
A Guide to Your Desired Lipid-Asymmetric Vesicles
1
Department of Pharmaceutical Technology and Biopharmacy, Institute for Pharmaceutical Sciences, University of Freiburg, 79104 Freiburg im Breisgau, Germany
2
Leslie Dan Faculty of Pharmacy, University of Toronto, Toronto, ON M5S 3M2, Canada
3
Signalling Research Centers BIOSS and CIBSS, University of Freiburg, 79085 Freiburg im Breisgau, Germany
*
Authors to whom correspondence should be addressed.
Membranes 2023, 13(3), 267; https://doi.org/10.3390/membranes13030267
Submission received: 31 January 2023
/
Revised: 14 February 2023
/
Accepted: 18 February 2023
/
Published: 23 February 2023
(This article belongs to the Special Issue Advances in Symmetric and Asymmetric Lipid Membranes)
Abstract
:Liposomes are prevalent model systems for studies on biological membranes. Recently, increasing attention has been paid to models also representing the lipid asymmetry of biological membranes. Here, we review in-vitro methods that have been established to prepare free-floating vesicles containing different compositions of the classic two-chain glycero- or sphingolipids in their outer and inner leaflet. In total, 72 reports are listed and assigned to four general strategies that are (A) enzymatic conversion of outer leaflet lipids, (B) re-sorting of lipids between leaflets, (C) assembly from different monolayers and (D) exchange of outer leaflet lipids. To guide the reader through this broad field of available techniques, we attempt to draw a road map that leads to the lipid-asymmetric vesicles that suit a given purpose. Of each method, we discuss advantages and limitations. In addition, various verification strategies of asymmetry as well as the role of cholesterol are briefly discussed. The ability to specifically induce lipid asymmetry in model membranes offers insights into the biological functions of asymmetry and may also benefit the technical applications of liposomes.
1. Aims and Content of This Review
For decades, it has been known that most biological, lipid-bilayer based membranes are asymmetric in containing other lipids in the outer than in the inner membrane leaflet [1]. The considerable effort of an organism to establish, maintain, and adapt this asymmetry implies important biological functions [2]. However, since virtually all model membranes used in biophysical and biochemical studies were symmetric, these functions have remained largely unclear. Over the last few years, this long-term shortcoming has been overcome by a large-scale effort to establish and apply new, asymmetric membrane models.
Our review of this highly dynamic field has two main aims. First, we attempt at compiling all assays and protocols to prepare free-floating, lipid-asymmetric vesicles of the classic two-chain, glycero- or sphingolipids reported so far. Table 1 compiles the impressive number of 72 reports differing in strategy or lipid composition that we were able to find. Second, our paper aims at sorting these strategies and protocols into different principal categories and offering a road map that might help with finding the right protocol for a given purpose. For the sake of keeping this paper short and focused, we excluded other, certainly also very interesting membrane models such as asymmetric black lipid membranes, droplet interface bilayers, supported lipid bilayers, multicompartment vesicles, hybrid polymer-lipid vesicles and plasma membrane vesicles, and we did not list work on other lipidic compounds such as ceramides, gangliosides, lyso-lipids or lipopolysaccharides. Finally, we compile the applications as reported, for example a method demonstrated for asymmetric giant unilamellar vesicles (aGUVs) only, and abstain from speculation whether and how existing methods could be adapted or developed to serve other purposes in the future. Of course, such developments are expectable.
Excellent, alternative reviews that focus on other aspects of the field are available. Some articles address the production and application of GUVs in particular [3,4,5,6,7]. Dimova et al. focused on the preparation of aGUVs, in particular their observation by optical microscopy [3]. Reports about the preparation of aGUVs also include various microfluidic-based technologies [5,6,7,8,9]. Huang et al. described microfluidic emulsification in terms of microfluidic fabrication of single, double, triple or higher-order emulsion drops [8]. Kamiya and coworkers [5,6] discussed several techniques based on microfluidics for GUV formation. They summarized the properties of each method, including effects on encapsulation efficiency, size range and asymmetry of membranes. In addition, they described the formation of complex structures in terms of fabricating artificial cell models [5,6]. Cespedes et al. [10] reviewed the interplay between membrane components and the physical properties of the plasma membrane. Their report is outstanding for its focus on the immunological synapse [10]. London and coworkers mainly reviewed cyclodextrin-based methods for preparing asymmetric liposomes [11,12]. Besides this, Kakuda et al. [11] summarized studies about pore-forming toxins, such as perfringolysin O (PFO), regarding lipid interactions in symmetric and asymmetric vesicles [11]. In another article, studies about the effects of asymmetry on the ability of membranes to form ordered domains are summarized [12]. Scott et al. [13] recently reviewed experimental and computational techniques to study membrane asymmetry. Their focus was on in vitro methods that have advanced the understanding of the plasma membrane, along with molecular dynamics simulations. Different techniques for the fabrication of large and giant vesicles are described, i.e. via Ca2+-ions, enzymes and cyclodextrins. With respect to GUV preparation, i.e. hemifusion and phase-transfer approaches are described [13].
Overviewing the different strategies compiled in Table 1, we state that most start with symmetric vesicles and render them asymmetric in another preparation step. This may be achieved by enzymatic conversion of one lipid species into another one (A), by inducing the flip or flop of a given lipid species to accumulate in one leaflet (B), or by exchanging lipids in the outer leaflet (D). A fundamentally different approach is to assemble the vesicle bilayer from individual monolayers from scratch (C). These four fundamental strategies are pursued by many different protocols which all have their specific requirements, limitations, benefits and drawbacks.
2. Navigating the Preparation of Asymmetric Model Membranes
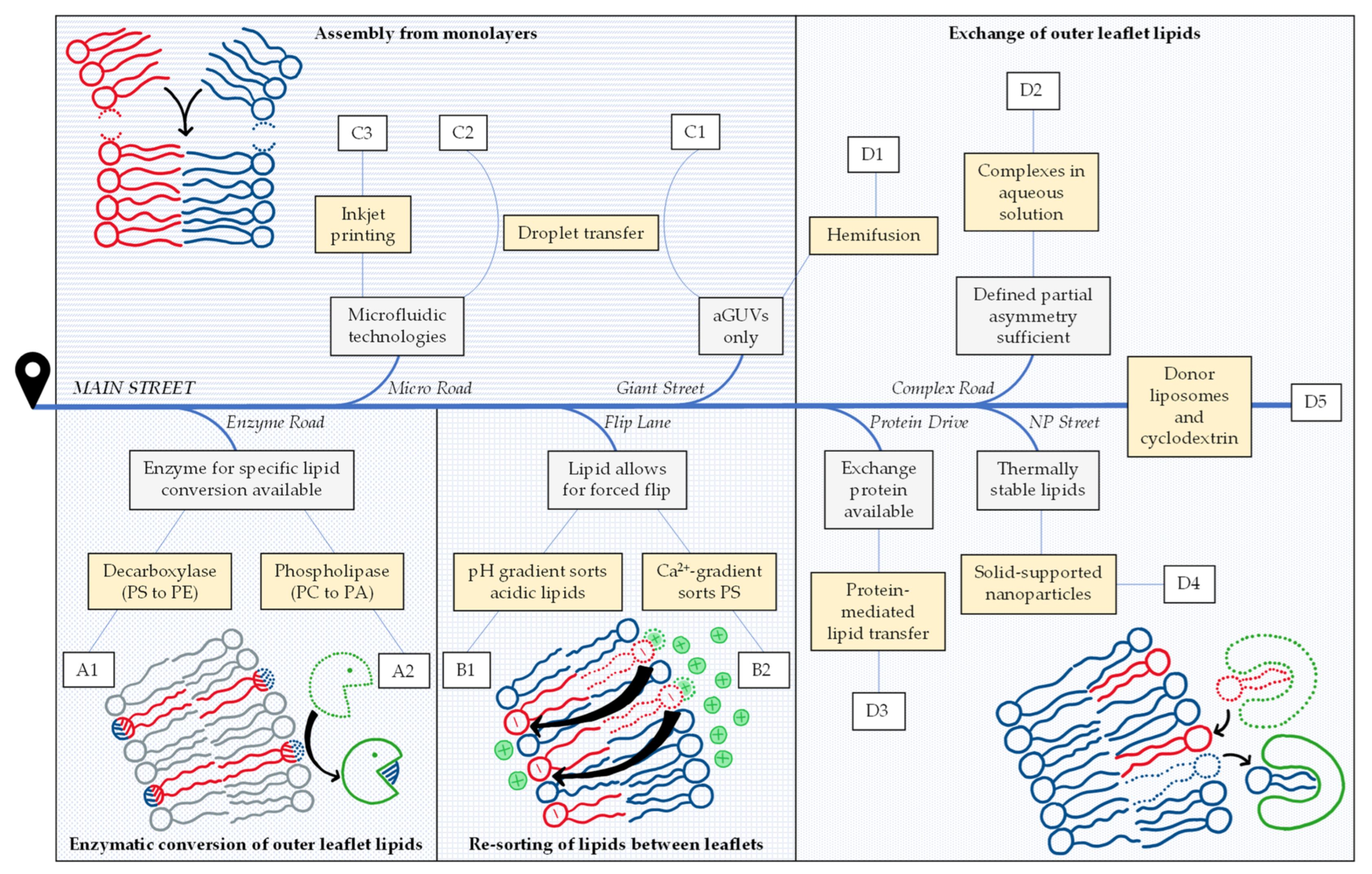
We provide a map to help navigate through this field to find a suitable preparation method that meets individual requirements and possibilities especially in terms of practical implementation (see Figure 1). If you have decided to use asymmetric vesicles but have not made up your mind regarding the specific protocol, “you are here” on the left side of the map and get going down Main Street. On your way, there will be exits to different methods that may or may not be available and favorable for you. As an alternative, you can always stay on Main Street.
For your choice, you will need to rank your options to optimally suit your purpose with respect to the degree of asymmetry, reliability, stability, absence of disturbing components and the time, equipment, materials and expertise needed. Our review cannot solve this problem for you. We are citing some available information on these points given in the original papers but we are lacking a true comparison of different protocols done by the same lab. Furthermore, the best choice depends on the problem and the equipment and experience of a given laboratory. In other words, we try to provide a map but your best path will depend on whether you are driving a racecar, a 4 × 4 or a bicycle.
2.1. Enzymatic Conversion of Outer Leaflet Lipids
The first option to turn off Main Street is taking a right on Enzyme Road. You may take this exit if you are lucky to have an enzyme available that locally (typically in the outer leaflet) converts an undesired lipid into a desired lipid. This is very advantageous for example if you aim at a limited amount of a lipid in the inner leaflet only. Eliminating a minor component in the outer leaflet by unspecific exchange would require the complete replacement of all outer leaflet lipids. The specific elimination of the minor fraction only is much more elegant and less harsh to the vesicles [14]. If such an enzyme is not available or favorable, you may skip this section and stay on Main Street.
