
Bioinspired Nanoplatforms Based on Graphene Oxide and Neurotrophin-Mimicking Peptides

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Preparation of Self-Assembled Lipids (SUVs, SLBs)
2.3. Peptides Synthesis and Purification
2.4. Peptides Immobilization on GO Sheets
2.5. Physicochemical Characterization
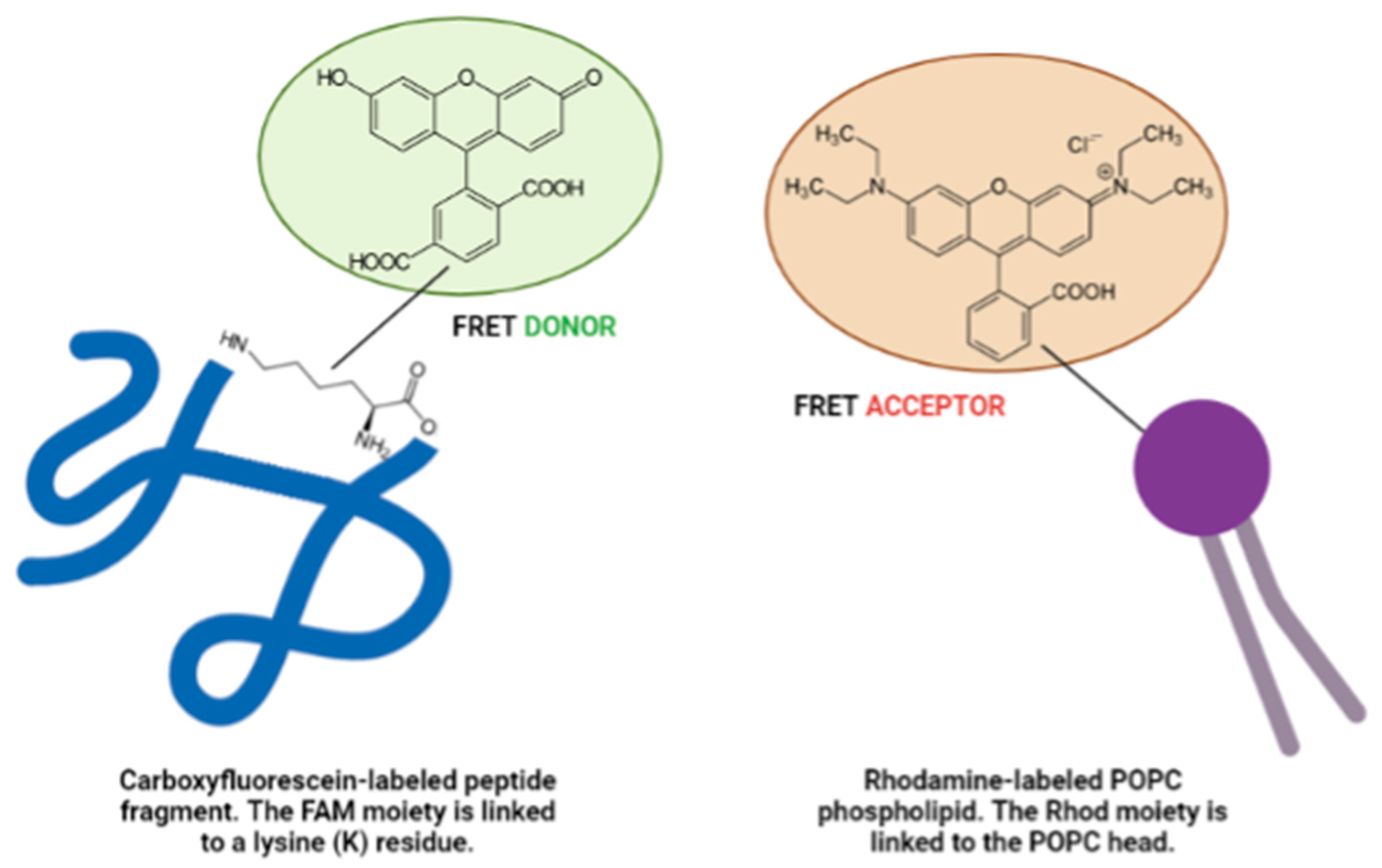
2.5.1. UV-Visible (UV-Vis) Spectroscopy and Fluorescence Spectroscopy with Fluorescence Resonance Energy Transfer (FRET) Analysis
2.5.2. Scanning Confocal Microscope (LSM) Imaging and Fluorescence Recovery after Photobleaching (FRAP) Analysis
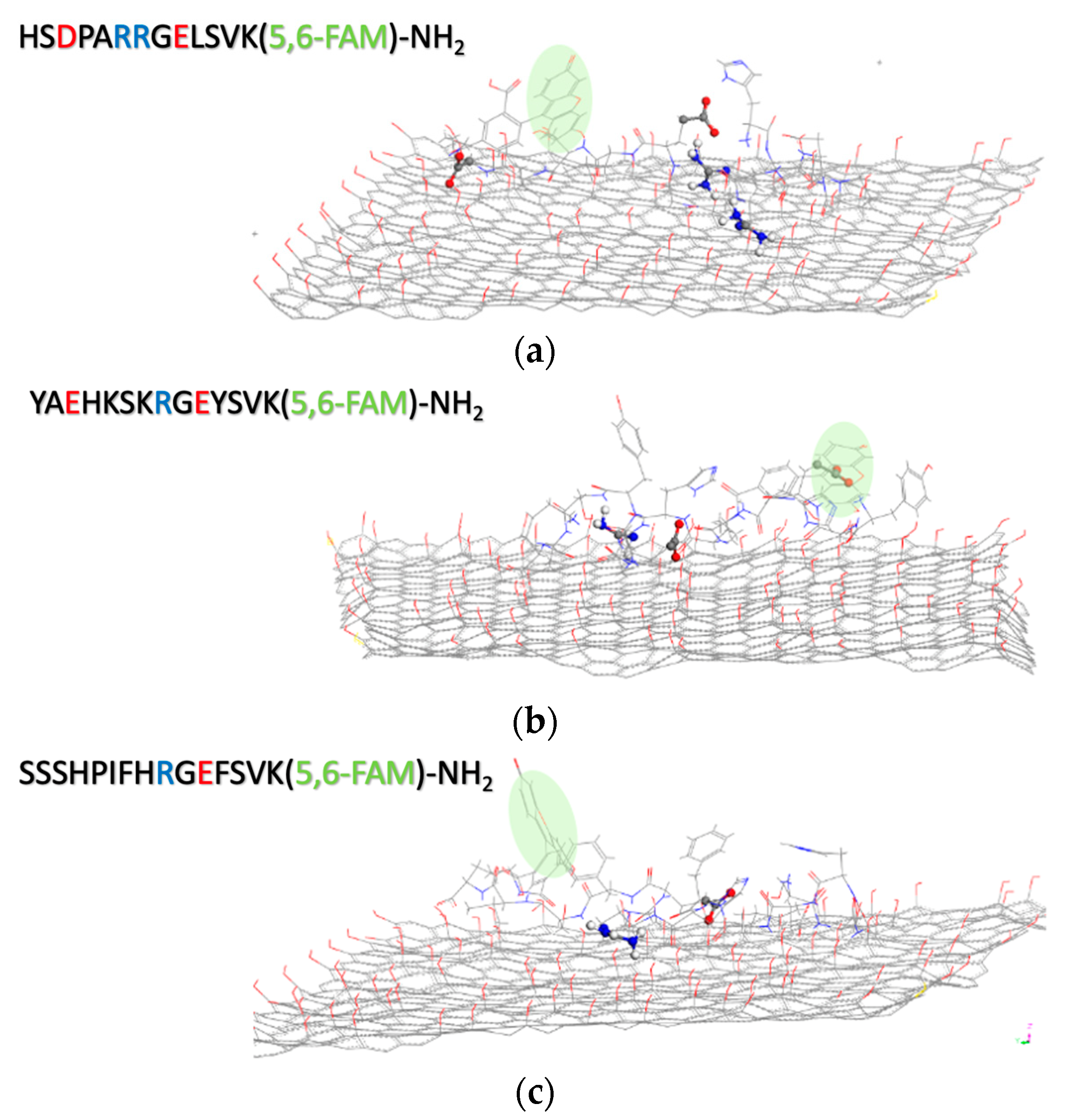
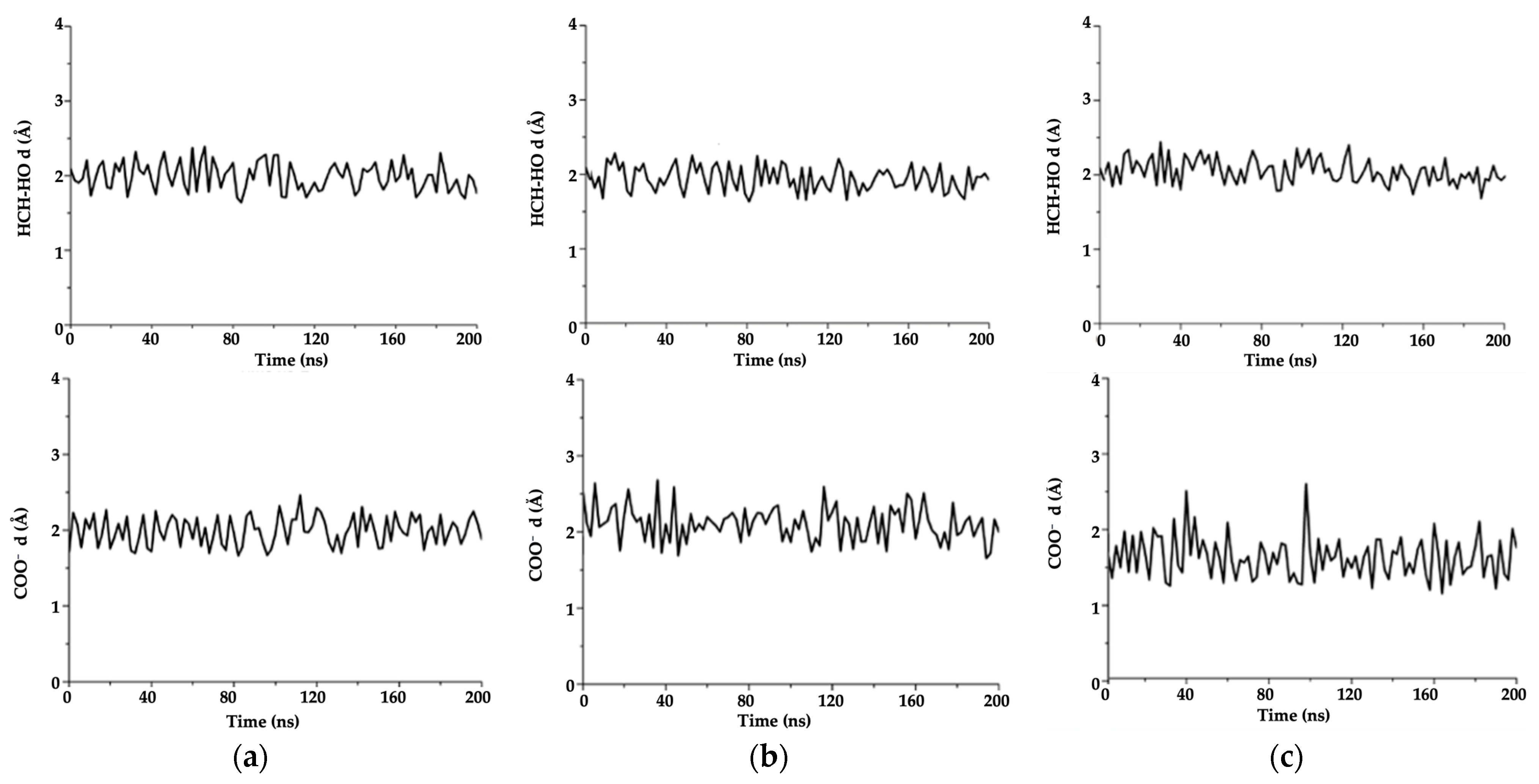
2.5.3. Molecular Dynamics (MD)
2.6. Biochemical/Cellular Characterization
2.6.1. Cell Cultures and Maintenance
2.6.2. Cytotoxicity
2.6.3. Neurite Outgrowth
2.6.4. Wound Closure Assay
2.6.5. Tube Formation Assay
3. Results and Discussion
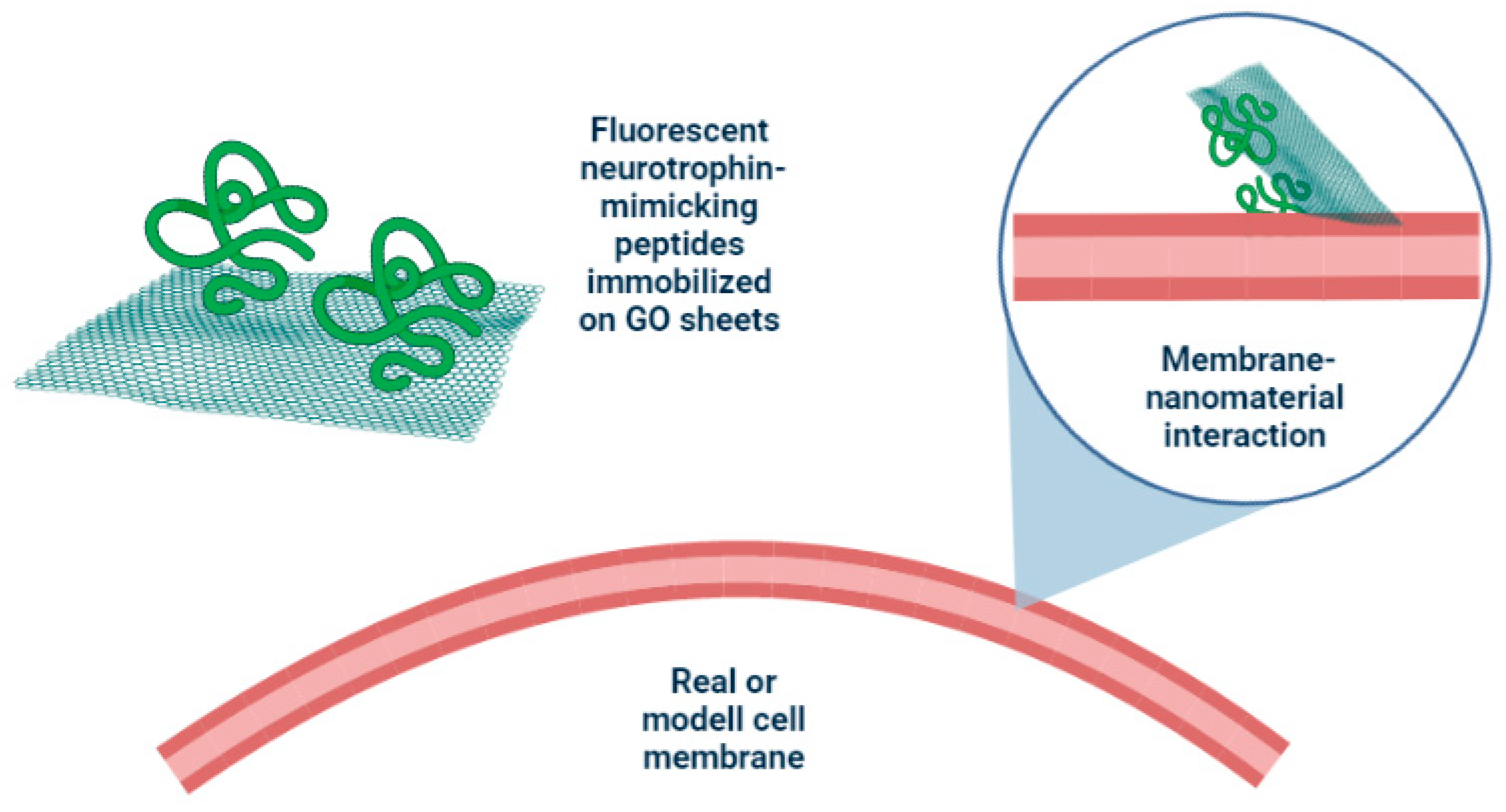
3.1. The Interaction of Peptide-Functionalized GO with Model Cell Membranes
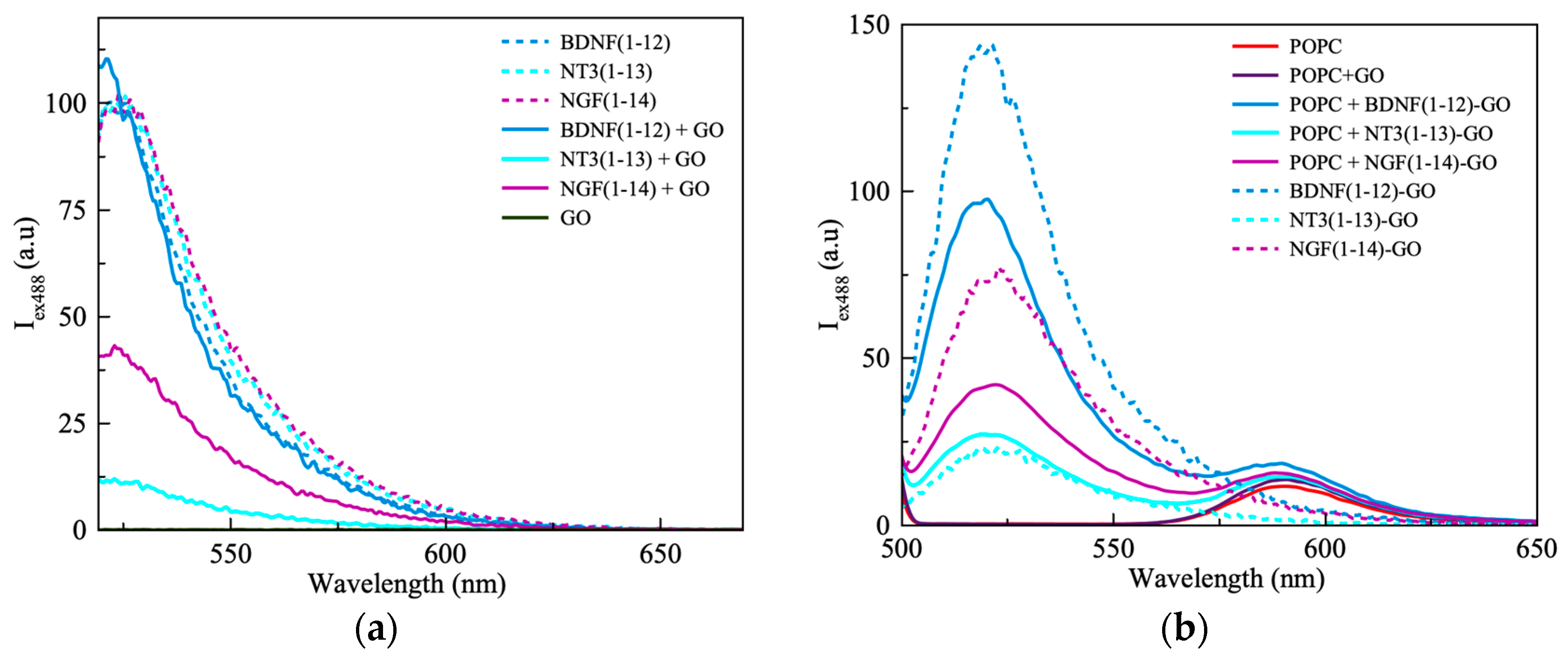
3.1.1. Three-Dimensional FRET Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pep−GO | Interaction Forces |
|---|---|
| BDNF(1-12) | Electrostatics, H-bonds between GO and NH (Arg, 2) and COOH (Asp, 1) |
| NT3(1-13) | More effective contact surface for H-bonds between GO and NH (Arg, 1) and COOH (Glu, 2) |
| NGF(1-14) | The highest number of binding sites between GO and NH (Arg, 1) and COOH (Glu, 1), lower H-bond strength. |
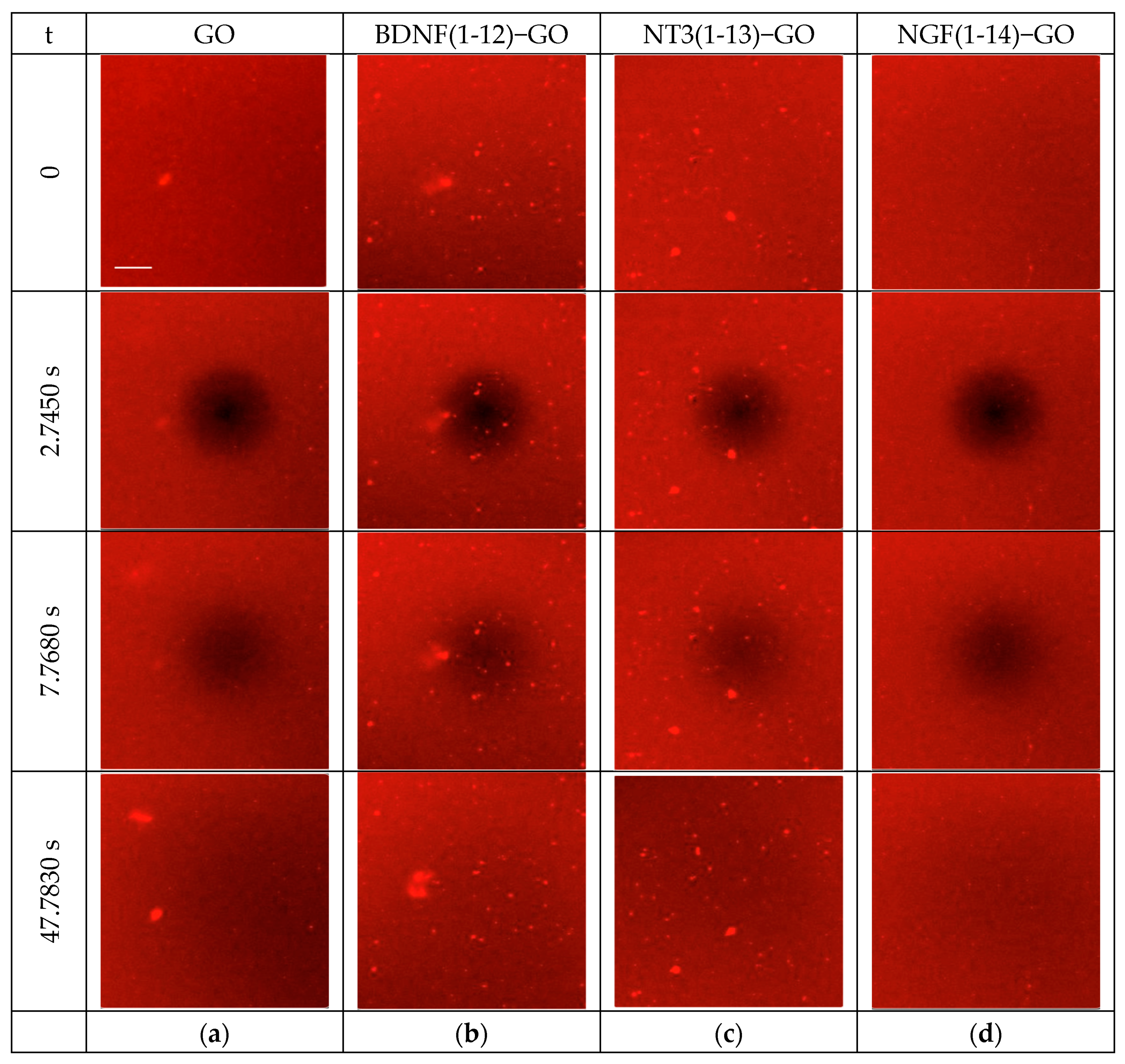
3.1.2. Two-Dimensional FRAP Analyses
3.2. The Interaction of Peptide-Functionalized GO with Cells
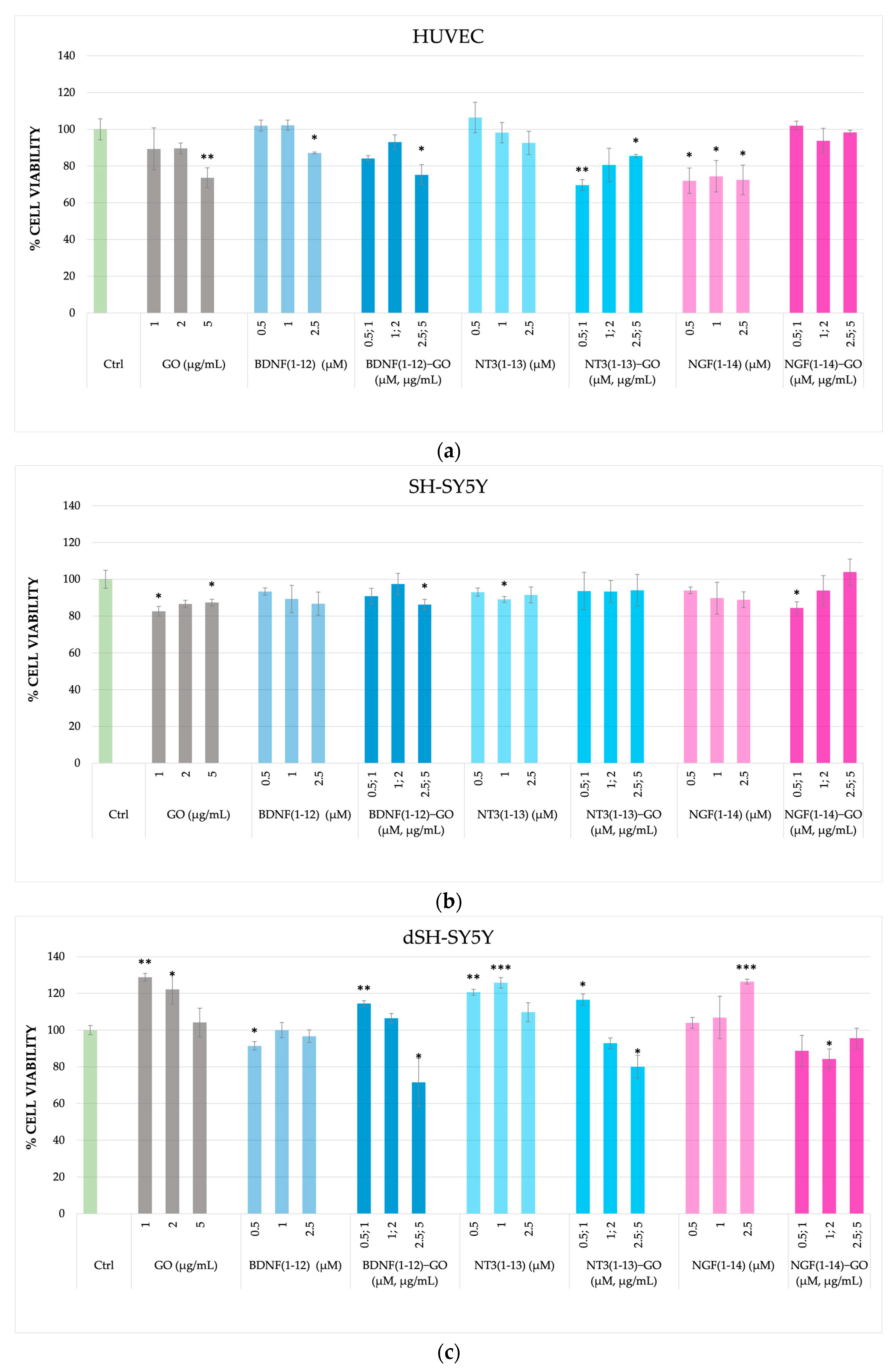
3.2.1. Cytotoxicity of NT Peptide-Functionalized GO in SH-SY5Y and HUVECs
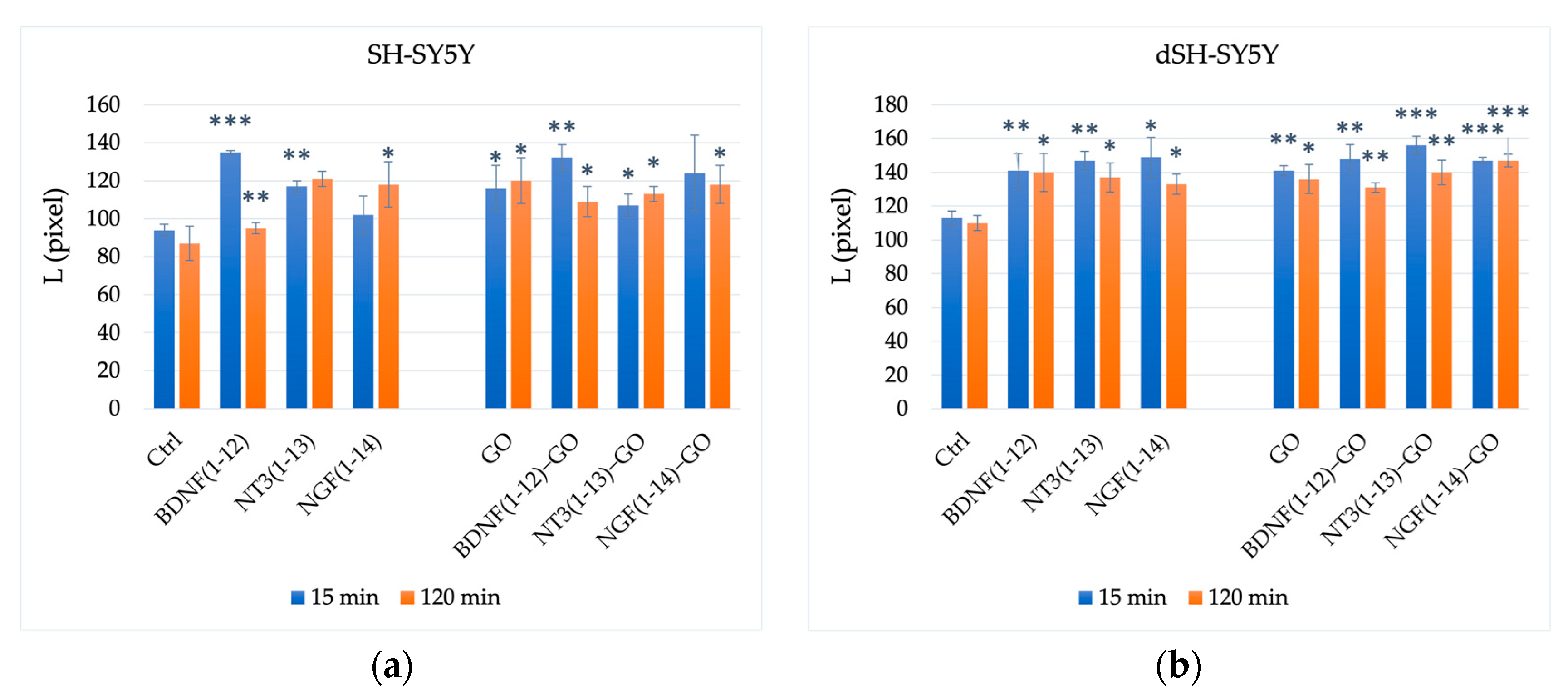
3.2.2. Neuroblastoma and Neuron-like Cells (Neurite Outgrowth)
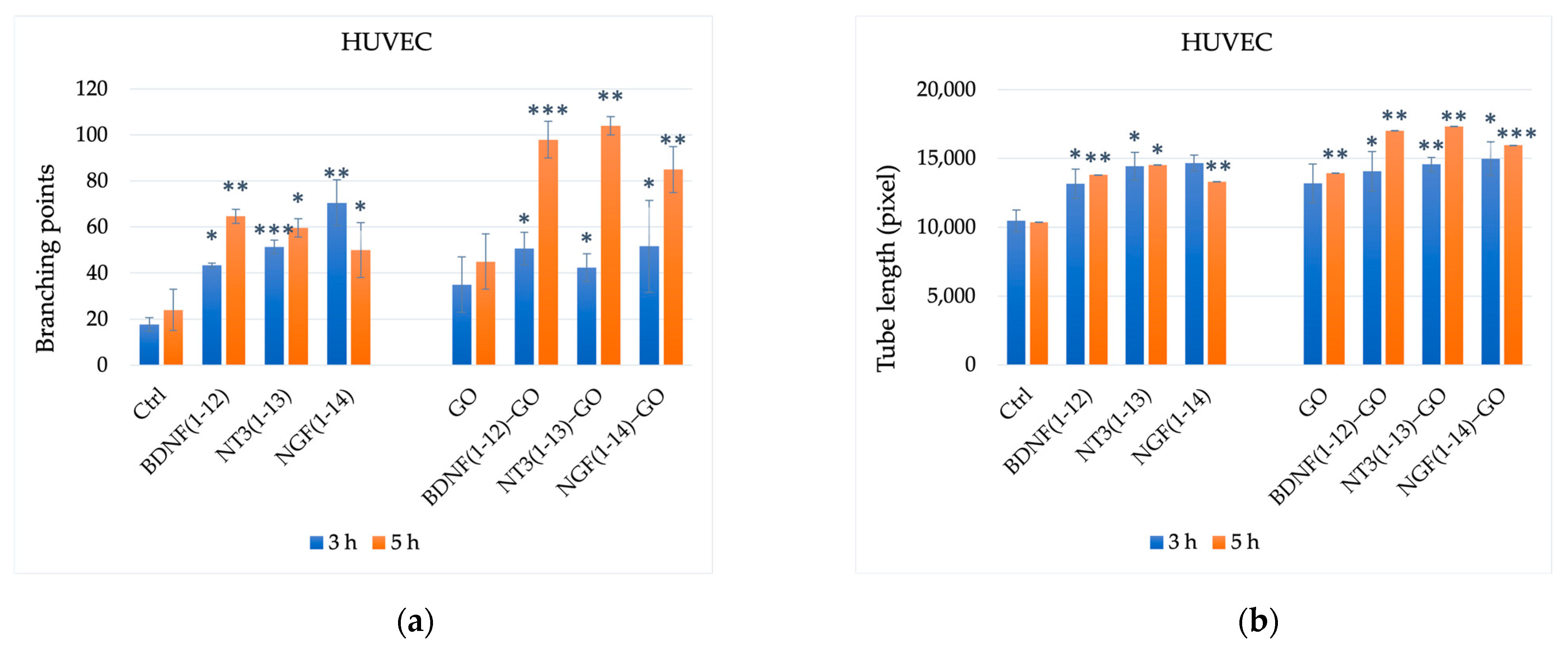
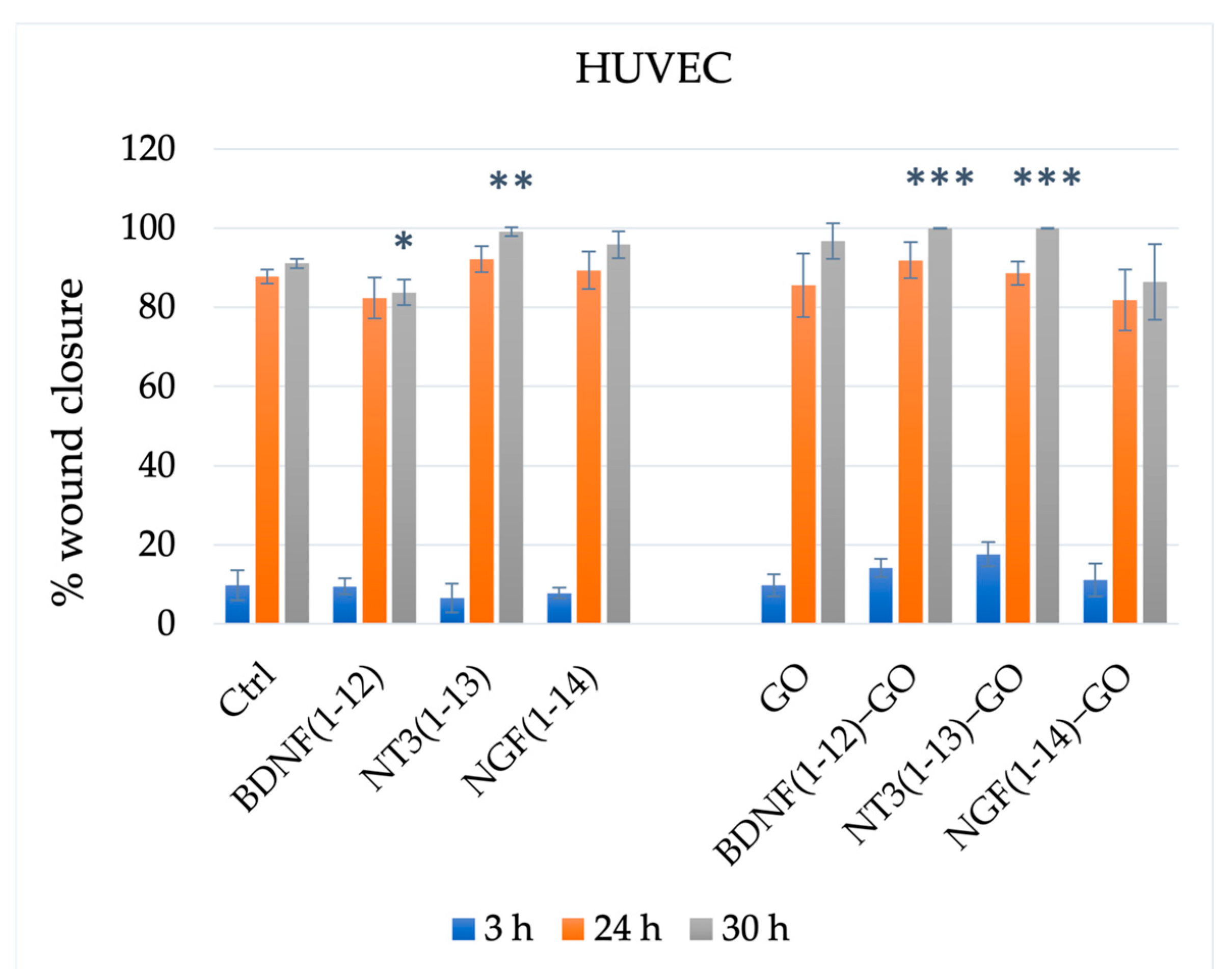
3.2.3. Endothelial Cells (Angiogenesis Assays)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Tupone, M.G.; Panella, G.; d’Angelo, M.; Castelli, V.; Caioni, G.; Catanesi, M.; Benedetti, E.; Cimini, A. An Update on Graphene-Based Nanomaterials for Neural Growth and Central Nervous System Regeneration. Int. J. Mol. Sci. 2021, 22, 13047. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Yang, Z.; Li, S.; Le, W. Nanotherapeutic and Stem Cell Therapeutic Strategies in Neurodegenerative Diseases: A Promising Therapeutic Approach. Int. J. Nanomed. 2023, 18, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Song, Q.; Li, N.; Jiang, Z.; Huang, R.; Cheng, G. Enhancement of electrical signaling in neural networks on graphene films. Biomaterials 2013, 34, 6402–6411. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, H.; Zhang, Y.; Huangfu, H.; Yang, Y.; Qin, Q.; Zhang, Y.; Zhou, Y. 3D printed reduced graphene oxide-GelMA hybrid hydrogel scaffolds for potential neuralized bone regeneration. J. Mater. Chem. B 2023, 11, 1288–1301. [Google Scholar] [CrossRef] [PubMed]
- Rauti, R.; Lozano, N.; León, V.; Scaini, D.; Musto, M.; Rago, I.; Ulloa Severino, F.P.; Fabbro, A.; Casalis, L.; Vázquez, E.; et al. Graphene Oxide Nanosheets Reshape Synaptic Function in Cultured Brain Networks. ACS Nano 2016, 10, 4459–4471. [Google Scholar] [CrossRef] [PubMed]
