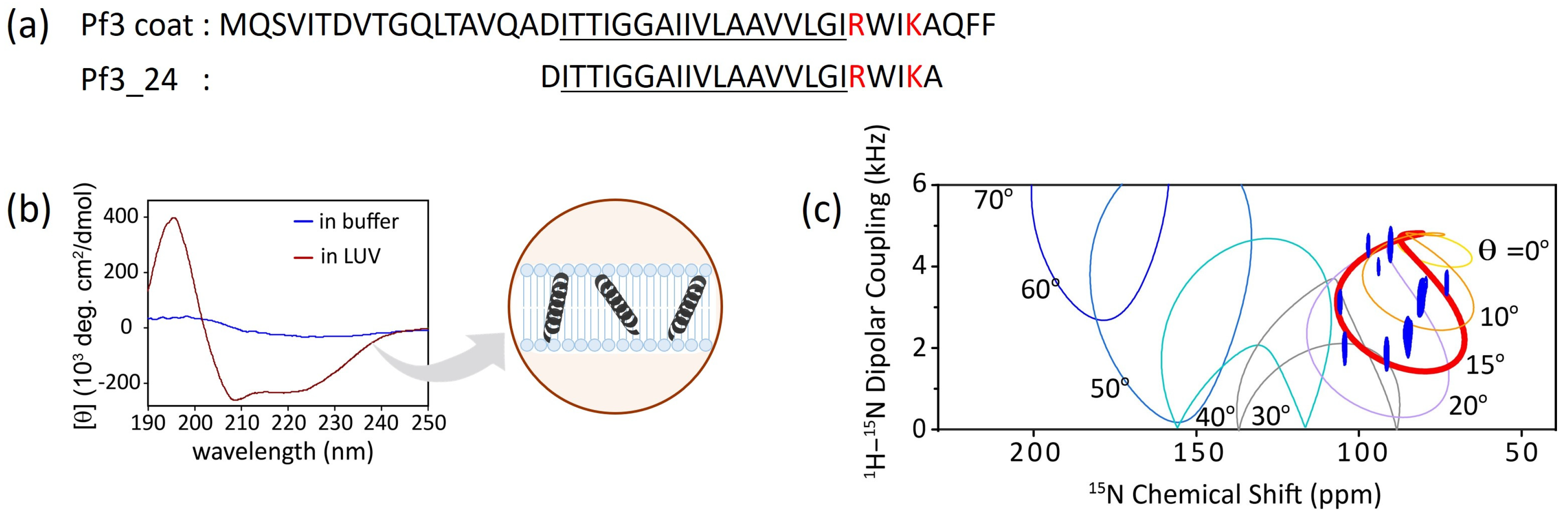
3.1. Conformational Changes and Topology of Pf3_24 in Membranes
3.2. Insertion of Pf3_24 into Membranes
3.3. Acyl Chain Mobility and Membrane Surface Packing
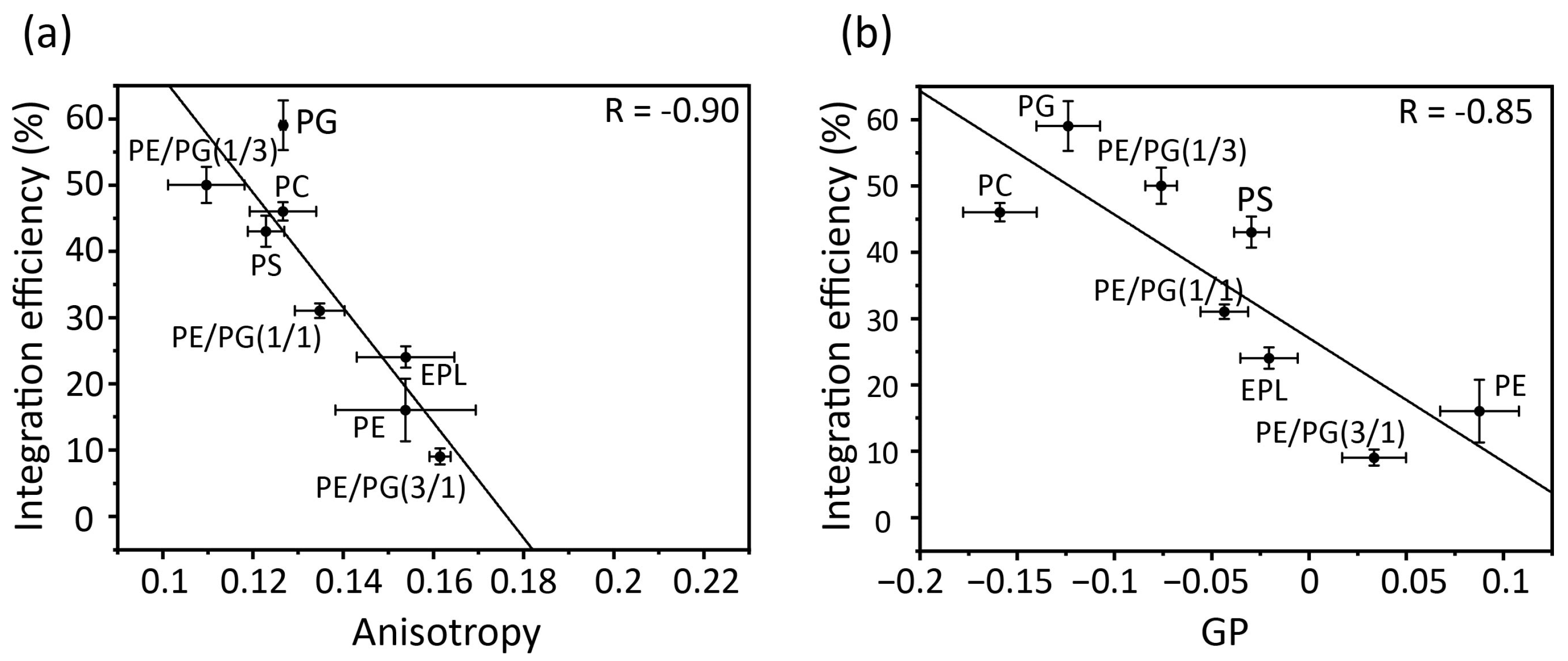
3.4. Correlation between Membrane Physicochemical Properties and Membrane Insertion Efficiency of Pf3_24
4. Role of MPIase on Membrane Protein Insertion