To our knowledge, two enzymatic methods have been developed so far, using a decarboxylase (A1) or phospholipase (A2). The approach allows for a minimal invasive formation of asymmetric lipid distributions in the vesicle bilayer, leaving other lipids unaffected [15].
Phosphatidylserine decarboxylase (PSD) converts only phosphatidylserine (PS) localized in the outer leaflet into phosphatidylethanolamine (PE) in only a few steps [16]. We previously described a protocol to produce “20 mol% PS inside” liposomes in only one or two hours, with high but not complete asymmetry and controlled composition that mimics the PS asymmetry of mammalian cell membranes [15].
Phospholipase D (PLD) hydrolyzes phospholipids to phosphatidic acid (PA), whereby the converted lipids can be head group-labeled fluorescent phospholipid analogues [17]. PLD also promotes the transphosphatidylation of phosphatidylcholine (PC) to PS and PE in the presence of serine and ethanolamine [14]. Asymmetric liposomes with about 95% of PC molecules localized to the inner leaflet [14], i.e., 49% conversion of total phospholipids [17] can be produced. The substrate specificity of enzymes, however, limits these methods to specific types of lipids [14]; therefore, it restricts the variety of lipid species that can be asymmetrically distributed in the bilayer. Complete enzymatic lipid conversion presents a difficulty [15], which is why full asymmetry cannot be achieved [14]–[17].
2.2. Re-Sorting of Lipids between Leaflets
The second, rather small lane branching off Main Street to the right is Flip Lane, leading to methods B1 and B2. In the case that a desired lipid allows for forced flip, re-sorting of lipids between leaflets via pH gradients (B1) or via Ca2+-ions (B2) can produce lipid-asymmetric liposomes.
The accumulation of weak acids or bases on one side of a membrane by gradients of pH or complexing agents has long been used to achieve extreme encapsulation efficiencies for the liposomal delivery of water-soluble drugs [18]. A limitation for membrane lipids is that, in contrast to exchanging one lipid for another, a directed transfer of lipids creates an imbalance between the intrinsic areas of the leaflets, i.e., asymmetry stress. If this issue cannot be dealt with specifically, this method must be limited to a very small fraction of a lipid in the membrane.
Weakly acidic lipids can be sorted using the pH gradient method: since lipids may flip across a membrane only in the neutral, but not in charged form, they will accumulate on the low-pH side where they get charged and membrane-impermeant [19]. For anionic lipids, net transport then proceeds from the low-pH side of the bilayer to the high-pH side [20]. This method involves only a few preparation steps such as buffer change, initiation and stop of lipid transport [19,20,21,22,23,24,25]. The pH gradient-induced generation of asymmetric vesicles has been used to modulate membrane fusion [20] and to prepare aGUVs [24]. Certain factors can influence the generation of asymmetry, such as negative surface charge on the membrane, lipid saturation or addition of cholesterol [23]. The amount of lipid transported is limited: for instance, transport of 5% of the total outer leaflet lipid to the inner leaflet has been reported. However, lipid redistribution can occur extremely rapidly [22]. Caution is advised when interpreting the degree of asymmetry specified in such systems. Imagine a vesicle containing 1 mol% of lipid X that is treated in a way to accumulate 95% of this lipid X in the inner leaflet. The resulting vesicle can be referred to as “95% asymmetric with respect to X” but is about 2% asymmetric overall (0.1 mol% of X in outer and 1.9 mol% in inner leaflet) [19,21,22,23,25].
Sun et al. [26] developed a protocol to re-sort PS lipids via ions: the presence of Ca2+-ions combined with an incubation temperature of 70 °C for a certain time allows for controlled production of PS-asymmetric vesicles [26]. The low Ca2+ concentration in the core of the vesicle lets the complex dissociate and entraps the PS. Guo et al. [27] showed that PS flip to the inner membrane leaflet is affected by vesicle size, incubation temperature and lipid composition. Particularly vesicle size and PS content affect the formation of asymmetric lipid distribution, which permits regulating the degree of asymmetry of PS-containing vesicles. Asymmetry remains for days due to a lower activation energy of the flip process compared to the flop process when incubated with Ca2+. However, using vesicles of 400 nm size slows down the formation process of asymmetric vesicles compared to smaller sizes (50 nm). With increasing PS contents of the vesicles, maximal asymmetry decreases [27]. Note that the thermal stability of lipids should be considered. Further, both methods are limited with respect to lipid variety: pH gradients can only induce asymmetric distribution of phospholipids which are weak acids [21], whereas the Ca2+ method is specifically applicable to PS lipids [26].
2.3. Assembly from Monolayers
The first two exits to the left belong to the ‘assembly from monolayers’ county. It is, of course, fascinating to put together a custom-designed asymmetric vesicle directly. One price to pay for this is the involvement of an organic phase that typically leads to more or less organic solvent to remain in the final vesicles (see below). Micro Road is a fairly fast and fancy way to microfluidic technologies which prepare lipid-asymmetric vesicles either via inkjet printing (C3) or via the droplet transfer method (C2). As of today, these instrumentations are not lab standard, so it is a toll road.
Giant Street to droplet transfer will lead you to aGUVs only (C1), but is easier and cheaper to travel. The droplet transfer method originally established by Pautot et al. [28,29] involves, first, the introduction of water droplets into an organic solution of the lipid desired to form the inner leaflet. Spontaneously, a monolayer of lipid covers the water droplets with the chains reaching to the outside (water-in-oil: w/o emulsion). Then, the droplet is forced to cross a boundary from the organic solvent to water, which is covered with the lipid needed for the outer leaflet. The lipid of the surface film will surround the droplet to make its outer surface hydrophilic. Moving micron-sized droplets across the boundary by centrifugation produces aGUVs in the aqueous phase [28]. Using microfluidic technologies, the inner leaflet of the membrane can be prepared by injecting finely tuned water droplets one by one into a flow of a continuous oil phase and then leading them to become surrounded by the water phase [30]. Modifications in microfluidics include layer-by-layer membrane assembly [31], double emulsion [32] and triple emulsion techniques [33], as well as polycarbonate filter systems [34] and dielectrophoretic separation of microemulsions [35]. The inkjet printing method starts from a planar, asymmetric bilayer formed at the contact of two aqueous droplets in an oily phase. From this bilayer, vesicles are ejected by the printing pulse [36,37,38,39,40].
Arriaga et al. [33] summarized various aspects of some of the protocols shown here, including time stability of asymmetry as well as advantages and disadvantages. The production of vesicles via inverse emulsion or droplet phase transfer without using microfluidic devices is easy to implement and leads to high asymmetries up to 95% [33]. Hamada et al. [41] provided a centrifuge-independent method with real-time observation of the transfer process. Vesicle size can be adjusted via sugar gradient [41]. However, the method is limited to low encapsulation and throughput. Vesicle size in general is difficult to control, leading to polydisperse sizes [33]. The phase transfer method is incompatible to lipids that display poor solubility in oil due to their net charge or saturated fatty acid tails [36].
Advantages of microfluidic technologies include high encapsulation efficiency, control over lamellarity and monodisperse vesicle sizes [30,31,32,33,34,35,42,43]. Single-chip microfluidic platforms combining several fabrication steps allow for high-throughput liposome production [32,33,43]. Inkjet printing is also applicable to lipids with poor solubility in oil [36] and achieves long-term stability of at least seven days [37]. However, it requires more specialized equipment than other approaches [36]. Yet, a limited number of solvents can be applied when using poly(dimethylsiloxane) (PDMS) based microfluidic devices [31,32,43].
In general, assembly from different monolayers and control over the composition of each leaflet with or without microfluidic devices leads to high asymmetries up to 100% [28,30,31,32,33,34,35,42]. It enables the encapsulation of macromolecules at any concentration and the use of a wide variety of lipids [28,30,36]. As the lipids are dissolved in organic solvents, oil residues are trapped in the bilayer, possibly affecting membrane properties [28,33,41]. Therefore, some protocols aim to minimize such oil residues [35,36]. If it cannot be avoided to use organic solvents within the vesicle formation procedure, it should be tested if such oil contaminants affect lipid or membrane properties. For instance, Elani et al. [44] studied mechanical properties in terms of vesicle bending rigidities and concluded that the entrapped oil does not influence the above mentioned properties of the membrane [44].
2.4. Exchange of Outer Leaflet Lipids
If none of the exits offered so far turned out to be accessible and particularly attractive, what is left is a set of techniques having in common the exploitation of lipid exchange. As of today, exchange of outer leaflet lipids seems to be the most versatile and widely used strategy able to tackle virtually every lipid asymmetry. Of course, this group of methods does not come without drawbacks and limitations, too. Let us, first, give an overview of the options.
Users aiming at aGUVs who did not exit to the emulsion-based methods (C1, C2) before may go for hemifusion-based exchange (D1) with an excess area of solid supported membranes (see also [45,46,47]). This approach works without a lipid carrier and the elimination of the donor reservoir after exchange is straightforward.
Introducing a defined amount of a component to the outer leaflet of large unilamellar vesicles (LUVs)—such as 20 mol% phosphatidylglycerol (PG) mimicking this asymmetry of bacterial membranes [48]—without the need for donor aggregates has been done by exchange between the liposomes and cyclodextrin-solubilized lipid in aqueous solution (D2). It has also served for forming lipid-asymmetric proteoliposomes [49,50]. An advantage compared to the exchange with donor liposomes or bilayers (D1, D3, D4, D5) is that all of the donor lipid equilibrates very quickly with the outer leaflet of the acceptor vesicles. Application requires knowledge of the cyclodextrin concentration needed to fully solubilize a certain amount of donor lipid.
The lane to protein-mediated lipid transfer (D3) has been little travelled and maintained recently, maybe since cyclodextrins are much simpler and more versatile, but may offer interesting future applications.
Solid-supported vesicles had been used for the TransilTM partitioning assay [51] and to render solid-supported lipid bilayers asymmetric [52]. More recently, they were used as donors for producing lipid-asymmetric liposomes (D4) [53]. In addition to offering an elegant solution to eliminate donors, they also activated exchange without a carrier by increasing the temperature. Naturally, this route should only be travelled for thermally stable lipids and liposomes with asymmetry of sufficient thermal stability.
It reflects our personal view of the current literature of the field that whoever did not have a chance or did not bother to exit Main Street to explore potentially advantageous sideroads will finally cross London Bridge to the protocol of lipid exchange between donor and acceptor vesicles.