- Secomandi, N.; Franceschi Biagioni, A.; Kostarelos, K.; Cellot, G.; Ballerini, L. Thin graphene oxide nanoflakes modulate glutamatergic synapses in the amygdala cultured circuits: Exploiting synaptic approaches to anxiety disorders. Nanomed. Nanotechnol. Biol. Med. 2020, 26, 102174. [Google Scholar] [CrossRef] [PubMed]
- Bramini, M.; Alberini, G.; Colombo, E.; Chiacchiaretta, M.; DiFrancesco, M.L.; Maya-Vetencourt, J.F.; Maragliano, L.; Benfenati, F.; Cesca, F. Interfacing Graphene-Based Materials with Neural Cells. Front. Syst. Neurosci. 2018, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Consiglio, G.; Di Pietro, P.; D’Urso, L.; Forte, G.; Grasso, G.; Sgarlata, C.; Cossement, D.; Snyders, R.; Satriano, C. Surface tailoring of polyacrylate-grafted graphene oxide for controlled interactions at the biointerface. J. Colloid Interface Sci. 2017, 506, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in Neuronal Development and Function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef]
- Lad, S.; Neet, K.; Mufson, E. Nerve Growth Factor: Structure, Function and Therapeutic Implications for Alzheimers Disease. Curr. Drug Target-CNS Neurol. Disord. 2003, 2, 315–334. [Google Scholar] [CrossRef]
- Yano, H.; Chao, M.V. Neurotrophin receptor structure and interactions. In Receptor Chemistry towards the Third Millennium, Proceedings of the 12th Camerino-Noordwijkerhout Symposium, Camerino, Italy, 5–9 September 1999; Pharmacochemistry Library: New York, NY, USA, 2000; pp. 253–260. [Google Scholar]
- Park, H.; Poo, M.-M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2012, 14, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Christian, K.; Lu, B. BDNF: A key regulator for protein synthesis-dependent LTP and long-term memory? Neurobiol. Learn. Mem. 2008, 89, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-H.; Liao, X.-M.; Liu, D.; Hu, J.; Yin, Y.-Y.; Wang, J.-Z.; Zhu, L.-Q. NGF promotes long-term memory formation by activating poly(ADP-ribose)polymerase-1. Neuropharmacology 2012, 63, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Amidfar, M.; de Oliveira, J.; Kucharska, E.; Budni, J.; Kim, Y.-K. The role of CREB and BDNF in neurobiology and treatment of Alzheimer’s disease. Life Sci. 2020, 257, 118020. [Google Scholar] [CrossRef]
- Castelli, V.; Alfonsetti, M.; d’Angelo, M. Neurotrophic factor-based pharmacological approaches in neurological disorders. Neural Regen. Res. 2023, 18, 1220–1228. [Google Scholar] [CrossRef]
- Numakawa, T.; Odaka, H. The Role of Neurotrophin Signaling in Age-Related Cognitive Decline and Cognitive Diseases. Int. J. Mol. Sci. 2022, 23, 7726. [Google Scholar] [CrossRef]
- Nordvall, G.; Forsell, P.; Sandin, J. Neurotrophin-targeted therapeutics: A gateway to cognition and more? Drug Discov. Today 2022, 27, 103318. [Google Scholar] [CrossRef]
- Bondarenko, O.; Saarma, M. Neurotrophic Factors in Parkinson’s Disease: Clinical Trials, Open Challenges and Nanoparticle-Mediated Delivery to the Brain. Front. Cell. Neurosci. 2021, 15, 682597. [Google Scholar] [CrossRef]
- Padmakumar, S.; Taha, M.S.; Kadakia, E.; Bleier, B.S.; Amiji, M.M. Delivery of neurotrophic factors in the treatment of age-related chronic neurodegenerative diseases. Expert Opin. Drug Deliv. 2020, 17, 323–340. [Google Scholar] [CrossRef]