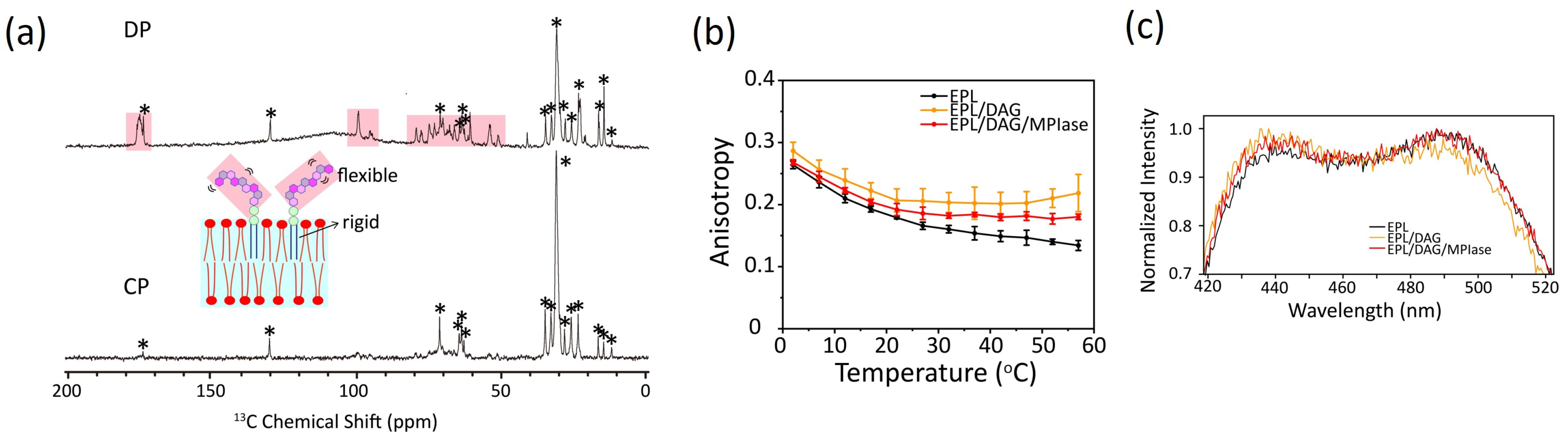
4.1. Mobility of MPIase in Membranes
4.2. Effects on Flip–Flop Motion of DAG
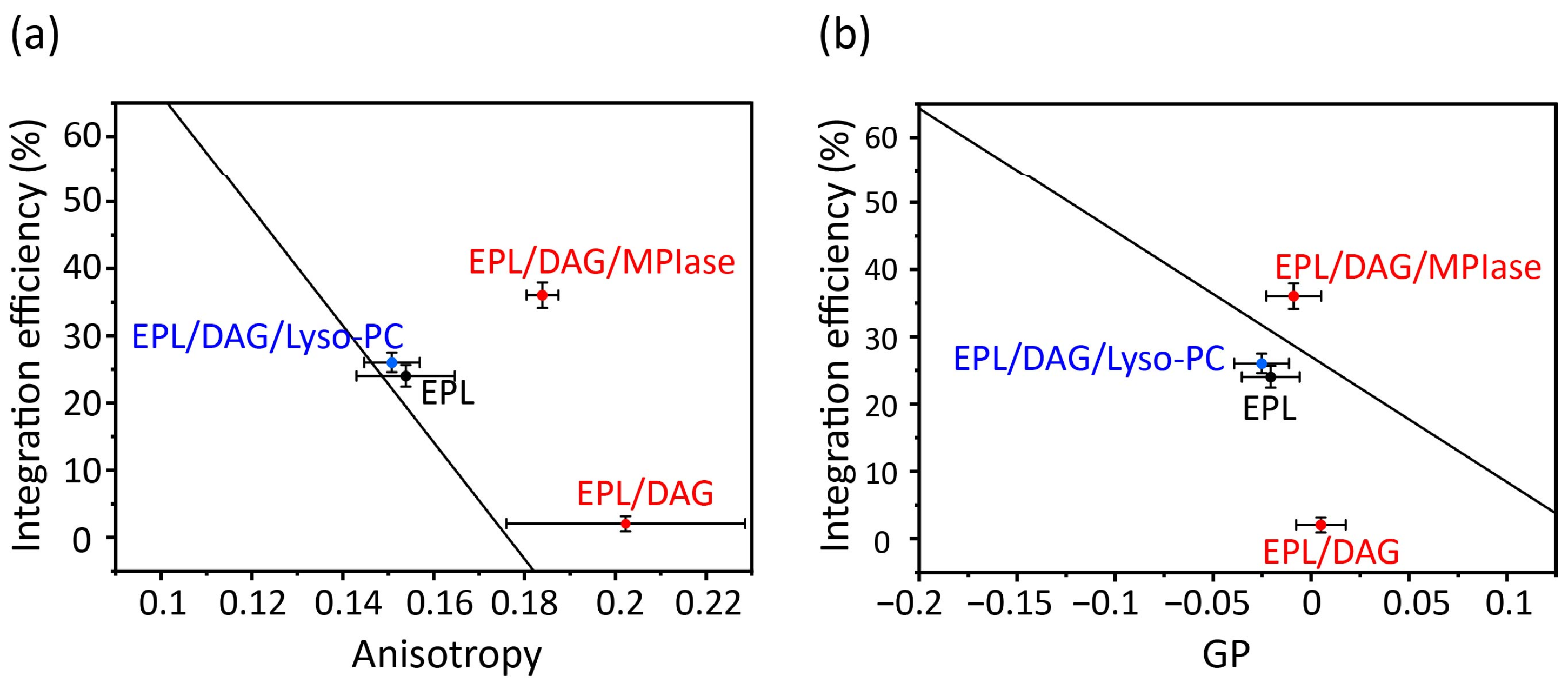
4.3. Effects of DAG and MPIase on Acyl Chain Mobility and Membrane Surface Packing
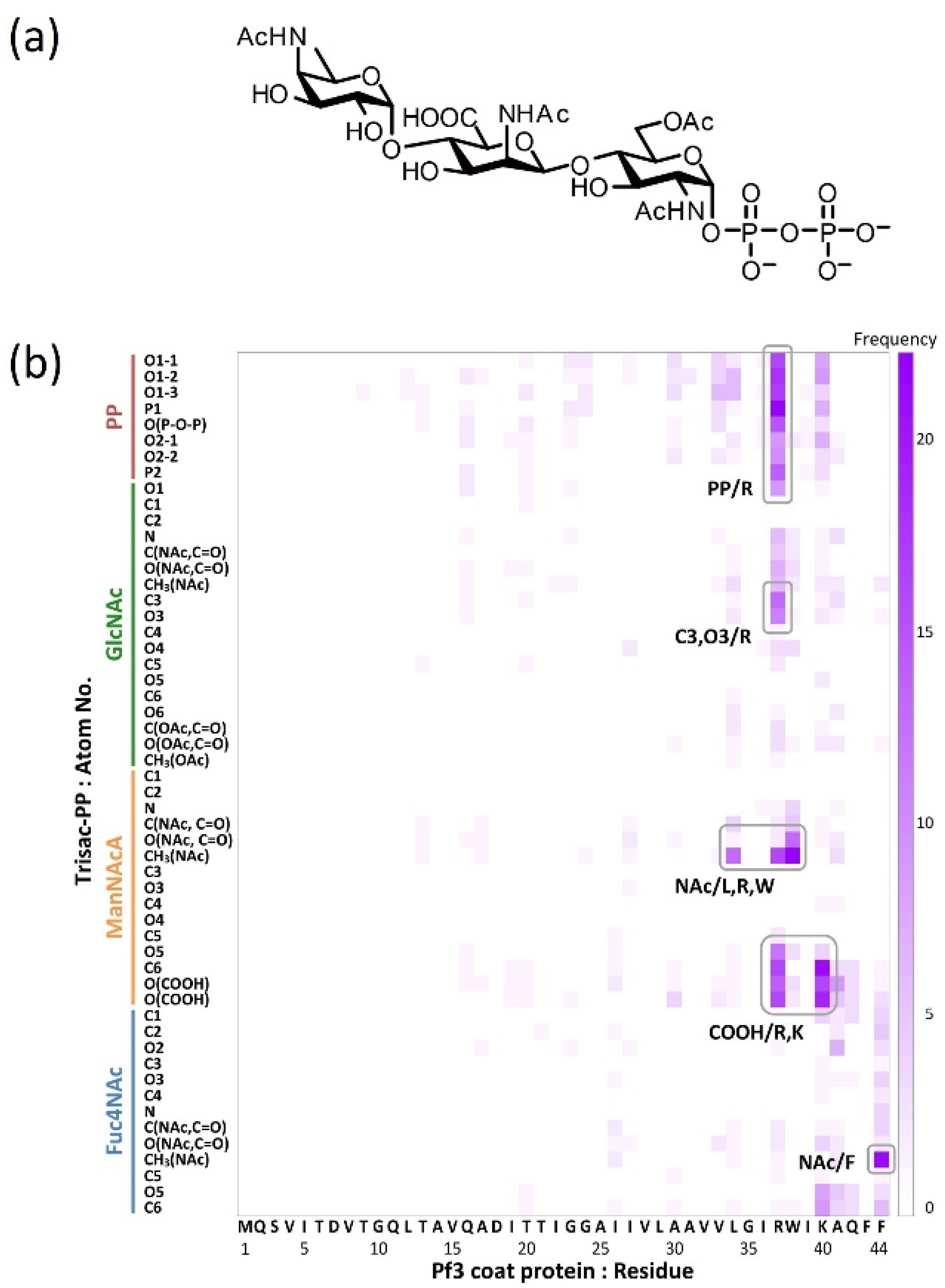
4.4. Identification of the Contact Sites by Using Docking Simulations