Let us address the methods D1-D5 in some more detail. Enoki et al. [45] established the preparation of aGUVs via hemifusion of symmetric GUVs with a solid supported bilayer (SLB). In the hemifusion state induced by the presence of Ca2+, GUVs dock to the support and outer leaflet lipids exchange by diffusion. By eliminating Ca2+ using a chelator, hemifusion is reversed and the aGUVs detach from the SLB [45]. By using the hemifusion method, the resulting aGUVs are free of any exogenous contaminants such as cyclodextrins or organic solvents, except for trace fractions of fluorescence dyes used for detecting asymmetric exchange. Preparation of aGUVs and data collection needs less than five hours. The resulting vesicles show high asymmetries approaching 100%, given the large excess of the SLB donor area over the GUV area [45,46,47]. However, during the hemifusion process or when aGUVs are sheared off the SLB, transient pores are formed so that lipid flip-flop may occur. Such “leaky” GUVs should be identified and excluded from subsequent aGUV experiments [45].
Using solubilized donor lipids (D2) [48] rather than donor vesicles or bilayers has two main advantages. Exchange is very fast and complete, yielding a well-defined content of donor lipid in the target liposomes. Second, the elimination of the cyclodextrin complexes after exchange is either unnecessary, if subsequent experiments are not compromised by ongoing equilibrium exchange, or trivial. This protocol was also utilized for the preparation of lipid-asymmetric proteoliposomes, containing a large, multi-spanning membrane protein, the antiporter ST-NhaA [49] and the ligand-gated ion channel ELIC [50]. By now, this has been demonstrated only with very few procedures [49,50,54,55].The principle of this approach is to first completely dissolve donor liposomes to obtain a solution of mβCD−donor lipid (in our example, PG) complexes. This solution is then equilibrated with a proper amount of “acceptor” liposomes so that PG enters the membrane and the corresponding amount of PC is solubilized instead. To determine the required amounts of lipid and cyclodextrin for the desired degree of lipid exchange, lipid-cyclodextrin interactions were previously investigated by isothermal titration calorimetry (ITC) experiments; an alternative method would be light scattering (see below). Within one “round” of exchange, 5 to 45 mol% lipid were exchanged in the outer vesicle leaflet. The asymmetry of LUVs remained stable for 14 days [48] and of proteoliposomes for seven days [49]. A detailed protocol and ExcelTM sheet is provided to calculate required lipid and mβCD concentrations [48].
Living organisms transport lipids by a variety of specific carrier proteins that may be used to facilitate exchange without the need for cyclodextrins. PC molecules, for instance, were introduced into the outer vesicle monolayer via exchange protein from bovine liver [56]. PC-specific exchange protein was also used to prepare vesicles with an asymmetric distribution of brominated PC molecules. However, brominated PC lipids possibly have adverse effects upon the enzymatic activity of some reconstituted systems in model membranes [57]. Sandra et al. [58] generated PE-asymmetric vesicles by incubating lipid vesicles with rat liver exchange protein and a suitable acceptor membrane. Only the outer surface of PE-containing vesicles is accessible to the exchange protein, which leads to an asymmetric lipid distribution across the bilayer [58]. Holzer et al. [59] initiated protein-mediated lipid transfer between egg-PC (EPC) acceptor vesicles and EPC:EPG 90:10 mol% donor vesicles with the help of recombinant pro-sterol carrier protein 2 (pro-SCP2). Using this protein-mediated strategy for lipid exchange, aLUVs were fabricated in less than three hours. As a result, the amount of EPG in acceptor vesicles increased to 3 mol%, whereas EPG in donor vesicles was reduced to 6 mol%. Pro-SCP2 accelerates the EPG transfer to half-times of between two and three hours, and thus, minimizes lipid flip-flop during the transfer process. In comparison, the spontaneous redistribution of EPG occurs at half-times of tens of hours. Note that liposome size affects the degree of asymmetry. Narrow size distributions are important for obtaining aLUVs with a uniform degree of asymmetry [59]. In general, the application of protein-mediated lipid exchange is limited to a few lipid species. Further, post-exchange acceptor vesicles can be contaminated with donor vesicles [56]. Unless the use of cyclodextrin has to be avoided in a given system or for a given experimental technique, there seems to be little motivation to use a transfer protein in a rather unspecific manner. The true potential of this approach would be to selectively add or remove a component while leaving all others unaffected, as discussed above for enzymatic conversion. However, specific insertion or extraction of a single lipid without creating or filling the “gaps” with another would create asymmetry stress as discussed in the section on induced flip. Reaching substantial asymmetries by selective transfer would require a solution to this problem.
Another possibility to exchange lipids of the outer leaflet is using solid-supported nanoparticles as the donor phase, and a high temperature to activate exchange [53]. Small unilamellar vesicles (SUVs) of desired composition were prepared by adjusting parameters such as temperature, time and ratio of lipid-coated silica nanoparticles to vesicles. The use of lipid-coated nanoparticles facilitates the purification process for the easy preparation and isolation of asymmetric vesicles. Here, lipid exchange proceeds at 75 °C [53]. Note that elevated temperatures activate both the desired lipid exchange and the detrimental intra-bilayer lipid flip-flop in asymmetric liposomes [60]. Hence, the exchange protocol has to find a compromise between these effects: the authors managed to reach a final content of donor lipid, 1,2-dipalmitoyl-sn-glycero-3-phosphocholine (hDPPC), of 20 mol% in the outer compared to 5 mol% in the inner leaflet. The method was demonstrated for saturated, thermally stable, isotopically distinct DPPC lipid molecules [53].
The exchange between donor and acceptor liposomes is a classic and most widely used method established by London and coworkers. The principle of this method is to facilitate the exchange between donor and acceptor vesicles by relatively small concentrations of cyclodextrin which solubilize only a little lipid at a time but shuttle some of it between the vesicles. After exchange, the now-asymmetric acceptor vesicles need to be separated from a potentially large excess of donor vesicles by centrifugation [61,62,63]. Over the last decade, several modifications of this method have been developed, which differ in, for instance, cyclodextrin species, vesicle size and centrifugation procedures. Besides methyl-β-cyclodextrin (mβCD) [55,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77], hydroxypropyl-α-cyclodextrin (HPαCD) [78,79,80,81,82] and methyl-α-cyclodextrin (mαCD) [83,84] was also used. Mainly aLUVs were produced, but some protocols resulted in fabricating aSUVs [61,63,75,76] and aGUVs [55,67,73,75,80]. Two main procedures have been established regarding the centrifugation step for vesicle separation: the heavy-acceptor (ha) and heavy-donor (hd) strategy. Cheng et al. first provided the ha-strategy [61], which was modified later by Heberle et al. [62] in terms of loading donor vesicles instead of acceptor vesicles with sucrose solution. Doktorova et al. [63] provided a detailed protocol, including the two strategies mentioned above. In about 12 h, this protocol can produce up to 20 mg of asymmetric vesicles. Comparing hd- and ha-strategies, the latter simplifies purification, whereas the hd-strategy excludes sucrose from acceptor vesicles. For the hd-strategy, additional purification steps may be required depending on the density of the donor lipid, resulting in reduced yield. Moreover, entrapped sucrose induces osmotic stress, potentially causing bilayer thinning and lipid area expansion [63]. An approach of Li et al. [84] entraps physiological osmolalities of cesium chloride (CsCl) inside acceptor aLUVs instead of sucrose. The density of liposomes is increased without the use of a hypertonic sucrose solution, preventing acceptor vesicles from osmotic pressure imbalance. CsCl entrapment did not interfere with the ability to produce aLUVs or maintain efficient exchange [84].

{kind=link}
Table 1.
Summary of protocols for in-vitro preparation of lipid-asymmetric vesicles. Articles are listed by category (A–D) and date of publication. Selected features are shown including vesicle type, degree of asymmetry (asy), outer leaflet and inner leaflet composition, asymmetry verification method and a short description of the respective article. Lipids primarily intended to be asymmetrically present in one leaflet are highlighted in bold. Lipids in light font are matrix or acceptor lipids that may be present in both leaflets. Note that the degree of asymmetry is interpreted in different ways; thus, footnotes are inserted to provide more detailed information.
Table 1.
Summary of protocols for in-vitro preparation of lipid-asymmetric vesicles. Articles are listed by category (A–D) and date of publication. Selected features are shown including vesicle type, degree of asymmetry (asy), outer leaflet and inner leaflet composition, asymmetry verification method and a short description of the respective article. Lipids primarily intended to be asymmetrically present in one leaflet are highlighted in bold. Lipids in light font are matrix or acceptor lipids that may be present in both leaflets. Note that the degree of asymmetry is interpreted in different ways; thus, footnotes are inserted to provide more detailed information.