- Gordon, T. The role of neurotrophic factors in nerve regeneration. Neurosurg. Focus 2009, 26, E3. [Google Scholar] [CrossRef]
- Houlton, J.; Abumaria, N.; Hinkley, S.F.R.; Clarkson, A.N. Therapeutic Potential of Neurotrophins for Repair After Brain Injury: A Helping Hand From Biomaterials. Front. Neurosci. 2019, 13, 790. [Google Scholar] [CrossRef] [PubMed]
- El Ouaamari, Y.; Van den Bos, J.; Willekens, B.; Cools, N.; Wens, I. Neurotrophic Factors as Regenerative Therapy for Neurodegenerative Diseases: Current Status, Challenges and Future Perspectives. Int. J. Mol. Sci. 2023, 24, 3866. [Google Scholar] [CrossRef]
- Lindholm, D.; Heumann, R.; Meyer, M.; Thoenen, H. Interleukin-1 regulates synthesis of nerve growth factor in non-neuronal cells of rat sciatic nerve. Nature 1987, 330, 658–659. [Google Scholar] [CrossRef]
- Ricci, A.; Felici, L.; Mariotta, S.; Mannino, F.; Schmid, G.; Terzano, C.; Cardillo, G.; Amenta, F.; Bronzetti, E. Neurotrophin and Neurotrophin Receptor Protein Expression in the Human Lung. Am. J. Respir. Cell Mol. Biol. 2004, 30, 12–19. [Google Scholar] [CrossRef]
- Pius-Sadowska, E.; Machaliński, B. BDNF—A key player in cardiovascular system. J. Mol. Cell. Cardiol. 2017, 110, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Blais, M.; Lévesque, P.; Bellenfant, S.; Berthod, F. Nerve Growth Factor, Brain-Derived Neurotrophic Factor, Neurotrophin-3 and Glial-Derived Neurotrophic Factor Enhance Angiogenesis in a Tissue-EngineeredIn VitroModel. Tissue Eng. Part A 2013, 19, 1655–1664. [Google Scholar] [CrossRef]
- Nico, B.; Mangieri, D.; Benagiano, V.; Crivellato, E.; Ribatti, D. Nerve growth factor as an angiogenic factor. Microvasc. Res. 2008, 75, 135–141. [Google Scholar] [CrossRef]
- Carmeliet, P. Angiogenesis in health and disease. Nat. Med. 2003, 9, 653–660. [Google Scholar] [CrossRef]
- Cantarella, G.; Lempereur, L.; Presta, M.; Ribatti, D.; Lombardo, G.; Lazarovici, P.; Zappalà, G.; Pafumi, C.; Bernardini, R. Nerve growth factor–endothelial cell interaction leads to angiogenesis in vitro and in vivo. FASEB J. 2002, 16, 1307–1309. [Google Scholar] [CrossRef]
- Raychaudhuri, S.K.; Raychaudhuri, S.P.; Weltman, H.; Farber, E.M. Effect of nerve growth factor on endothelial cell biology: Proliferation and adherence molecule expression on human dermal microvascular endothelial cells. Arch. Dermatol. Res. 2001, 293, 291–295. [Google Scholar] [CrossRef]
- Moser, K.V.; Reindl, M.; Blasig, I.; Humpel, C. Brain capillary endothelial cells proliferate in response to NGF, express NGF receptors and secrete NGF after inflammation. Brain Res. 2004, 1017, 53–60. [Google Scholar] [CrossRef]
- Lambiase, A.; Manni, L.; Bonini, S.; Rama, P.; Micera, A.; Aloe, L. Nerve growth factor promotes corneal healing: Structural, biochemical, and molecular analyses of rat and human corneas. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1063–1069. [Google Scholar]
- Diao, Y.-P.; Cui, F.-K.; Yan, S.; Chen, Z.-G.; Lian, L.-S.; Guo, L.-L.; Li, Y.-J. Nerve Growth Factor Promotes Angiogenesis and Skeletal Muscle Fiber Remodeling in a Murine Model of Hindlimb Ischemia. Chin. Med. J. 2016, 129, 313–319. [Google Scholar] [CrossRef]