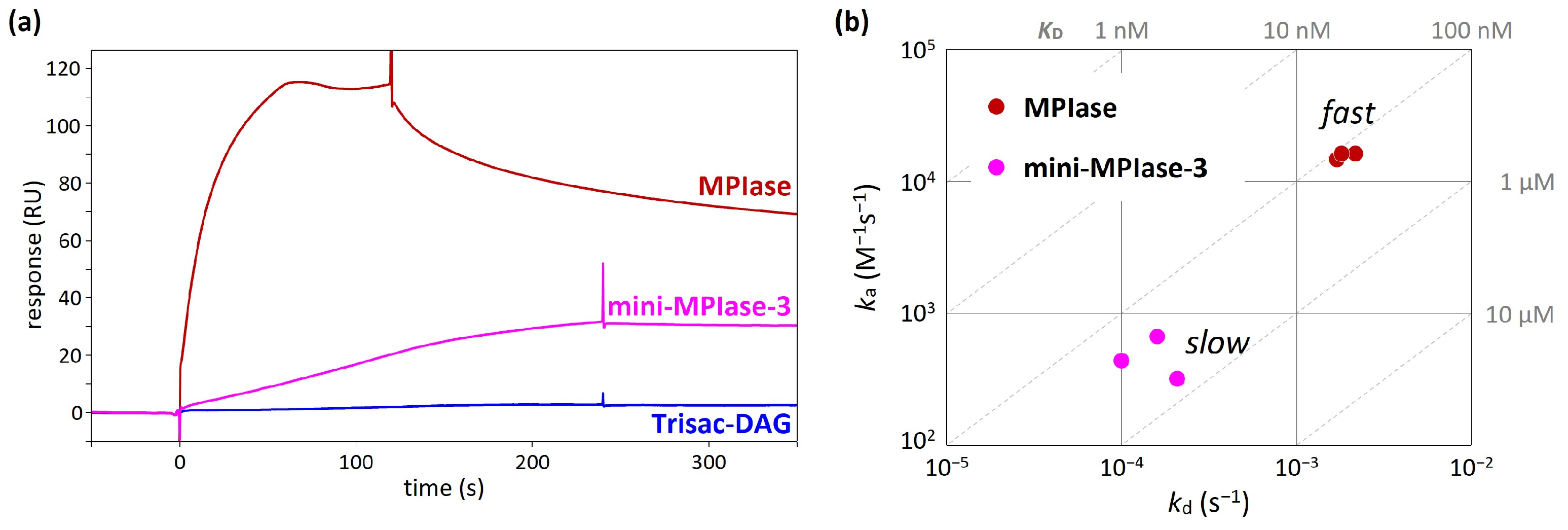
4.5. Binding Kinetics and Affinity of MPIase−Pf3 Coat Interactions
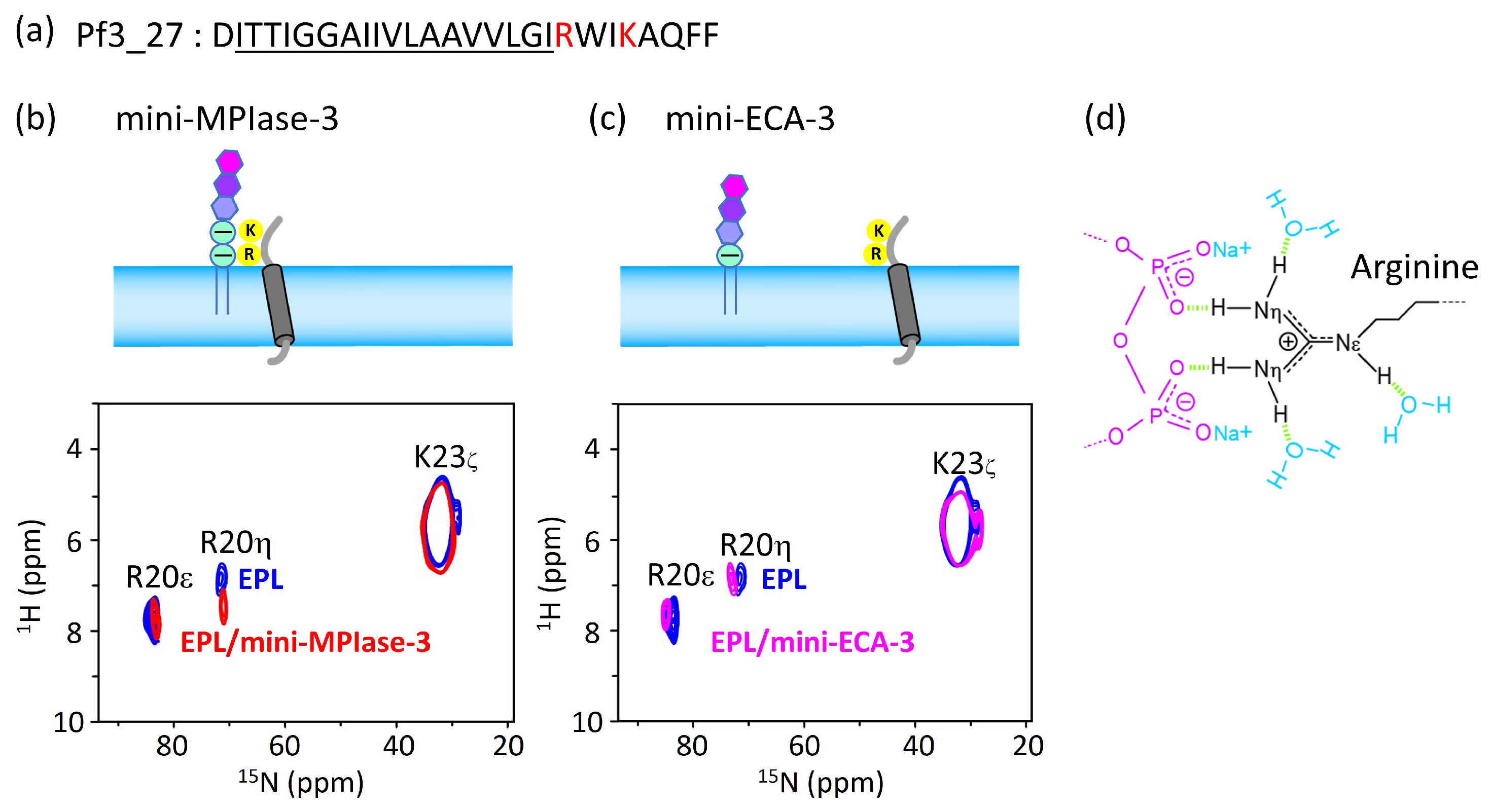
4.6. Interaction of the MPIase Pyrophosphate Group with Basic Residues in Pf3_27
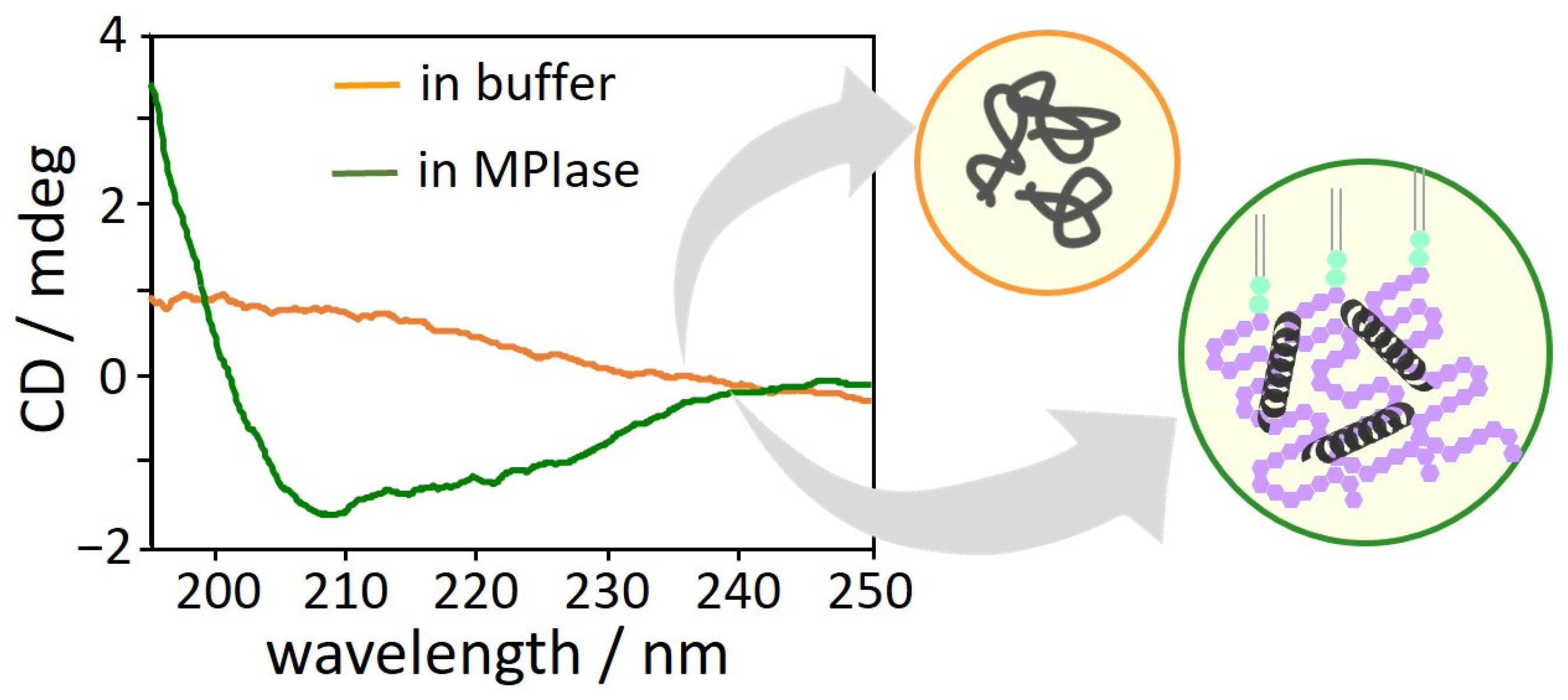
4.7. Inhibition of Protein Aggregation by MPIase
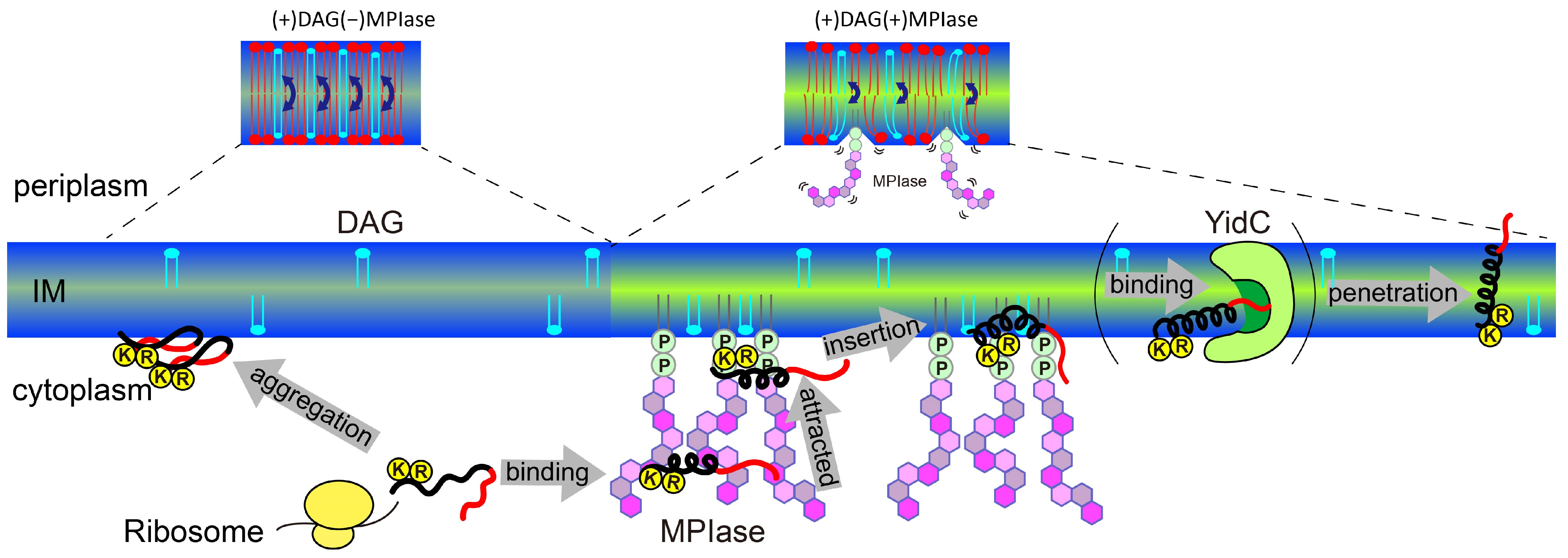
5. Mechanism of Sec-Independent Membrane Protein Insertion by MPIase
- DAG blocks spontaneous insertion by reducing acyl chain mobility in the membrane core. The rapid flip–flop of DAG also reduces lateral diffusion and fills membrane defects, exposing acyl chains and making it difficult for proteins to contact the inside of the membranes. However, the presence of MPIase cancels out the effect of DAG and restores mobility. The flexible bulky sugar chain of MPIase makes the membrane surface flexible, increases the mobility of the membrane core region, and reduces the flip–flop motion of DAG. Small hydrophobic membrane proteins preferentially associate with the membrane core region rather than with the aqueous environment outside the membrane.