| Vesicle Type | Asy | Outer Leaflet | Inner Leaflet | Verification of Asy | Short Description | Ref. |
|---|---|---|---|---|---|---|
| A. Enzymatic conversion of outer leaflet lipids | ||||||
| A.1. Decarboxylase | ||||||
| LUV | 97% 1 | DOPC/NBD-PE | NBD-PS | FRET, trinitrophenylation | one-step method, enzyme conversion of PS to PE by PS-decarboxylase | [16] |
| LUV | a = −0.5 (PS), a ≈ 1 (PE) 2 | ePC/PE PC/chol/eSM/PE PC/PE PC/PE/PG/PE | POPS POPG POPS/POPG | ζ-potential, HPTLC | PS-decarboxylase converts PS to PE, aLUVs mimic PS-asymmetry of eukaryotic plasma membranes | [15] |
| A.2. Phospholipase D | ||||||
| LUV | 49% 1 | PA | PC/PE/N-NBD-PE/N-Rho-PE | F (N-Rho-PE, N-NBD-PE) | outer lipid conversion to PA, influenza-induced fusion between viral and liposome membrane | [17] |
| LUV | >95% 3 | POPS/POPE | POPC± chol | enzymatic assay/optical absorption, HPLC | enzymatic conversion of PC in the presence of serine and ethanolamine | [14] |
| B. Re-sorting of lipids between leaflets | ||||||
| B.1. pH gradient | ||||||
| LUV | 80–90% 10 | DOPC | ePG DOPA | ion-exchange C, 13C NMR, periodate oxidation | asymmetric distributions of PA in aLUVs via pH gradients | [21] |
| LUV | 50% 13 | DPPC DPoPC DOPC ePC ePC/chol ePC/PS | ePG DOPG MOPG | periodate oxidation | mechanism of pH-induced PG trans-bilayer transport | [22] |
| LUV | >80% | PA PC CL | PC PG SA | two-phase polymer partition, 3H-radioactivity | effect of temperature and lipid composition on formation and extent of asymmetry | [23] |
| LUV | >95% 13 | DOPA DOPE/DOPC/PI | DOPE/DOPC/PI DOPA | F (TNS) | influence of lipid asymmetry on Ca2+-stimulated vesicles fusion | [19] |
| GUV | n.a. | ePG | ePC | phase contrast M | influence of lipid redistribution on the shape of GUVs | [24] |
| LUV | n.a. | ePC/chol ePC/DOPE/chol | amino lipids AL1-AL6 | F (TNS) | pH gradient induced fusion of liposomes containing synthetic amino lipids | [20] |
| LUV | >80% 13 | DOPC ePC | DOPA ePA | NMR | NMR observation on transbilayer distribution of Chlorpromazine | [25] |
| B.2. Ca2+-ions | ||||||
| LUV | ≤30% 13 | DPPC | DOPS | FQ (NBD-PS), nanoDSC | Ca2+-induced inward flip of PS for controlled production of aLUVs | [26] |
| LUV | 38.5–52.3% 14 | DPPC | DOPS | FQ (NBD-PS), nanoDSC | effect of size, temperature and lipid composition on Ca2+-induced PS inward flip | [27] |
| C. Assembly from monolayers | ||||||
| C.1. Droplet transfer/emulsion phase transfer | ||||||
| GUV | ≤95% | POPC ePC | POPS polystyrene-polyacrylic acid | FQ (NBD-PE, NBD-PS) | engineering aGUVs with two independently prepared monolayers | [28] |
| GUV | n.a. | POPC/py-16-PC POPC/POPE/CL | POPC/POPE/CL POPC/py-16-P | F (pyrene) | membrane − protein interactions between Bax and liposomes of size 0.3-1.5 µm | [85] |
| GUV | n.a. | DOPC | ePC DOPC/DPPC/chol | FM (Rho-PE, NBD-PE) | cell-sized aGUVs, control over vesicle size via sugar gradient | [41] |
| GUV | n.a. | ePC DOPG | DOPE ePC DOPG | n.a. | reconstitution of the potassium channel KcsA into aGUVs | [54] |
| GUV | n.a. | POPC DOPC | DOPC POPC | FM of hemifused GUVs (Rho-PE, NBD-PE) | effects of lipid asymmetry on membrane bending rigidity | [44] |
| GUV | n.a. | DOPC/chol NBPC/chol | NBPC/chol DOPC/chol | FM (Rho-DHPE) | asymmetric distribution of photocleavable lipid, photoinduced pinocytosis behaviour | [86] |
| GUV | n.a. | DSPE/DSPG | DSPG DSPE DOPG DOPC | FQ (NBD-PE) | influence of lipid head group and acyl chain on Daptomycin-induced membrane permeability | [87] |
| GUV | n.a. | DOPC DOPC/DOPS DOPC/DOPG | DOPC/DOPS DOPC/DOPG DOPC | F Annexin V (Alexa Fluor 488) | protein translocation via cell-penetrating peptides, start of enzymatic reactions in aGUVs | [88] |
| C.2. Droplet transfer/microfluidic technologies | ||||||
| GUV | 85% | DPPC DOPC PS | DPPC DOPC PS | FQ (Texas Red (TR)-modified DPPE), biotin-binding (biotin-DPPE, avidin), F Annexin V (Alexa Fluor 488) | two-step fabrication of monodisperse and unilamellar aGUVs | [30] |
| GUV | 100% | NBD-DOPC | DOPC | F (NBD-DOPC) | controlled construction of uni- or multilamellar aGUVs using layer-by-layer membrane assembly | [31] |
| GUV | 90–95% | DOPC | DOPE | FM, FQ (NBD-DOPC, TR-DOPE) | continuous fabrication of aGUVs via double emulsions with customized membrane composition, size and luminal content | [32] |
| GUV | 95% | DMPC DOPC | DOPC DMPC | FQ (NBD-PC) | influence of asymmetry on area expansion modulus, customized micropipette aspiration system | [42] |
| GUV | n.a. | DOPC | POPC | F click chemistry (DSPE-DBCO, 3-azido-7-hydroxycoumarin) | high-throughput fabrication of aGUVs from aqueous lipid dispersions | [43] |
| GUV | ≤70% | DOPC DOPE-biotinyl | DOPC DOPE-biotinyl/DOPC | F/biotin-streptavidin (DOPE-biotinyl, streptavidin fluorescein isothiocyanate ST-FITC) | continuous single-step fabrication in a glass device using triple emulsion drops | [33] |
| GUV, LUV | 79% | POPS | POPC | FM (carboxy-fluorescein), FQ (NBD-PC) | preparation of liposomes with desired diameters using a tunable microfluidic device including a polycarbonate filter | [34] |
| GUV | 83% | POPC | DOPC | FQ (NBD-DHPE) | aGUVs with precisely modulated size and minimal oil contamination | [35] |
| C.3. Inkjet printing | ||||||
| GUV | n.a. | DPhPC/TMR-PIP2 PE-PEG2000 DPhPC/DGS-NTA-Ni DPhPC DPhPC/DPhPS/chol | DPhPC/TMR-PIP2 PE-PEG2000 DPhPC/DGS-NTA-Ni DPhPC DPhPC/DPhPS/chol | FM (TMR-PIP2, His-GFP) | separate vesicle and bilayer formation allows for monitoring and minimizing oil contamination | [36] |
| GUV | n.a. | DOPC DOPS | DOPC DOPS DOPS/DOPE/DOPC | F Annexin V (Alexa Fluor 488, 546) | membrane dynamics and protein interactions, use of little organic solvent | [37] |
| GUV | n.a. | DOPC DOPC/DOPE | DOPC/DOPS | FQ (Rho-DOPE) | device for sequentially generating aGUVs, influence of the peptide Cinnamycin on lipid dynamics | [38] |
| LUV | n.a. | DOPC biotin-DOPE | biotin-DOPE DOPC | Biotin-streptavidin (biotin-PEG(2000)-DSPE, Streptavidin-conjugated gold) and TEM, FM (Rho-DOPE) | fabricating nano-sized liposomes from a planar lipid bilayer by applying a pulsed-jet flow with optimized duration and pressure | [39] |
| GUV | n.a. | DOPC DOPC/DOPS | DOPC/DOPS DOPC/DOTAP | FM (Rho-DOPE) | fusion between LUVs and a monolayer, followed by application of a pulsed jet flow | [40] |
| D. Exchange of outer leaflet lipids | ||||||
| D.1. Hemifusion | ||||||
| GUV | 50–99% 12 | DOPC/chol | DSPC | FM (TRPC, DiD), FQ (NBD-PE) | hemifusion of giant vesicles and a supported lipid bilayer | [45] |
| GUV | ≤86% 12 | DOPC/chol | DSPC/POPC | FM (TRPC, DiD) | systematic study of aGUVs to investigate modulated phases | [46] |
| GUV | >70% 12 | DOPC DOPC/chol | DSPC bSM DSPC | FM (TRPC, DiD) | phase behavior and cholesterol movement in aGUVs | [47] |
| D.2. Complexes in aqueous solution | ||||||
| LUV | 0.05–0.45 9 | POPG | POPC | ζ-potential | PG-loaded mβCD-lipid-complexes in solution replace PC by PG | [48] |
| LUV | 0.2 9 | POPG | POPE/POPC/TOCL | ζ-potential | five-step protocol to proteoliposomes with incorporated ST-NhaA | [49] |
| LUV | 0.25 9 | POPG | POPC POPC/POPE | ζ-potential | phospholipid modulation of ELIC in PG-asymmetric proteoliposomes | [50] |
| D.3. Protein-mediated lipid transfer | ||||||
| SUV | 10–20% 4 | [N-13CH3]-DOPC [N-13CH3]-DMPC | DMPC DOPC | 13C-NMR | transfer of PC between acceptor and donor PC vesicles | [56] |
| MV | 62% 4 | [3H]-DOPC [3H]-DOPE | DOPC/DOPE/CL | radioactivity, TNBS-labeling | lipid transfer between isotopically asymmetric vesicles and chinese hamster fibroblasts | [58] |
| SUV | ≤59.1% 4 | BRPC POPC | POPC BRPC | F (CUGA), GC | studies on the membrane-binding domain of cytochrome b5 in brominated aSUVs | [57] |
| LUV | 60% 13 or 3 mol% 5 | ePG | ePC | FFE | pro-SCP2 mediated EPG transfer, separation of donor and acceptor vesicles via FFE | [59] |
| D.4. Solid-supported nanoparticles | ||||||
| SUV | 75.6 mol% 5, 24.4 mol% 10 | hDPPC | d62DPPC d75DPPC | SANS, 1H-NMR | lipid exchange via lipid-coated silica nanoparticles | [53] |
| D.5. Donor liposomes and cyclodextrin | ||||||
| SUV | 75–82% 4 | bSM | DOPC POPC POPS/POPE DOPC/chol POPE/POPS/chol | FA (DPH, TMA-DPH), HPTLC, pL4A18 peptide binding | mβCD-mediated lipid exchange, ha-strategy | [61] |
| GUV | 60% 4 | bSM | DOPC bPC bPC/bPE ± chol | FCS (Nile-red NR12S, NBD-PE) | solvent free method for mβCD-induced lipid exchange to prepare aGUVs, ha-strategy | [73] |
| LUV | 80–100% 4 | bSM bSM/POPC | DOPE/POPS ± chol | FA (DPH, TMA-DPH), HPTLC, pL4A18 peptide binding | mβCD-mediated exchange (ha-strategy), investigation of interleaflet coupling | [74] |
| GUV, SUV | 20–80 mol% 4 | bSM mSM C24:0-SM/bSM | DOPC POPC/bSM SOPC/bSM OMPC/bSM | FLIM (NBD-DPPE, NBD-DOPE, TMA-DPH) | mβCD-induced exchange (ha-strategy) with lipids of various acyl chains, investigation of interleaflet coupling | [75] |
| SUV | 62–96% 4 | bSM | di14:1PC di16:1PC di18:1PC di20:1PC di22:1PC diphyPC 16:01-18:2PC 16:0-20:4PC di18:2PC di18:3PC di20:4PC | FA (TMA-DPH), HPTLC | mβCD-mediated lipid exchange (ha-strategy), effects of PC acyl chain structure | [76] |
| LUV | >90% 4 | bSM PC bSM/PC | PE/PS/chol | HPTLC, pL4A18 peptide binding, TNBS-labeling | HPαCD-mediated exchange (ha-strategy) with controlled amount of cholesterol (0-50 mol%) | [78] |
| LUV | 80–90% 4 | POPC | POPE/POPS/chol | HPTLC, pL4A18 peptide binding, TNBS-labeling | HPαCD-mediated exchange (ha-strategy), influence of lipid composition and asymmetry on Perfringolysin O | [79] |
| GUV, LUV | 50 mol% 5 | eSM mSM | DOPC/chol | F (Rho-DOPE, NBD-DOPE), FA (TMA-DPH), HPTLC | solvent-free method for HPαCD-induced lipid exchange (ha-strategy) and control of cholesterol | [80] |