- Cristofaro, B.; Stone, O.A.; Caporali, A.; Dawbarn, D.; Ieronimakis, N.; Reyes, M.; Madeddu, P.; Bates, D.O.; Emanueli, C. Neurotrophin-3 Is a Novel Angiogenic Factor Capable of Therapeutic Neovascularization in a Mouse Model of Limb Ischemia. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1143–1150. [Google Scholar] [CrossRef]
- El Baassiri, M.; Dosh, L.; Haidar, H.; Gerges, A.; Baassiri, S.; Leone, A.; Rappa, F.; Jurjus, A. Nerve growth factor and burn wound healing: Update of molecular interactions with skin cells. Burns 2022. [Google Scholar] [CrossRef]
- Kermani, P.; Hempstead, B. Brain-Derived Neurotrophic Factor: A Newly Described Mediator of Angiogenesis. Trends Cardiovasc. Med. 2007, 17, 140–143. [Google Scholar] [CrossRef]
- Usui, T.; Naruo, A.; Okada, M.; Hayabe, Y.; Yamawaki, H. Brain-derived neurotrophic factor promotes angiogenic tube formation through generation of oxidative stress in human vascular endothelial cells. Acta Physiol. 2014, 211, 385–394. [Google Scholar] [CrossRef]
- Julio-Pieper, M.; Lozada, P.; Tapia, V.; Vega, M.; Miranda, C.; Vantman, D.; Ojeda, S.R.; Romero, C. Nerve Growth Factor Induces Vascular Endothelial Growth Factor Expression in Granulosa Cells via a trkA Receptor/Mitogen-Activated Protein Kinase-Extracellularly Regulated Kinase 2-Dependent Pathway. J. Clin. Endocrinol. Metab. 2009, 94, 3065–3071. [Google Scholar] [CrossRef]
- Mori, A.; Nishioka, Y.; Yamada, M.; Nishibata, Y.; Masuda, S.; Tomaru, U.; Honma, N.; Moriyama, T.; Ishizu, A. Brain-derived neurotrophic factor induces angiogenin secretion and nuclear translocation in human umbilical vein endothelial cells. Pathol. Res. Pract. 2018, 214, 521–526. [Google Scholar] [CrossRef]
- Magrì, A.; Munzone, A.; Peana, M.; Medici, S.; Zoroddu, M.; Hansson, O.; Satriano, C.; Rizzarelli, E.; La Mendola, D. Coordination Environment of Cu(II) Ions Bound to N-Terminal Peptide Fragments of Angiogenin Protein. Int. J. Mol. Sci. 2016, 17, 1240. [Google Scholar] [CrossRef] [PubMed]
- Cucci, L.M.; Satriano, C.; Marzo, T.; La Mendola, D. Angiogenin and Copper Crossing in Wound Healing. Int. J. Mol. Sci. 2021, 22, 10704. [Google Scholar] [CrossRef] [PubMed]
- Grasman, J.M.; Kaplan, D.L. Human endothelial cells secrete neurotropic factors to direct axonal growth of peripheral nerves. Sci. Rep. 2017, 7, 4092. [Google Scholar] [CrossRef] [PubMed]
- Alastra, G.; Aloe, L.; Baldassarro, V.A.; Calzà, L.; Cescatti, M.; Duskey, J.T.; Focarete, M.L.; Giacomini, D.; Giardino, L.; Giraldi, V.; et al. Nerve Growth Factor Biodelivery: A Limiting Step in Moving Toward Extensive Clinical Application? Front. Neurosci. 2021, 15, 695592. [Google Scholar] [CrossRef] [PubMed]
- Kemp, S.W.P.; Webb, A.A.; Dhaliwal, S.; Syed, S.; Walsh, S.K.; Midha, R. Dose and duration of nerve growth factor (NGF) administration determine the extent of behavioral recovery following peripheral nerve injury in the rat. Exp. Neurol. 2011, 229, 460–470. [Google Scholar] [CrossRef]
- Trapani, G.; Satriano, C.; La Mendola, D. Peptides and their Metal Complexes in Neurodegenerative Diseases: From Structural Studies to Nanomedicine Prospects. Curr. Med. Chem. 2018, 25, 715–747. [Google Scholar] [CrossRef]