- After a protein is released from a ribosome, the long sugar chain of MPIase captures the protein and prevents its aggregation. The protein efficiently changes its conformation through rapid association with and dissociation from the flexible long sugar chains of MPIase.
- The basic residues that generally exist in the cytoplasmic loops near the hydrophobic transmembrane regions of the protein are attracted towards the membrane surface through electrostatic interactions with the pyrophosphate of MPIase.
- The protein easily gains access to the loosened membrane core through hydrophobic interactions and finally integrates into the membrane.
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| membrane protein integrase | MPIase |
| Escherichia coli | E. coli |
| diacylglycerol | DAG |
| E. coli phospholipids | EPLs |
| phosphatidylethanolamine | PE |
| phosphatidylglycerol | PG |
| cardiolipin | CL |
| antimicrobial peptides | AMPs |
| phosphatidylcholine | PC |
| phosphatidylserine | PS |
| 1,2-dimyristoyl-sn-glycero-3-phosphocholine | DMPC |
| 1,2-diheptanoyl-sn-glycero-3-phosphocholine | DHPC |
| large unilamellar vesicles | LUV |
| cross-polarization under magic angle spinning | CPMAS |
| 1,6-diphenyl-1,3,5-hexatriene | DPH |
| generalized polarization | GP |
| 1,2-dimyristoyl-sn-glycero-3-phospho-(1′-rac-glycerol) | DMPG |
| 1-palmitoyl-2-oleoyl-sn-glycero-3-phospho-L-serine | POPS |
| 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoethanolamine | POPE |
| magic angle spinning | MAS |
| surface plasmon resonance | SPR |
| association rate | ka |
| dissociation rate | kd |
| circular dichroism | CD |
References
- Luirink, J.; von Heijne, G.; Houben, E.; de Gier, J.W. Biogenesis of inner membrane proteins in Escherichia coli. Annu. Rev. Microbiol. 2005, 59, 329–355. [Google Scholar] [CrossRef]
- Wolfe, P.B.; Rice, M.; Wickner, W. Effects of two sec genes on protein assembly into the plasma membrane of Escherichia coli. J. Biol. Chem. 1985, 260, 1836–1841. [Google Scholar] [CrossRef] [PubMed]
- Geller, B.L.; Wickner, W. M13 procoat inserts into liposomes in the absence of other membrane proteins. J. Biol. Chem. 1985, 260, 13281–13285. [Google Scholar] [CrossRef]
- Kiefer, D.; Hu, X.; Dalbey, R.; Kuhn, A. Negatively charged amino acid residues play an active role in orienting the Sec-independent Pf3 coat protein in the Escherichia coli inner membrane. EMBO J. 1997, 16, 2197–2204. [Google Scholar] [CrossRef] [PubMed]
- Mariappan, M.; Li, X.; Stefanovic, S.; Sharma, A.; Mateja, A.; Keenan, R.J.; Hegde, R.S. A ribosome-associating factor chaperones tail-anchored membrane proteins. Nature 2010, 466, 1120–1124. [Google Scholar] [CrossRef] [PubMed]
- Mariappan, M.; Mateja, A.; Dobosz, M.; Bove, E.; Hegde, R.S.; Keenan, R.J. The mechanism of membrane-associated steps in tail-anchored protein insertion. Nature 2011, 477, 61–66. [Google Scholar] [CrossRef]
- Nishiyama, K.; Ikegami, A.; Moser, M.; Schiltz, E.; Tokuda, H.; Muller, M. A derivative of lipid A is involved in signal recognition particle/SecYEG-dependent and -independent membrane integrations. J. Biol. Chem. 2006, 281, 35667–35676. [Google Scholar] [CrossRef]
- Kawashima, Y.; Miyazaki, E.; Müller, M.; Tokuda, H.; Nishiyama, K.-I. Diacylglycerol Specifically Blocks Spontaneous Integration of Membrane Proteins and Allows Detection of a Factor-assisted Integration. J. Biol. Chem. 2008, 283, 24489–24496. [Google Scholar] [CrossRef]
- Nishiyama, K.; Maeda, M.; Abe, M.; Kanamori, T.; Shimamoto, K.; Kusumoto, S.; Ueda, T.; Tokuda, H. A novel complete reconstitution system for membrane integration of the simplest membrane protein. Biochem. Biophys. Res. Commun. 2010, 394, 733–736. [Google Scholar] [CrossRef]
- Nishiyama, K.-I.; Maeda, M.; Yanagisawa, K.; Nagase, R.; Komura, H.; Iwashita, T.; Yamagaki, T.; Kusumoto, S.; Tokuda, H.; Shimamoto, K. MPIase is a glycolipozyme essential for membrane protein integration. Nat. Commun. 2012, 3, 1260. [Google Scholar] [CrossRef]
- Moser, M.; Nagamori, S.; Huber, M.; Tokuda, H.; Nishiyama, K. Glycolipozyme MPIase is essential for topology inversion of SecG during preprotein translocation. Proc. Natl. Acad. Sci. USA 2013, 110, 9734–9739. [Google Scholar] [CrossRef]
- Chen, M.; Samuelson, J.C.; Jiang, F.; Muller, M.; Kuhn, A.; Dalbey, R.E. Direct Interaction of YidC with the Sec-independent Pf3 Coat Protein during Its Membrane Protein Insertion. J. Biol. Chem. 2002, 277, 7670–7675. [Google Scholar] [CrossRef] [PubMed]
- Samuelson, J.C.; Chen, M.; Jiang, F.; Möller, I.; Wiedmann, M.; Kuhn, A.; Phillips, G.J.; Dalbey, R.E. YidC mediates membrane protein insertion in bacteria. Nature 2000, 406, 637–641. [Google Scholar] [CrossRef] [PubMed]