| LUV | n.a. | bSM | DOPE/POPS | F (Topfluor-PC) | mβCD-induced exchange (ha-strategy), antibody-decorated aLUVs bind HIV-1 virus-like particles | [77] |
| LUV | ≤0.95 6 | POPC-dHC POPC DPPC-dC | POPC POPC-dH POPC-dHC | GC-MS, 1H-NMR, SANS | solvent-free and sucrose-free aLUVs prepared via mβCD-mediated exchange, hd-strategy | [62] |
| GUV | n.a. | bSM 24:1-SM 16:0-SM 18:0-SM | POPC/SM POPC ± chol | TLC, FA (DPH, TMA-SPH) | mβCD-induced exchange (ha-strategy), influence of lipid composition on AChR distribution in symmetric and asymmetric liposomes | [55] |
| LUV | n.a. | py-PG py-PG py-PI | POPC | F (pyrene) | kinetic analysis of mβCD-mediated exchange via real-time monitoring of intervesicular lipid transfer | [64] |
| LUV | 60% 4 | DPPC DPPC-d62 POPC-d44 POPC | POPC POPC-d44 POPC-d31 | GC-MS, UPLC-MS, 1H-NMR, SANS | SANS and SAXS analysis of aLUVs prepared via mβCD-induced lipid exchange, hd-strategy | [65] |
| LUV | 59% 4 | DPPC-dC | DPPC-dH | 1H-NMR, GC | mβCD-mediated exchange (hd-strategy), lipid flip-flop in gel and fluid bilayers | [60] |
| LUV | 0.48–0.67 7 | POPC POPE | POPE POPC | DSC, UPLC-MS, 1H-NMR | mβCD-induced exchange (hd-strategy), leaflet-specific lipid packing and melting | [66] |
| LUV, SUV | 0.34–0.45 8 | DMPC-d54 eSM DPPC DPPC | POPC-d13 POPC POPE ± chol | GC-MS, 1H-NMR | detailed protocol for the preparation of asymmetric vesicles via mβCD-mediated lipid exchange | [63] |
| LUV | ≤85.9 4 | mSM bSM eSM DMPC DPPC diC(15:0)PC DSPC | DOPC ± chol | 1H-NMR, HPTLC, FQ (Rho-DMPE, Rho-DOPE) | HPαCD-induced lipid exchange (ha-strategy), domain formation and interleaflet coupling using FRET | [81] |
| GUV, LUV | 30–40% 4 | C24-SM C18-SM C16-SM C16-SM/C14-SM PC | DPPC/DOPC/chol POPC/POPS/POPE POPC/POPS/DOPE | FM (NBD-DPPE, Rho-DPPE), MS FA (TMA-DPH) | mβCD-induced exchange (ha-strategy), influence of C24 sphingolipids on cholesterol and membrane microdomains | [67] |
| LUV | ≤75% 4 | bSM | DOPC sterol (chol, epichol, lanosterol, 7-dehydrochol, 4-cholesten-3-one) | FRET (Rho-DOPE, DPH), HPTLC | HPαCD-induced exchange (ha-strategy), incorporation of different sterol structures into aLUVs | [82] |
| LUV | 0.32–0.45 9 | POPC-d31 DMPC-d54 | POPC POPC-d13 | GC-MS, 1H-NMR | mβCD-mediated exchange (hd-strategy), influence of Gramicidin on lipid flip-flop and membrane- protein interactions | [68] |
| LUV | 0.44 9 | eSM | POPE | 1H-NMR, 31P-NMR | mβCD-induced lipid exchange (hd-strategy), studies of bending fluctuations via neutron spin-echo spectroscopy | [69] |
| LUV | 7 mol% 10, 3 mol% 5 | POPC-d31 | POPS | GC-MS, F Annexin V assay (Annexin V-568) | mβCD-mediated exchange (hd-strategy), influence of PS asymmetry on the membrane interaction of pHLIP | [70] |
| LUV | ≤100% 8 | POePC DOTAP POPS POPG POPA | POPC POePC/POPC DOTAP/POPC POPS/POPC POPG/POPC POPA/POPC ± chol | HPTLC, F (DPH, TMA-DPH) | mαCD-induced lipid exchange (ha-strategy), entrapment properties of aLUVs containing one cationic and/or anionic leaflet | [83] |
| LUV | 41–96% 11 | POPE/TOCL | POPC | TNBS-labeling F (TTAPE-Me) | mβCD-mediated exchange with donor-SUVs instead of MLVs, effect of lipid asymmetry on MOM permeabilization by apoptotic proteins (tBid/Bax) | [89] |
| LUV | 73% 4 | POPC | POPE/POPS/chol | TLC, F (TMA-DPH) | mαCD-induced lipid exchange with CsCl entrapped in aLUVs | [84] |
| LUV | n.a. | POPE | POPG POPG-d31 | UPLC-MS, GC | mβCD-mediated exchange (hd-strategy), interactions between aLUVs and frog peptides (L18W-PGLa, MG2a) or lactoferricin derivative LF11-215 | [71] |
| LUV | 55–70% 4 | MSPC SMPC PMPC MSM POPC SOPC | DPPC | GC, SANS | mβCD-induced exchange (hd-strategy), transleaflet coupling of aLUVs in the fluid phase | [72] |
1 Lipid conversion (in the outer leaflet); 2 a = asymmetry parameter, ranges from −1 (all-inside localization of PS) via 0 (symmetric distribution) to 1 (outside-only PS) [15]; 3 Total PC molecules located to the inner leaflet; 4 Outer leaflet lipids exchanged; 5 Amount of destined lipid located in the outer leaflet; 6 Component fraction in the outer versus inner leaflet; 7 ∑as = degree of asymmetry, mole fractions of donor lipid in the outer and inner bilayer leaflets [66]; 8 Exchange efficiency: fraction of outer leaflet exchange [63]; 9 Mole fraction of incorporated lipid in the outer leaflet; 10 Amount of destined lipid located in the inner leaflet; 11 Lipid molar ratio between outer leaflet and total individual lipid content; 12 Pa% = percentage of lipid exchange in the asymmetric vesicle [45,46,47]; 13 Lipid transported; 14 a = asymmetric degree, based on PS monolayer concentration in the inner and outer leaflet [27]; Acronyms: C chromatography, F fluorescence, FA fluorescence anisotropy, FM fluorescence microscopy, FQ fluorescence quenching, FCS fluorescence correlation spectroscopy, FFE free-flow electrophoresis, FLIM fluorescence lifetime imaging, FRET Förster resonance energy transfer, GC gas chromatography, HPTLC high-performance thin-layer chromatography, M microscopy, MS mass spectrometry, NMR nuclear magnetic resonance spectroscopy, SANS small angle neutron scattering, TLC thin layer chromatography, UPLC ultra-performance liquid chromatography.
The method can be fine-tuned to fabricate bilayers of different intended compositions including a variety of acyl chain structures as well as lipid mixtures and cholesterol [61,73,74,76]. Note that asymmetric vesicles are hardly formed from lipids with two short or polyunsaturated acyl chains due to transverse diffusion [76]. Any change in lipid composition should first be examined with dynamic light scattering (DLS)to determine the mβCD concentration at which vesicles are dissolving [63]. Lin et al. [78] used HPαCD instead of mβCD to improve vesicle yields and control the amount of cholesterol introduced. However, difference in affinity of HPαCD for various lipids may lead to less efficient exchange [78]. Exchange efficiencies, defined as the fraction of outer leaflet exchange and calculated in consideration of the donor mole fraction, are about 0.35–0.45 [63]. If higher efficiency is desired, the donor/acceptor ratio can be increased, or multiple rounds of exchange can be performed. However, additional exchange steps may reduce vesicle yield [63]. In contrast, the presence of cholesterol significantly improves vesicle yield in many cases. Further, lipid charge affects exchange efficiency: using donor vesicles containing cationic phospholipids results in lower exchange efficiency, compared to using donor vesicles containing anionic phospholipids [83]. Donor lipid contamination can occur in the inner leaflet of acceptor vesicles in some cases [60,63]. Despite separation, residual aqueous cyclodextrin is still found in the system but has no measurable effects on lipid motion within the bilayer [60].
The extent of asymmetric insertion of a lipid from donor liposomes or bilayers (D1, D3, D4, D5) depends on time and the number, accessibility and composition of the donors. Let us, for example, assume one is aiming at exchanging 50 mol% of PC for PG in the outer leaflet of acceptor liposomes. One principal way would be to equilibrate the acceptor liposomes with the proper amount of donor liposomes. In our example, to reach 50% donor lipid, the accessible, outermost leaflet of the donors would need to include the same amount of lipid as that of the acceptor. Alternatively, one could equilibrate acceptors with a large excess of 50 mol%-PG donors. The rate of equilibration increases with cyclodextrin concentration (D5), donor curvature (D4) and temperature. If equilibration is too slow to be completed, more donor would be needed for the same result.
3. Testing Asymmetry
Establishing and, to a large degree, safely employing a protocol to establish lipid asymmetry requires a means to quantify asymmetry. The choice of a validation method is not connected to the preparation protocol and hence, not within the scope of this review. But to give an overview, we will briefly take a look at the different strategies.
An ideal method to quantify the transmembrane distribution of a single charged lipid is the precise measurement of the zeta potential. It utilizes the fact that the low dielectric permittivity of the membrane core renders the charges in the inner leaflet invisible without any additional quenching or labeling as needed for other methods. Zeta potential measurement provides a label-free and non-destructive assay for asymmetry verification. It should be noted that experience and non-standard procedures or accessories such as high-concentration or dip cells might be needed to ensure that the zeta measurement reaches the necessary precision.
Fluorescence is a very versatile technique and fluorescence can be quenched selectively in the outer leaflet. However, usually high amounts of quencher must be used, resulting in increased osmolarities outside the liposomes. Annexin-V assay is suitable for specific quantification of PS lipids in the outer leaflet. It is a very sensitive technique but the sample cannot be used for further analysis. Fluorescence anisotropy can be used for studies of lipid order. Other options include fluorescence microscopy, which is suitable solely for testing GUVs.
It should be noted that the presence of fluorophores attached to lipids changes some of their properties crucially. This limits the applicability to asymmetric vesicles. For example, it essentially disqualifies fluorescent lipids from being proper models of unlabeled lipids for flip-flop studies.