- Gascon, S.; Jann, J.; Langlois-Blais, C.; Plourde, M.; Lavoie, C.; Faucheux, N. Peptides Derived from Growth Factors to Treat Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 6071. [Google Scholar] [CrossRef]
- La Mendola, D.; Arena, G.; Pietropaolo, A.; Satriano, C.; Rizzarelli, E. Metal ion coordination in peptide fragments of neurotrophins: A crucial step for understanding the role and signaling of these proteins in the brain. Coord. Chem. Rev. 2021, 435, 213790. [Google Scholar] [CrossRef]
- Berrera, M.; Cattaneo, A.; Carloni, P. Molecular Simulation of the Binding of Nerve Growth Factor Peptide Mimics to the Receptor Tyrosine Kinase A. Biophys. J. 2006, 91, 2063–2071. [Google Scholar] [CrossRef]
- Pattarawarapan, M.; Burgess, K. Molecular Basis of Neurotrophin−Receptor Interactions. J. Med. Chem. 2003, 46, 5277–5291. [Google Scholar] [CrossRef]
- Vittorio, S.; Manelfi, C.; Gervasoni, S.; Beccari, A.R.; Pedretti, A.; Vistoli, G.; Talarico, C. Computational Insights into the Sequence-Activity Relationships of the NGF(1–14) Peptide by Molecular Dynamics Simulations. Cells 2022, 11, 2808. [Google Scholar] [CrossRef] [PubMed]
- Travaglia, A.; Pietropaolo, A.; Di Martino, R.; Nicoletti, V.G.; La Mendola, D.; Calissano, P.; Rizzarelli, E. A Small Linear Peptide Encompassing the NGF N-Terminus Partly Mimics the Biological Activities of the Entire Neurotrophin in PC12 Cells. ACS Chem. Neurosci. 2015, 6, 1379–1392. [Google Scholar] [CrossRef] [PubMed]
- Pandini, G.; Satriano, C.; Pietropaolo, A.; Gianì, F.; Travaglia, A.; La Mendola, D.; Nicoletti, V.G.; Rizzarelli, E. The Inorganic Side of NGF: Copper(II) and Zinc(II) Affect the NGF Mimicking Signaling of the N-Terminus Peptides Encompassing the Recognition Domain of TrkA Receptor. Front. Neurosci. 2016, 10, 569. [Google Scholar] [CrossRef]
- Forte, G.; Travaglia, A.; Magrì, A.; Satriano, C.; La Mendola, D. Adsorption of NGF and BDNF derived peptides on gold surfaces. Phys. Chem. Chem. Phys. 2014, 16, 1536–1544. [Google Scholar] [CrossRef]
- Satriano, C.; Forte, G.; Magrì, A.; Di Pietro, P.; Travaglia, A.; Pandini, G.; Gianì, F.; La Mendola, D. Neurotrophin-mimicking peptides at the biointerface with gold respond to copper ion stimuli. Phys. Chem. Chem. Phys. 2016, 18, 30595–30604. [Google Scholar] [CrossRef]
- Naletova, I.; Grasso, G.I.; Satriano, C.; Travaglia, A.; La Mendola, D.; Arena, G.; Rizzarelli, E. Copper complexes of synthetic peptides mimicking neurotrophin-3 enhance neurite outgrowth and CREB phosphorylation. Metallomics 2019, 11, 1567–1578. [Google Scholar] [CrossRef]
- Naletova, I.; Satriano, C.; Pietropaolo, A.; Gianì, F.; Pandini, G.; Triaca, V.; Amadoro, G.; Latina, V.; Calissano, P.; Travaglia, A.; et al. The Copper(II)-Assisted Connection between NGF and BDNF by Means of Nerve Growth Factor-Mimicking Short Peptides. Cells 2019, 8, 301. [Google Scholar] [CrossRef]
- Di Pietro, P.; Caporarello, N.; Anfuso, C.D.; Lupo, G.; Magrì, A.; La Mendola, D.; Satriano, C. Immobilization of Neurotrophin Peptides on Gold Nanoparticles by Direct and Lipid-Mediated Interaction: A New Multipotential Therapeutic Nanoplatform for CNS Disorders. ACS Omega 2017, 2, 4071–4079. [Google Scholar] [CrossRef]