- Serek, J.; Bauer-Manz, G.; Struhalla, G.; van den Berg, L.; Kiefer, D.; Dalbey, R.; Kuhn, A. Escherichia coli YidC is a membrane insertase for Sec-independent proteins. EMBO J. 2004, 23, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Kumazaki, K.; Chiba, S.; Takemoto, M.; Furukawa, A.; Nishiyama, K.; Sugano, Y.; Mori, T.; Dohmae, N.; Hirata, K.; Nakada-Nakura, Y.; et al. Structural basis of Sec-independent membrane protein insertion by YidC. Nature 2014, 509, 516–520. [Google Scholar] [CrossRef] [PubMed]
- McDowell, M.A.; Heimes, M.; Sinning, I. Structural and molecular mechanisms for membrane protein biogenesis by the Oxa1 superfamily. Nat. Struct. Mol. Biol. 2021, 28, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Wickles, S.; Singharoy, A.; Andreani, J.; Seemayer, S.; Bischoff, L.; Berninghausen, O.; Soeding, J.; Schulten, K.; van der Sluis, E.O.; Beckmann, R. A structural model of the active ribosome-bound membrane protein insertase YidC. Elife 2014, 3, e03035. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y.; Shimizu, Y.; Nishikawa, H.; Sawasato, K.; Nishiyama, K.I. Interplay between MPIase, YidC, and PMF during Sec-independent insertion of membrane proteins. Life Sci. Alliance 2022, 5, e202101162. [Google Scholar] [CrossRef]
- Sasaki, M.; Nishikawa, H.; Suzuki, S.; Moser, M.; Huber, M.; Sawasato, K.; Matsubayashi, H.T.; Kumazaki, K.; Tsukazaki, T.; Kuruma, Y.; et al. The bacterial protein YidC accelerates MPIase-dependent integration of membrane proteins. J. Biol. Chem. 2019, 294, 18898–18908. [Google Scholar] [CrossRef]
- Fujikawa, K.; Han, Y.; Osawa, T.; Mori, S.; Nomura, K.; Muramoto, M.; Nishiyama, K.-I.; Shimamoto, K. Structural Requirements of a Glycolipid MPIase for Membrane Protein Integration. Chem. Eur. J. 2023, 29, e202300437. [Google Scholar] [CrossRef]
- Fujikawa, K.; Suzuki, S.; Nagase, R.; Ikeda, S.; Mori, S.; Nomura, K.; Nishiyama, K.I.; Shimamoto, K. Syntheses and Activities of the Functional Structures of a Glycolipid Essential for Membrane Protein Integration. ACS Chem. Biol. 2018, 13, 2719–2727. [Google Scholar] [CrossRef] [PubMed]
- Mori, S.; Nomura, K.; Fujikawa, K.; Osawa, T.; Shionyu, M.; Yoda, T.; Shirai, T.; Tsuda, S.; Yoshizawa-Kumagaye, K.; Masuda, S.; et al. Intermolecular Interactions between a Membrane Protein and a Glycolipid Essential for Membrane Protein Integration. ACS Chem. Biol. 2022, 17, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Nomura, K.; Mori, S.; Fujikawa, K.; Osawa, T.; Tsuda, S.; Yoshizawa-Kumagaye, K.; Masuda, S.; Nishio, H.; Yoshiya, T.; Yoda, T.; et al. Role of a bacterial glycolipid in Sec-independent membrane protein insertion. Sci. Rep. 2022, 12, 12231. [Google Scholar] [CrossRef] [PubMed]
- Nomura, K.; Yamaguchi, T.; Mori, S.; Fujikawa, K.; Nishiyama, K.I.; Shimanouchi, T.; Tanimoto, Y.; Morigaki, K.; Shimamoto, K. Alteration of Membrane Physicochemical Properties by Two Factors for Membrane Protein Integration. Biophys. J. 2019, 117, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Soblosky, L.; Nguyen, K.; Geng, J.; Yu, X.; Ramamoorthy, A.; Chen, Z. Physiologically-relevant modes of membrane interactions by the human antimicrobial peptide, LL-37, revealed by SFG experiments. Sci. Rep. 2013, 3, 1854. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.H.; Heng, C.; Separovic, F.; Aguilar, M.I. Comparison of reversible membrane destabilisation induced by antimicrobial peptides derived from Australian frogs. Biochim. Biophys. Acta 2014, 1838, 2205–2215. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Feigenson, G.W. A microscopic interaction model of maximum solubility of cholesterol in lipid bilayers. Biophys. J. 1999, 76, 2142–2157. [Google Scholar] [CrossRef] [PubMed]
- Strandberg, E.; Ulrich, A.S. AMPs and OMPs: Is the folding and bilayer insertion of beta-stranded outer membrane proteins governed by the same biophysical principles as for alpha-helical antimicrobial peptides? Biochim. Biophys. Acta 2015, 1848, 1944–1954. [Google Scholar] [CrossRef]
- Yano, Y.; Yamamoto, A.; Ogura, M.; Matsuzaki, K. Thermodynamics of Insertion and Self-Association of a Transmembrane Helix: A Lipophobic Interaction by Phosphatidylethanolamine. Biochemistry 2011, 50, 6806–6814. [Google Scholar] [CrossRef]
- Shintou, K.; Nakano, M.; Kamo, T.; Kuroda, Y.; Handa, T. Interaction of an Amphipathic Peptide with Phosphatidycholine/Phosphatidylethanolamine Mixed Membranes. Biophys. J. 2007, 93, 3900–3906. [Google Scholar] [CrossRef]
- Murayama, T.; Masuda, T.; Afonin, S.; Kawano, K.; Takatani-Nakase, T.; Ida, H.; Takahashi, Y.; Fukuma, T.; Ulrich, A.S.; Futaki, S. Loosening of Lipid Packing Promotes Oligoarginine Entry into Cells. Angew. Chem. Int. Ed. 2017, 56, 7644–7647. [Google Scholar] [CrossRef]
- Pujals, S.; Miyamae, H.; Afonin, S.; Murayama, T.; Hirose, H.; Nakase, I.; Taniuchi, K.; Umeda, M.; Sakamoto, K.; Ulrich, A.S.; et al. Curvature engineering: Positive membrane curvature induced by epsin N-terminal peptide boosts internalization of octaarginine. ACS Chem. Biol. 2013, 8, 1894–1899. [Google Scholar] [CrossRef]