NMR with shift reagents detects the fraction of a lipid species that is accessible from outside. Scattering techniques (SANS, SAXS) require or profit from deuterium labelling, which should be less intrusive to membrane properties than fluorophores. Mass spectrometry, optionally combined with other analytical methods such as gas chromatography, enables the analysis of molar masses, i.e. isotopically asymmetric labelled lipids. However, destruction of the sample has to be accepted.
4. Cholesterol
A membrane component outside the defined scope of lipids discussed here but of key interest for model membranes is cholesterol. Biological membranes, such as erythrocyte membranes, contain considerably larger amounts of cholesterol in their outer leaflet. Unlike the phospholipids discussed so far, cholesterol undergoes a fast flip-flop across the membrane so that its distribution is essentially equilibrated. Asymmetry is imposed by the facts that (I) the outer leaflet contains more unsaturated (sphingo)lipids with high cholesterol affinity than the inner and (II) the outer leaflet contains lesser phospholipid (intrinsic area) all together, giving way to the area requirement of additional cholesterol.
Together with mixing entropy opposing asymmetry, these properties give rise to cholesterol asymmetry [90,91]. It appears that creating sphingomyelin- and area asymmetry would be a means to establish a proper cholesterol asymmetry in a model membrane.
Two strategies should be possible to avoid cholesterol to interfere with cyclodextrin-based protocols for phospholipid asymmetry. First, α-cyclodextrins may be more challenging for lipid transfer but do not complex cholesterol. Hence, lipids can be handled selectively in the presence of cholesterol. Second, β-cyclodextrins bind cholesterol primarily with a stoichiometry of 2:1 but phospholipids with 4:1; given the binding constants, it needs relatively low cyclodextrin concentrations (of the order of 5–10 mM mβCD) to transport cholesterol [92] but higher cyclodextrin concentrations of about 30–50 mM to extract significant amounts of lipid [93]. Hence, one could in principle deal with the lipids first, at high cyclodextrin, and then add cholesterol using low cyclodextrin. The issue remains, though, that a relaxed lipid membrane with matching intrinsic areas of the outside and inside lipids will also incorporate cholesterol in a largely symmetrical manner to avoid asymmetry stress.
5. Outlook
Hoping the reader can forgive our roadmap story one more time, it needs to be reiterated that there is heavy construction going on in the country we reviewed. A new freeway or a new settlement in one area may suddenly redirect traffic and render Main Street a neglected place. In a foreseeable future, there shall be several alternative protocols available for every simple model of interest and time will have to tell which of them become standard. At some point, using symmetric vesicles for a model study may become as unpopular with reviewers as it happened with DMPC as generic membrane model some decades ago. More sophisticated types of asymmetry as exemplified for cholesterol will become accessible at some point.
From our perspective, it would be useful to agree on a uniform definition of lipid asymmetry in the future. The column regarding degree of asymmetry in Table 1. shows that 14 different definitions of asymmetry are present. To date, there have been a number of attempts to define lipid asymmetry more clearly. We previously introduced an asymmetry parameter a that comprises the amount of asymmetrically distributed lipid, i.e. PS, and ranges from −1 (all-inside localization of PS) via 0 (symmetric distribution) to 1 (outside-only PS) [15]. Guo et al. described the asymmetric degree a of PS molecules in the membrane based on its monolayer concentration in the inner and outer bilayer leaflet [27]. Eicher et al. defined asymmetry, ∑as, as the difference of donor lipid mole fraction in the outer and inner leaflet [66]. Another approach defines Pa% as percentage of lipid exchange in asymmetric vesicles with regard to symmetric and asymmetric vesicles [45,46,47].
The above mentioned, varying and in some cases lacking definitions of lipid asymmetry demonstrate the need for a uniform specification to improve comparability of the large number of methods for preparation of liposomes.
Another aspect of interest and potential for further investigation is the asymmetry stability issue. In individual cases, if specifically highlighted in the article concerned, we have already mentioned asymmetry stability in chapter two (see above). That topic receives varying levels of attention; while in some protocol asymmetry is confirmed for at least a few hours, others provide stability data over several days. Again, it is not trivial to compare data of different methods and systems.
We tried our best to compile a selection of optimal protocols based on what the individual papers are providing. In the future, it will be important to have comparative reports of one laboratory having tested different protocols to serve a given purpose. This will give rise to improved, dedicated roadmaps for a vehicle of interest.
We now slowly start getting rewarded for using asymmetric models by learning about the functions of lipid asymmetry in biology and its potential use for technical applications.
Author Contributions
Conceptualization, M.K. and H.H.; investigation, M.K.; writing—original draft preparation, M.K.; writing—review and editing, M.K. and H.H.; visualization, M.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We acknowledge the wonderfully collaborative, open-minded and friendly spirit of the lipid asymmetry community. In particular, we dedicate this contribution to Georg’s “asymmetry roundtable” held on skype long before we all became online-conference experts. It successfully ignored that we all were supposed to behave as competitors and this way, became a liveable Utopia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Marquardt, D.; Geier, B.; Pabst, G. Asymmetric lipid membranes: Towards more realistic model systems. Membranes 2015, 5, 180–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, R.J.; Hossain, K.R.; Cao, K. Physiological roles of transverse lipid asymmetry of animal membranes. Biochim. Biophys. Acta-Biomembr. 2020, 1862, 183382. [Google Scholar] [CrossRef]
- Dimova, R.; Aranda, S.; Bezlyepkina, N.; Nikolov, V.; Riske, K.A.; Lipowsky, R. A practical guide to giant vesicles. Probing the membrane nanoregime via optical microscopy. J. Phys. Condens. Matter 2006, 18, S1151–S1176. [Google Scholar] [CrossRef] [Green Version]
- Walde, P.; Cosentino, K.; Engel, H.; Stano, P. Giant Vesicles: Preparations and Applications. ChemBioChem 2010, 11, 848–865. [Google Scholar] [CrossRef]
- Kamiya, K.; Takeuchi, S. Giant liposome formation toward the synthesis of well-defined artificial cells. J. Mater. Chem. B 2017, 5, 5911–5923. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, K. Development of artificial cell models using microfluidic technology and synthetic biology. Micromachines 2020, 11, 559. [Google Scholar] [CrossRef]
- Hu, P.C.; Malmstadt, N. Asymmetric Giant Lipid Vesicle Fabrication. Methods Membr. Lipids Second Ed. 2014, 1232, 1–327. [Google Scholar] [CrossRef]
- Huang, Y.; Kim, S.H.; Arriaga, L.R. Emulsion templated vesicles with symmetric or asymmetric membranes. Adv. Colloid Interface Sci. 2017, 247, 413–425. [Google Scholar] [CrossRef]
- Ai, Y.; Xie, R.; Xiong, J.; Liang, Q. Microfluidics for Biosynthesizing: From Droplets and Vesicles to Artificial Cells. Small 2020, 16, e1903940. [Google Scholar] [CrossRef]
- Céspedes, P.F.; Beckers, D.; Dustin, M.L.; Sezgin, E. Model membrane systems to reconstitute immune cell signaling. FEBS J. 2021, 288, 1070–1090. [Google Scholar] [CrossRef] [PubMed]
- Kakuda, S.; Li, B.; London, E. Preparation and Utility of Asymmetric Lipid Vesicles for Studies of Perfringolysin O-Lipid Interactions, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2021; Volume 649, ISBN 9780128238585. [Google Scholar] [CrossRef]
- London, E. Ordered Domain (Raft) Formation in Asymmetric Vesicles and Its Induction upon Loss of Lipid Asymmetry in Artificial and Natural Membranes. Membranes 2022, 12, 870. [Google Scholar] [CrossRef] [PubMed]
- Scott, H.L.; Kennison, K.B.; Enoki, T.A.; Doktorova, M.; Kinnun, J.J.; Heberle, F.A.; Katsaras, J. Model Membrane Systems Used to Study Plasma Membrane Lipid Asymmetry. Symmetry 2021, 13, 1356. [Google Scholar] [CrossRef] [PubMed]
- Takaoka, R.; Kurosaki, H.; Nakao, H.; Ikeda, K.; Nakano, M. Formation of asymmetric vesicles via phospholipase D-mediated transphosphatidylation. Biochim. Biophys. Acta-Biomembr. 2018, 1860, 245–249. [Google Scholar] [CrossRef]
- Drechsler, C.; Markones, M.; Choi, J.Y.; Frieling, N.; Fiedler, S.; Voelker, D.R.; Schubert, R.; Heerklotz, H. Preparation of Asymmetric Liposomes Using a Phosphatidylserine Decarboxylase. Biophys. J. 2018, 115, 1509–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denkins, Y.M.; Schroit, A.J. Phosphatidylserine decarboxylase: Generation of asymmetric vesicles and determination of the transbilayer distribution of fluorescent phosphatidylserine in model membrane systems. BBA-Biomembr. 1986, 862, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Klotz, K.H.; Bartoldus, I.; Stegmann, T. Membrane asymmetry is maintained during influenza-induced fusion. J. Biol. Chem. 1996, 271, 2383–2386. [Google Scholar] [CrossRef] [Green Version]
- Madden, T.D.; Harrigan, P.R.; Tai, L.C.L.; Bally, M.B.; Mayer, L.D.; Redelmeier, T.E.; Loughrey, H.C.; Tilcock, C.P.S.; Reinish, L.W.; Cullis, P.R. The accumulation of drugs within large unilamellar vesicles exhibiting a proton gradient: A survey. Chem. Phys. Lipids 1990, 53, 37–46. [Google Scholar] [CrossRef]