- Hu, H.; Chen, X.; Zhao, K.; Zheng, W.; Gao, C. Recent Advances in Biomaterials-Based Therapies for Alleviation and Regeneration of Traumatic Brain Injury. Macromol. Biosci. 2023, 2200577. [Google Scholar] [CrossRef]
- Barui, A.K.; Roy, A.; Das, S.; Bhamidipati, K.; Patra, C.R. Therapeutic Applications of Graphene Oxides in Angiogenesis and Cancers. In Nanoparticles and Their Biomedical Applications; Shukla, A.K., Ed.; Springer Nature: New York, NY, USA, 2020; pp. 147–189. [Google Scholar]
- Sontakke, A.D.; Tiwari, S.; Purkait, M.K. A comprehensive review on graphene oxide-based nanocarriers: Synthesis, functionalization and biomedical applications. FlatChem 2023, 38, 100484. [Google Scholar] [CrossRef]
- Taheriazam, A.; Abad, G.G.Y.; Hajimazdarany, S.; Imani, M.H.; Ziaolhagh, S.; Zandieh, M.A.; Bayanzadeh, S.D.; Mirzaei, S.; Hamblin, M.R.; Entezari, M.; et al. Graphene oxide nanoarchitectures in cancer biology: Nano-modulators of autophagy and apoptosis. J. Control. Release 2023, 354, 503–522. [Google Scholar] [CrossRef] [PubMed]
- Satriano, C.; Svedhem, S.; Kasemo, B. Well-defined lipid interfaces for protein adsorption studies. Phys. Chem. Chem. Phys. 2012, 14, 16695–16698. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Huang, C.; Li, N.; Ma, X.; Li, Z.; Fan, J. Distinct roles of graphene and graphene oxide nanosheets in regulating phospholipid flip-flop. J. Colloid Interface Sci. 2023, 637, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Travaglia, A.; Satriano, C.; Giuffrida, M.L.; La Mendola, D.; Rampazzo, E.; Prodi, L.; Rizzarelli, E. Electrostatically driven interaction of silica-supported lipid bilayer nanoplatforms and a nerve growth factor-mimicking peptide. Soft Matter 2013, 9, 4648–4654. [Google Scholar] [CrossRef]
- Satriano, C.; Fragalà, M.E.; Forte, G.; Santoro, A.M.; La Mendola, D.; Kasemo, B. Surface adsorption of fibronectin-derived peptide fragments: The influence of electrostatics and hydrophobicity for endothelial cells adhesion. Soft Matter 2012, 8, 53–56. [Google Scholar] [CrossRef]
- Tomasella, P.; Sanfilippo, V.; Bonaccorso, C.; Cucci, L.M.; Consiglio, G.; Nicosia, A.; Mineo, P.G.; Forte, G.; Satriano, C. Theranostic Nanoplatforms of Thiolated Reduced Graphene Oxide Nanosheets and Gold Nanoparticles. Appl. Sci. 2020, 10, 5529. [Google Scholar] [CrossRef]
- Axelrod, D.; Koppel, D.E.; Schlessinger, J.; Elson, E.; Webb, W.W. Mobility measurement by analysis of fluorescence photobleaching recovery kinetics. Biophys. J. 1976, 16, 1055–1069. [Google Scholar] [CrossRef]
- BIOVIA. Dassault Systemes, Material Studio, San Diego; BIOVIA: San Diego, CA, USA, 2017. [Google Scholar]
- Wu, X.; Xing, Y.; Zeng, K.; Huber, K.; Zhao, J.X. Study of Fluorescence Quenching Ability of Graphene Oxide with a Layer of Rigid and Tunable Silica Spacer. Langmuir 2018, 34, 603–611. [Google Scholar] [CrossRef]
- Qi, X.; Li, H.; Lam, J.W.Y.; Yuan, X.; Wei, J.; Tang, B.Z.; Zhang, H. Graphene Oxide as a Novel Nanoplatform for Enhancement of Aggregation-Induced Emission of Silole Fluorophores. Adv. Mater. 2012, 24, 4191–4195. [Google Scholar] [CrossRef]
- Roy, A.; Kundu, N.; Banik, D.; Sarkar, N. Comparative Fluorescence Resonance Energy-Transfer Study in Pluronic Triblock Copolymer Micelle and Niosome Composed of Biological Component Cholesterol: An Investigation of Effect of Cholesterol and Sucrose on the FRET Parameters. J. Phys. Chem. B 2015, 120, 131–142. [Google Scholar] [CrossRef] [PubMed]