- Bogdanov, M.; Xie, J.; Dowhan, W. Lipid-protein interactions drive membrane protein topogenesis in accordance with the positive inside rule. J. Biol. Chem. 2009, 284, 9637–9641. [Google Scholar] [CrossRef]
- Von Heijne, G. Membrane-protein topology. Nat. Rev. Mol. Cell. Biol. 2006, 7, 909–918. [Google Scholar] [CrossRef] [PubMed]
- Nevzorov, A.A.; Opella, S.J. Selective averaging for high-resolution solid-state NMR spectroscopy of aligned samples. J. Magn. Reson. 2007, 185, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Nevzorov, A.A.; Opella, S.J. A “magic sandwich” pulse sequence with reduced offset dependence for high-resolution separated local field spectroscopy. J. Magn. Reson. 2003, 164, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jardetzky, O. Probability-based protein secondary structure identification using combined NMR chemical-shift data. Protein Sci. 2002, 11, 852–861. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Sykes, B.D.; Richards, F.M. Relationship between nuclear magnetic resonance chemical shift and protein secondary structure. J. Mol. Biol. 1991, 222, 311–333. [Google Scholar] [CrossRef] [PubMed]
- Lentz, B.R. Use of fluorescent probes to monitor molecular order and motions within liposome bilayers. Chem. Phys. Lipids 1993, 64, 99–116. [Google Scholar] [CrossRef] [PubMed]
- Antollini, S.S.; Soto, M.A.; Bonini de Romanelli, I.; Gutiérrez-Merino, C.; Sotomayor, P.; Barrantes, F.J. Physical state of bulk and protein-associated lipid in nicotinic acetylcholine receptor-rich membrane studied by laurdan generalized polarization and fluorescence energy transfer. Biophys. J. 1996, 70, 1275–1284. [Google Scholar] [CrossRef]
- Carrier, D.; Giziewicz, J.B.; Moir, D.; Smith, I.C.; Jarrell, H.C. Dynamics and orientation of glycolipid headgroups by 2H-NMR: Gentiobiose. Biochim. Biophys. Acta 1989, 983, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Jarrell, H.C.; Wand, A.J.; Giziewicz, J.B.; Smith, I.C. The dependence of glyceroglycolipid orientation and dynamics on head-group structure. Biochim. Biophys. Acta 1987, 897, 69–82. [Google Scholar] [CrossRef]
- Renou, J.P.; Giziewicz, J.B.; Smith, I.C.; Jarrell, H.C. Glycolipid membrane surface structure: Orientation, conformation, and motion of a disaccharide headgroup. Biochemistry 1989, 28, 1804–1814. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Sakae, Y.; Zhang, Y.; Yamamoto, S.; Okamoto, Y.; Kato, K. Exploration of conformational spaces of high-mannose-type oligosaccharides by an NMR-validated simulation. Angew. Chem. Int. Ed. 2014, 53, 10941–10944. [Google Scholar] [CrossRef]
- Bai, J.; Pagano, R.E. Measurement of spontaneous transfer and transbilayer movement of BODIPY-labeled lipids in lipid vesicles. Biochemistry 1997, 36, 8840–8848. [Google Scholar] [CrossRef] [PubMed]
- Strandberg, E.; Tiltak, D.; Ehni, S.; Wadhwani, P.; Ulrich, A.S. Lipid shape is a key factor for membrane interactions of amphipathic helical peptides. Biochim. Biophys. Acta 2012, 1818, 1764–1776. [Google Scholar] [CrossRef]
- Chemical Computing Group ULC. MOE (Molecular Operating Environment); Version 2022.02; Chemical Computing Group ULC: Montreal, QC, Canada, 2022. [Google Scholar]
- Nishikawa, H.; Sasaki, M.; Nishiyama, K.I. In vitro Assay for Bacterial Membrane Protein Integration into Proteoliposomes. Bio-Protoc. 2020, 10, e3626. [Google Scholar] [CrossRef] [PubMed]
- Jason-Moller, L.; Murphy, M.; Bruno, J. Overview of Biacore systems and their applications. Curr. Protoc. Protein Sci. 2006, 45, 19.13.1–19.13.14. [Google Scholar] [CrossRef]
- O’Shannessy, D.J.; Brigham-Burke, M.; Soneson, K.K.; Hensley, P.; Brooks, I. Determination of rate and equilibrium binding constants for macromolecular interactions by surface plasmon resonance. Meth. Enzymol. 1994, 240, 323–349. [Google Scholar]
- Yamazaki, T.; Pascal, S.M.; Singer, A.U.; Forman-Kay, J.D.; Kay, L.E. NMR Pulse Schemes for the Sequence-Specific Assignment of Arginine Guanidino 15N and 1H Chemical Shifts in Proteins. J. Am. Chem. Soc. 1995, 117, 3556–3564. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nomura, K.; Mori, S.; Shimamoto, K. Roles of a Glycolipid MPIase in Sec-Independent Membrane Protein Insertion. Membranes 2024, 14, 48. https://doi.org/10.3390/membranes14020048
Nomura K, Mori S, Shimamoto K. Roles of a Glycolipid MPIase in Sec-Independent Membrane Protein Insertion. Membranes. 2024; 14(2):48. https://doi.org/10.3390/membranes14020048
Chicago/Turabian StyleNomura, Kaoru, Shoko Mori, and Keiko Shimamoto. 2024. "Roles of a Glycolipid MPIase in Sec-Independent Membrane Protein Insertion" Membranes 14, no. 2: 48. https://doi.org/10.3390/membranes14020048