- Eastman, S.J.; Hope, M.J.; Wong, K.F.; Cullis, P.R. Influence of Phospholipid Asymmetry on Fusion Between Large Unilamellar Esicles. Biochemistry 1992, 31, 4262–4268. [Google Scholar] [CrossRef]
- Bailey, A.L.; Cullis, P.R. Modulation of Membrane Fusion by Asymmetric Transbilayer Distributions of Amino Lipids. Biochemistry 1994, 33, 12573–12580. [Google Scholar] [CrossRef] [PubMed]
- Hope, M.J.; Redelmeier, T.E.; Wong, K.F.; Rodrigueza, W.; Cullis, P.R. Phospholipid Asymmetry in Large Unilamellar Vesicles Induced by Transmembrane pH Gradients. Biochemistry 1989, 28, 4181–4187. [Google Scholar] [CrossRef] [PubMed]
- Redelmeier, T.E.; Hope, M.J.; Cullis, P.R. On the Mechanism of Transbilayer Transport of Phosphatidylglycerol in Response to Transmembrane pH Gradients. Biochemistry 1990, 29, 3046–3053. [Google Scholar] [CrossRef]
- Tilcock, C.; Eastman, S.; Fisher, D. Induction of lipid asymmetry and exchange in model membrane systems. J. Dispers. Sci. Technol. 1991, 12, 129–144. [Google Scholar] [CrossRef]
- Farge, E.; Devaux, P.F. Shape changes of giant liposomes induced by an asymmetric transmembrane distribution of phospholipids. Biophys. J. 1992, 61, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Tachibana, K. NMR observation on transbilayer distribution of N-[13C]methylated chlorpromazine in asymmetric lipid bilayer of unilamellar vesicles. Chem. Lett. 2000, 29, 302–303. [Google Scholar] [CrossRef]
- Sun, H.Y.; Deng, G.; Jiang, Y.W.; Zhou, Y.; Xu, J.; Wu, F.G.; Yu, Z.W. Controllable engineering of asymmetric phosphatidylserine-containing lipid vesicles using calcium cations. Chem. Commun. 2017, 53, 12762–12765. [Google Scholar] [CrossRef]
- Guo, H.Y.; Sun, H.Y.; Deng, G.; Xu, J.; Wu, F.G.; Yu, Z.W. Fabrication of asymmetric phosphatidylserine-containing lipid vesicles: A study on the effects of size, temperature, and lipid composition. Langmuir 2020, 36, 12684–12691. [Google Scholar] [CrossRef] [PubMed]
- Pautot, S.; Frisken, B.J.; Weitz, D.A. Engineering Asymmetric Vesicles. Proc. Natl. Acad. Sci. USA 2003, 100, 10718–10721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pautot, S.; Frisken, B.J.; Weitz, D.A. Production of unilamellar vesicles using an inverted emulsion. Langmuir 2003, 19, 2870–2879. [Google Scholar] [CrossRef]
- Hu, P.C.; Li, S.; Malmstadt, N. Microfluidic fabrication of asymmetric giant lipid vesicles. ACS Appl. Mater. Interfaces 2011, 3, 1434–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matosevic, S.; Paegel, B.M. Layer-by-layer cell membrane assembly. Nat. Chem. 2013, 5, 958–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Schertzer, J.W.; Chiarot, P.R. Continuous microfluidic fabrication of synthetic asymmetric vesicles. Lab Chip 2015, 15, 3591–3599. [Google Scholar] [CrossRef] [PubMed]
- Arriaga, L.R.; Huang, Y.; Kim, S.H.; Aragones, J.L.; Ziblat, R.; Koehler, S.A.; Weitz, D.A. Single-step assembly of asymmetric vesicles. Lab Chip 2019, 19, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Romanov, V.; McCullough, J.; Gale, B.K.; Frost, A. A Tunable Microfluidic Device Enables Cargo Encapsulation by Cell- or Organelle-Sized Lipid Vesicles Comprising Asymmetric Lipid Bilayers. Adv. Biosyst. 2019, 3, 1900010. [Google Scholar] [CrossRef]
- Maktabi, S.; Malmstadt, N.; Schertzer, J.W.; Chiarot, P.R. An integrated microfluidic platform to fabricate single-micrometer asymmetric giant unilamellar vesicles (GUVs) using dielectrophoretic separation of microemulsions. Biomicrofluidics 2021, 15, 024112. [Google Scholar] [CrossRef] [PubMed]
- Richmond, D.L.; Schmid, E.M.; Martens, S.; Stachowiak, J.C.; Liska, N.; Fletcher, D.A. Forming giant vesicles with controlled membrane composition, asymmetry, and contents. Proc. Natl. Acad. Sci. USA 2011, 108, 9431–9436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamiya, K.; Kawano, R.; Osaki, T.; Akiyoshi, K.; Takeuchi, S. Cell-sized asymmetric lipid vesicles facilitate the investigation of asymmetric membranes. Nat. Chem. 2016, 8, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Gotanda, M.; Kamiya, K.; Osaki, T.; Fujii, S.; Misawa, N.; Miki, N.; Takeuchi, S. Sequential generation of asymmetric lipid vesicles using a pulsed-jetting method in rotational wells. Sensors Actuators B Chem. 2018, 261, 392–397. [Google Scholar] [CrossRef]
- Kamiya, K.; Osaki, T.; Takeuchi, S. Formation of nano-sized lipid vesicles with asymmetric lipid components using a pulsed-jet flow method. Sensors Actuators B Chem. 2021, 327, 128917. [Google Scholar] [CrossRef]
- Kamiya, K.; Arisaka, C.; Suzuki, M. Investigation of fusion between nanosized lipid vesicles and a lipid monolayer toward formation of giant lipid vesicles with various kinds of biomolecules. Micromachines 2021, 12, 133. [Google Scholar] [CrossRef] [PubMed]
- Hamada, T.; Miura, Y.; Komatsu, Y.; Kishimoto, Y.; Vestergaard, M.; Takagi, M. Construction of asymmetric cell-sized lipid vesicles from lipid-coated water-in-oil microdroplets. J. Phys. Chem. B 2008, 112, 14678–14681. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Doak, W.J.; Schertzer, J.W.; Chiarot, P.R. Membrane mechanical properties of synthetic asymmetric phospholipid vesicles. Soft Matter 2016, 12, 7521–7528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamdad, K.; Law, R.V.; Seddon, J.M.; Brooks, N.J.; Ces, O. Studying the effects of asymmetry on the bending rigidity of lipid membranes formed by microfluidics. Chem. Commun. 2016, 52, 5277–5280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elani, Y.; Purushothaman, S.; Booth, P.J.; Seddon, J.M.; Brooks, N.J.; Law, R.V.; Ces, O. Measurements of the effect of membrane asymmetry on the mechanical properties of lipid bilayers. Chem. Commun. 2015, 51, 6976–6979. [Google Scholar] [CrossRef] [Green Version]
- Enoki, T.A.; Feigenson, G.W. Asymmetric Bilayers by Hemifusion: Method and Leaflet Behaviors. Biophys. J. 2019, 117, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- Enoki, T.A.; Wu, J.; Heberle, F.A.; Feigenson, G.W. Investigation of the domain line tension in asymmetric vesicles prepared via hemifusion. Biochim. Biophys. Acta-Biomembr. 2021, 1863, 183586. [Google Scholar] [CrossRef]
- Enoki, T.A.; Feigenson, G.W. Improving our picture of the plasma membrane: Rafts induce ordered domains in a simplified model cytoplasmic leaflet. Biochim. Biophys. Acta-Biomembr. 2022, 1864, 183995. [Google Scholar] [CrossRef] [PubMed]
- Markones, M.; Drechsler, C.; Kaiser, M.; Kalie, L.; Heerklotz, H.; Fiedler, S. Engineering Asymmetric Lipid Vesicles: Accurate and Convenient Control of the Outer Leaflet Lipid Composition. Langmuir 2018, 34, 1999–2005. [Google Scholar] [CrossRef] [PubMed]
- Markones, M.; Fippel, A.; Kaiser, M.; Drechsler, C.; Hunte, C.; Heerklotz, H. Stairway to Asymmetry: Five Steps to Lipid-Asymmetric Proteoliposomes. Biophys. J. 2020, 118, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Petroff, J.T.; Dietzen, N.M.; Santiago-McRae, E.; Deng, B.; Washington, M.S.; Chen, L.J.; Trent Moreland, K.; Deng, Z.; Rau, M.; Fitzpatrick, J.A.J.; et al. Open-channel structure of a pentameric ligand-gated ion channel reveals a mechanism of leaflet-specific phospholipid modulation. Nat. Commun. 2022, 13, 7017. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, A.A.P.; Schleiff, E.; Röhring, C.; Loidl-Stahlhofen, A.; Vergères, G. Interactions of myristoylated alanine-rich C kinase substrate (MARCKS)- related protein with a novel solid-supported lipid membrane system (TRANSIL). Anal. Biochem. 1999, 268, 343–353. [Google Scholar] [CrossRef]
- Reinl, H.M.; Bayerl, T.M. Lipid Transfer between Small Unilamellar Vesicles and Single Bilayers on a Solid Support: Self-Assembly of Supported Bilayers with Asymmetric Lipid Distribution. Biochemistry 1994, 33, 14091–14099. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kelley, E.G.; Batchu, K.C.; Porcar, L.; Perez-Salas, U. Creating Asymmetric Phospholipid Vesicles via Exchange with Lipid-Coated Silica Nanoparticles. Langmuir 2020, 36, 8865–8873. [Google Scholar] [CrossRef]
- Yanagisawa, M.; Iwamoto, M.; Kato, A.; Yoshikawa, K.; Oiki, S. Oriented reconstitution of a membrane protein in a giant unilamellar vesicle: Experimental verification with the potassium channel KcsA. J. Am. Chem. Soc. 2011, 133, 11774–11779. [Google Scholar] [CrossRef] [PubMed]
- Perillo, V.L.; Peñalva, D.A.; Vitale, A.J.; Barrantes, F.J.; Antollini, S.S. Transbilayer asymmetry and sphingomyelin composition modulate the preferential membrane partitioning of the nicotinic acetylcholine receptor in Lo domains. Arch. Biochem. Biophys. 2016, 591, 76–86. [Google Scholar] [CrossRef] [PubMed]
- De Kruijff, B.; Wirtz, K.W.A. Induction of a relatively fast transbilayer movement of phosphatidylcholine in vesicles. A 13C NMR study. BBA-Biomembr. 1977, 468, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Everett, J.; Zlotnick, A.; Tennyson, J.; Holloway, P.W. Fluorescence quenching of cytochrome b5 in vesicles with an asymmetric transbilayer distribution of brominated phosphatidylcholine. J. Biol. Chem. 1986, 261, 6725–6729. [Google Scholar] [CrossRef] [PubMed]
- Sandra, A.; Pagano, R.E. Liposome-cell interactions. Studies of lipid transfer using isotopically asymmetric vesicles. J. Biol. Chem. 1979, 254, 2244–2249. [Google Scholar] [CrossRef]
- Holzer, M.; Momm, J.; Schubert, R. Lipid transfer mediated by a recombinant pro-sterol carrier protein 2 for the accurate preparation of asymmetrical membrane vesicles requires a narrow vesicle size distribution: A free-flow electrophoresis study. Langmuir 2010, 26, 4142–4151. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, D.; Heberle, F.A.; Miti, T.; Eicher, B.; London, E.; Katsaras, J.; Pabst, G. 1H NMR Shows Slow Phospholipid Flip-Flop in Gel and Fluid Bilayers. Langmuir 2017, 33, 3731–3741. [Google Scholar] [CrossRef]