- dos Remedios, C.G.; Moens, P.D.J. Fluorescence Resonance Energy Transfer Spectroscopy Is a Reliable “Ruler” for Measuring Structural Changes in Proteins. J. Struct. Biol. 1995, 115, 175–185. [Google Scholar] [CrossRef]
- Pandit, S.; De, M. Interaction of Amino Acids and Graphene Oxide: Trends in Thermodynamic Properties. J. Phys. Chem. C 2017, 121, 600–608. [Google Scholar] [CrossRef]
- Neklyudov, V.V.; Khafizov, N.R.; Sedov, I.A.; Dimiev, A.M. New insights into the solubility of graphene oxide in water and alcohols. Phys. Chem. Chem. Phys. 2017, 19, 17000–17008. [Google Scholar] [CrossRef] [PubMed]
- Minton, A.P. Adsorption of Globular Proteins on Locally Planar Surfaces. II. Models for the Effect of Multiple Adsorbate Conformations on Adsorption Equilibria and Kinetics. Biophys. J. 1999, 76, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Cibecchini, G.; Veronesi, M.; Catelani, T.; Bandiera, T.; Guarnieri, D.; Pompa, P.P. Antiangiogenic Effect of Graphene Oxide in Primary Human Endothelial Cells. ACS Appl. Mater. Interfaces 2020, 12, 22507–22518. [Google Scholar] [CrossRef] [PubMed]
- Dabrowski, B.; Zuchowska, A.; Kasprzak, A.; Zukowska, G.Z.; Brzozka, Z. Cellular uptake of biotransformed graphene oxide into lung cells. Chem.-Biol. Interact. 2023, 376, 110444. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, P.; Xie, C.; Voyiatzis, E.; Faserl, K.; Chetwynd, A.J.; Monikh, F.A.; Melagraki, G.; Zhang, Z.; Peijnenburg, W.J.G.M.; et al. Defining the Surface Oxygen Threshold That Switches the Interaction Mode of Graphene Oxide with Bacteria. ACS Nano 2023, 17, 6350–6361. [Google Scholar] [CrossRef]
- Mari, E.; Mardente, S.; Morgante, E.; Tafani, M.; Lococo, E.; Fico, F.; Valentini, F.; Zicari, A. Graphene Oxide Nanoribbons Induce Autophagic Vacuoles in Neuroblastoma Cell Lines. Int. J. Mol. Sci. 2016, 17, 1995. [Google Scholar] [CrossRef]
- Cucci, L.; Naletova, I.; Consiglio, G.; Satriano, C. A Hybrid Nanoplatform of Graphene Oxide/Nanogold for Plasmonic Sensing and Cellular Applications at the Nanobiointerface. Appl. Sci. 2019, 9, 676. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kim, J.-H. Graphene Oxide–Silver Nanoparticles Nanocomposite Stimulates Differentiation in Human Neuroblastoma Cancer Cells (SH-SY5Y). Int. J. Mol. Sci. 2017, 18, 2549. [Google Scholar] [CrossRef]
- Ghashghaei, H.T.; Lai, C.; Anton, E.S. Neuronal migration in the adult brain: Are we there yet? Nat. Rev. Neurosci. 2007, 8, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Smit, M.; Leng, J.; Klemke, R.L. Assay for neurite outgrowth quantification. Biotechniques 2003, 35, 254–256. [Google Scholar] [CrossRef] [PubMed]
- Pool, M.; Thiemann, J.; Bar-Or, A.; Fournier, A.E. NeuriteTracer: A novel ImageJ plugin for automated quantification of neurite outgrowth. J. Neurosci. Methods 2008, 168, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.M. Neurotrophins: Neurotrophic modulation of neurite growth. Curr. Biol. 2000, 10, R198–R200. [Google Scholar] [CrossRef]