- Cheng, H.T.; Megha; London, E. Preparation and properties of asymmetric vesicles that mimic cell membranes. Effect upon lipid raft formation and transmembrane helix orientation. J. Biol. Chem. 2009, 284, 6079–6092. [Google Scholar] [CrossRef] [Green Version]
- Heberle, F.A.; Marquardt, D.; Doktorova, M.; Geier, B.; Standaert, R.F.; Heftberger, P.; Kollmitzer, B.; Nickels, J.D.; Dick, R.A.; Feigenson, G.W.; et al. Subnanometer Structure of an Asymmetric Model Membrane: Interleaflet Coupling Influences Domain Properties. Langmuir 2016, 32, 5195–5200. [Google Scholar] [CrossRef] [PubMed]
- Doktorova, M.; Heberle, F.A.; Eicher, B.; Standaert, R.F.; Katsaras, J.; London, E.; Pabst, G.; Marquardt, D. Preparation of asymmetric phospholipid vesicles for use as cell membrane models. Nat. Protoc. 2018, 13, 2086–2101. [Google Scholar] [CrossRef]
- Sugiura, T.; Ikeda, K.; Nakano, M. Kinetic Analysis of the Methyl-β-cyclodextrin-Mediated Intervesicular Transfer of Pyrene-Labeled Phospholipids. Langmuir 2016, 32, 13697–13705. [Google Scholar] [CrossRef] [PubMed]
- Eicher, B.; Heberle, F.A.; Marquardt, D.; Rechberger, G.N.; Katsaras, J.; Pabst, G. Joint small-angle X-ray and neutron scattering data analysis of asymmetric lipid vesicles. J. Appl. Crystallogr. 2017, 50, 419–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eicher, B.; Marquardt, D.; Heberle, F.A.; Letofsky-Papst, I.; Rechberger, G.N.; Appavou, M.S.; Katsaras, J.; Pabst, G. Intrinsic Curvature-Mediated Transbilayer Coupling in Asymmetric Lipid Vesicles. Biophys. J. 2018, 114, 146–157. [Google Scholar] [CrossRef] [Green Version]
- Courtney, K.C.; Pezeshkian, W.; Raghupathy, R.; Zhang, C.; Darbyson, A.; Ipsen, J.H.; Ford, D.A.; Khandelia, H.; Presley, J.F.; Zha, X. C24 Sphingolipids Govern the Transbilayer Asymmetry of Cholesterol and Lateral Organization of Model and Live-Cell Plasma Membranes. Cell Rep. 2018, 24, 1037–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doktorova, M.; Heberle, F.A.; Marquardt, D.; Rusinova, R.; Sanford, R.L.; Peyear, T.A.; Katsaras, J.; Feigenson, G.W.; Weinstein, H.; Andersen, O.S. Gramicidin Increases Lipid Flip-Flop in Symmetric and Asymmetric Lipid Vesicles. Biophys. J. 2019, 116, 860–873. [Google Scholar] [CrossRef] [Green Version]
- Rickeard, B.W.; Nguyen, M.H.L.; Dipasquale, M.; Yip, C.G.; Baker, H.; Heberle, F.A.; Zuo, X.; Kelley, E.G.; Nagao, M.; Marquardt, D. Transverse lipid organization dictates bending fluctuations in model plasma membranes. Nanoscale 2020, 12, 1438–1447. [Google Scholar] [CrossRef]
- Scott, H.L.; Heberle, F.A.; Katsaras, J.; Barrera, F.N. Phosphatidylserine Asymmetry Promotes the Membrane Insertion of a Transmembrane Helix. Biophys. J. 2019, 116, 1495–1506. [Google Scholar] [CrossRef]
- Marx, L.; Frewein, M.P.K.; Semeraro, E.F.; Rechberger, G.N.; Lohner, K.; Porcar, L.; Pabst, G. Antimicrobial peptide activity in asymmetric bacterial membrane mimics. Faraday Discuss. 2021, 232, 435–447. [Google Scholar] [CrossRef]
- Frewein, M.P.K.; Piller, P.; Semeraro, E.F.; Batchu, K.C.; Heberle, F.A.; Scott, H.L.; Gerelli, Y.; Porcar, L.; Pabst, G. Interdigitation-Induced Order and Disorder in Asymmetric Membranes. J. Membr. Biol. 2022, 255, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Chiantia, S.; Schwille, P.; Klymchenko, A.S.; London, E. Asymmetric GUVs prepared by MβCD-mediated lipid exchange: An FCS study. Biophys. J. 2011, 100, L1–L3. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.T.; London, E. Preparation and properties of asymmetric large unilamellar vesicles: Interleaflet coupling in asymmetric vesicles is dependent on temperature but not curvature. Biophys. J. 2011, 100, 2671–2678. [Google Scholar] [CrossRef] [Green Version]
- Chiantia, S.; London, E. Acyl Chain length and saturation modulate interleaflet coupling in asymmetric bilayers: Effects on dynamics and structural order. Biophys. J. 2012, 103, 2311–2319. [Google Scholar] [CrossRef] [Green Version]
- Son, M.; London, E. The dependence of lipid asymmetry upon phosphatidylcholine acyl chain structure. J. Lipid Res. 2013, 54, 223–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petazzi, R.A.; Gramatica, A.; Herrmann, A.; Chiantia, S. Time-controlled phagocytosis of asymmetric liposomes: Application to phosphatidylserine immunoliposomes binding HIV-1 virus-like particles. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 1985–1992. [Google Scholar] [CrossRef]
- Lin, Q.; London, E. Preparation of artificial plasma membrane mimicking vesicles with lipid asymmetry. PLoS ONE 2014, 9, e87903. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; London, E. The influence of natural lipid asymmetry upon the conformation of a membrane-inserted protein (perfringolysin O). J. Biol. Chem. 2014, 289, 5467–5478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Q.; London, E. Ordered raft domains induced by outer leaflet sphingomyelin in cholesterol-rich asymmetric vesicles. Biophys. J. 2015, 108, 2212–2222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; London, E. Lipid Structure and Composition Control Consequences of Interleaflet Coupling in Asymmetric Vesicles. Biophys. J. 2018, 115, 664–678. [Google Scholar] [CrossRef] [PubMed]
- St. Clair, J.W.; London, E. Effect of sterol structure on ordered membrane domain (raft) stability in symmetric and asymmetric vesicles. Biochim. Biophys. Acta-Biomembr. 2019, 1861, 1112–1122. [Google Scholar] [CrossRef]
- Li, B.; London, E. Preparation and drug entrapment properties of asymmetric liposomes containing cationic and anionic lipids. Langmuir 2020, 36, 12521–12531. [Google Scholar] [CrossRef]
- Li, M.H.; Raleigh, D.P.; London, E. Preparation of Asymmetric Vesicles with Trapped CsCl Avoids Osmotic Imbalance, Non-Physiological External Solutions, and Minimizes Leakage. Langmuir 2021, 37, 11611–11617. [Google Scholar] [CrossRef]
- Epand, R.F.; Martinou, J.C.; Montessuit, S.; Epand, R.M. Transbilayer Lipid Diffusion Promoted by Bax: Implications for Apoptosis. Biochemistry 2003, 42, 14576–14582. [Google Scholar] [CrossRef]
- Konetski, D.; Zhang, D.; Schwartz, D.K.; Bowman, C.N. Photoinduced Pinocytosis for Artificial Cell and Protocell Systems. Chem. Mater. 2018, 30, 8757–8763. [Google Scholar] [CrossRef]
- Howe, A.; Sofou, S. Daptomycin-Induced Lipid Phases on Model Lipid Bilayers: Effect of Lipid Type and of Lipid Leaflet Order on Membrane Permeability. J. Phys. Chem. B 2021, 125, 5775–5785. [Google Scholar] [CrossRef] [PubMed]
- Miwa, A.; Kamiya, K. Control of Enzyme Reaction Initiation inside Giant Unilamellar Vesicles by the Cell-Penetrating Peptide-Mediated Translocation of Cargo Proteins. ACS Synth. Biol. 2022, 11, 3836–3846. [Google Scholar] [CrossRef] [PubMed]
- Bozelli, J.C.; Hou, Y.H.; Schreier, S.; Epand, R.M. Lipid asymmetry of a model mitochondrial outer membrane affects Bax-dependent permeabilization. Biochim. Biophys. Acta-Biomembr. 2020, 1862, 183241. [Google Scholar] [CrossRef]
- Varma, M.; Deserno, M. Distribution of cholesterol in asymmetric membranes driven by composition and differential stress. Biophys. J. 2022, 121, 4001–4018. [Google Scholar] [CrossRef] [PubMed]
- Doktorova, M.; Levental, I. Cholesterol’s balancing act: Defying the status quo. Biophys. J. 2022, 121, 3771–3773. [Google Scholar] [CrossRef] [PubMed]
- Tsamaloukas, A.; Szadkowska, H.; Slotte, P.J.; Heerklotz, H. Interactions of cholesterol with lipid membranes and cyclodextrin characterized by calorimetry. Biophys. J. 2005, 89, 1109–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, T.G.; Tan, A.; Ganz, P.; Seelig, J. Calorimetric Measurement of Phospholipid Interaction with Methyl-β-Cyclodextrin. Biochemistry 2004, 43, 2251–2261. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Road map to asymmetry, illustrating the criteria and considerations to select one or more suitable protocols described in the literature for preparing lipid-asymmetric vesicles that suit a given purpose. Eleven protocols are distinguished (yellow boxes) that can be grouped into four strategies (A–D). So far, exchange protocols represent the most abundant and most versatile strategy—illustrated here as the destination of Main Street. Gray boxes (see text for more detail) indicate criteria for exits to alternative protocols that may be favorable for a given purpose. Examples for each protocol (A1–D5) are listed in Table 1.
Figure 1.
Road map to asymmetry, illustrating the criteria and considerations to select one or more suitable protocols described in the literature for preparing lipid-asymmetric vesicles that suit a given purpose. Eleven protocols are distinguished (yellow boxes) that can be grouped into four strategies (A–D). So far, exchange protocols represent the most abundant and most versatile strategy—illustrated here as the destination of Main Street. Gray boxes (see text for more detail) indicate criteria for exits to alternative protocols that may be favorable for a given purpose. Examples for each protocol (A1–D5) are listed in Table 1.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Krompers, M.; Heerklotz, H. A Guide to Your Desired Lipid-Asymmetric Vesicles. Membranes 2023, 13, 267. https://doi.org/10.3390/membranes13030267
AMA Style
Krompers M, Heerklotz H. A Guide to Your Desired Lipid-Asymmetric Vesicles. Membranes. 2023; 13(3):267. https://doi.org/10.3390/membranes13030267
Chicago/Turabian StyleKrompers, Mona, and Heiko Heerklotz. 2023. "A Guide to Your Desired Lipid-Asymmetric Vesicles" Membranes 13, no. 3: 267. https://doi.org/10.3390/membranes13030267
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.