- Ahmed, Z.; Mazibrada, G.; Seabright, R.J.; Dent, R.G.; Berry, M.; Logan, A.; Ahmed, Z.; Mazibrada, G.; Seabright, R.J.; Dent, R.G.; et al. TACE-induced cleavage of NgR and p75 NTR in dorsal root ganglion cultures disinhibits outgrowth and promotes branching of neurites in the presence of inhibitory CNS myelin. FASEB J. 2006, 20, 1939–1941. [Google Scholar] [CrossRef]
- Koprivica, V.; Cho, K.-S.; Park, J.B.; Yiu, G.; Atwal, J.; Gore, B.; Kim, J.A.; Lin, E.; Tessier-Lavigne, M.; Chen, D.F.; et al. EGFR Activation Mediates Inhibition of Axon Regeneration by Myelin and Chondroitin Sulfate Proteoglycans. Science 2005, 310, 106–110. [Google Scholar] [CrossRef]
- Zhang, W.; Duan, W.; Cheung, N.S.; Huang, Z.; Shao, K.; Li, Q.-T. Pituitary adenylate cyclase-activating polypeptide induces translocation of its G-protein-coupled receptor into caveolin-enriched membrane microdomains, leading to enhanced cyclic AMP generation and neurite outgrowth in PC12 cells. J. Neurochem. 2007, 103, 1157–1167. [Google Scholar] [CrossRef]
- Rodriguez-Losada, N.; Wendelbob, R.; Ocaña, M.C.; Casares, A.D.; Guzman de Villoría, R.; Aguirre Gomez, J.A.; Arraez, M.A.; Gonzalez-Alegre, P.; Medina, M.A.; Arenas, E.; et al. Graphene Oxide and Reduced Derivatives, as Powder or Film Scaffolds, Differentially Promote Dopaminergic Neuron Differentiation and Survival. Front. Neurosci. 2020, 14, 570409. [Google Scholar] [CrossRef]
- Lv, M.; Zhang, Y.; Liang, L.; Wei, M.; Hu, W.; Li, X.; Huang, Q. Effect of graphene oxide on undifferentiated and retinoic acid-differentiated SH-SY5Y cells line. Nanoscale 2012, 4, 3861–3866. [Google Scholar] [CrossRef]
- Rizvanov, A.; Idrisova, K.; Zeinalova, A.; Masgutova, G.; Bogov, A.; Allegrucci, C.; Syromiatnikova, V.; Salafutdinov, I.; Garanina, E.; Andreeva, D.; et al. Application of neurotrophic and proangiogenic factors as therapy after peripheral nervous system injury. Neural Regen. Res. 2022, 17, 1240. [Google Scholar] [CrossRef]










| Name | Sequence | MW (g/mol) |
|---|---|---|
| BDNF(1-12) | HSDPARRGELSVK(5,6-FAM)-NH2 | 1680.77 |
| NT3(1-13) | YAEHKSHRGEYSVK(5-FAM)-NH2 | 1920.00 |
| NGF(1-14) | SSSHPIFHRGEFSVK(5-FAM)-NH2 | 1944.06 |
| Sample | E (%) 1 |
|---|---|
| BDNF(1-12)−GO | 12.0 |
| NT3(1-13)−GO | 87.5 |
| NGF(1-14)−GO | 56.7 |
| Pep−GO | E (%) | ||
|---|---|---|---|
| BDNF(1-12) | 1.66 | 32 | 6.16 |
| NT3(1-13) | 1.26 | 10 | 1.64 |
| NGF(1-14) | 1.38 | 44 | 7.84 |
| Sample | D ± Standard Deviation (μm2/s) 1 | 2 |
|---|---|---|
| SLB | 2.7 ± 0 | 0.84 |
| SLB/GO | 2.7 ± 0.1 | 0.75 |
| SLB/BDNF(1-12)−GO | 2.7 ± 0.2 | 0.82 |
| SLB/NT3(1-13)−GO | 3.7 ± 2.3 | 0.10 |
| SLB/NGF(1-14)−GO | 1.3 ± 0 | 0.40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Redigolo, L.; Sanfilippo, V.; La Mendola, D.; Forte, G.; Satriano, C. Bioinspired Nanoplatforms Based on Graphene Oxide and Neurotrophin-Mimicking Peptides. Membranes 2023, 13, 489. https://doi.org/10.3390/membranes13050489
Redigolo L, Sanfilippo V, La Mendola D, Forte G, Satriano C. Bioinspired Nanoplatforms Based on Graphene Oxide and Neurotrophin-Mimicking Peptides. Membranes. 2023; 13(5):489. https://doi.org/10.3390/membranes13050489
Chicago/Turabian StyleRedigolo, Luigi, Vanessa Sanfilippo, Diego La Mendola, Giuseppe Forte, and Cristina Satriano. 2023. "Bioinspired Nanoplatforms Based on Graphene Oxide and Neurotrophin-Mimicking Peptides" Membranes 13, no. 5: 489. https://doi.org/10.3390/membranes13050489