
Formation of β-Strand Oligomers of Antimicrobial Peptide Magainin 2 Contributes to Disruption of Phospholipid Membrane


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Liposomes and Sample Preparation
2.3. Synchrotron-Radiation Circular Dichroism
2.4. Synchrotron-Radiation Linear Dichroism
2.5. Fluorescence Anisotropy
2.6. Calcein Leakage Assay
2.7. Adsorption Model Fitting Procedure
3. Results
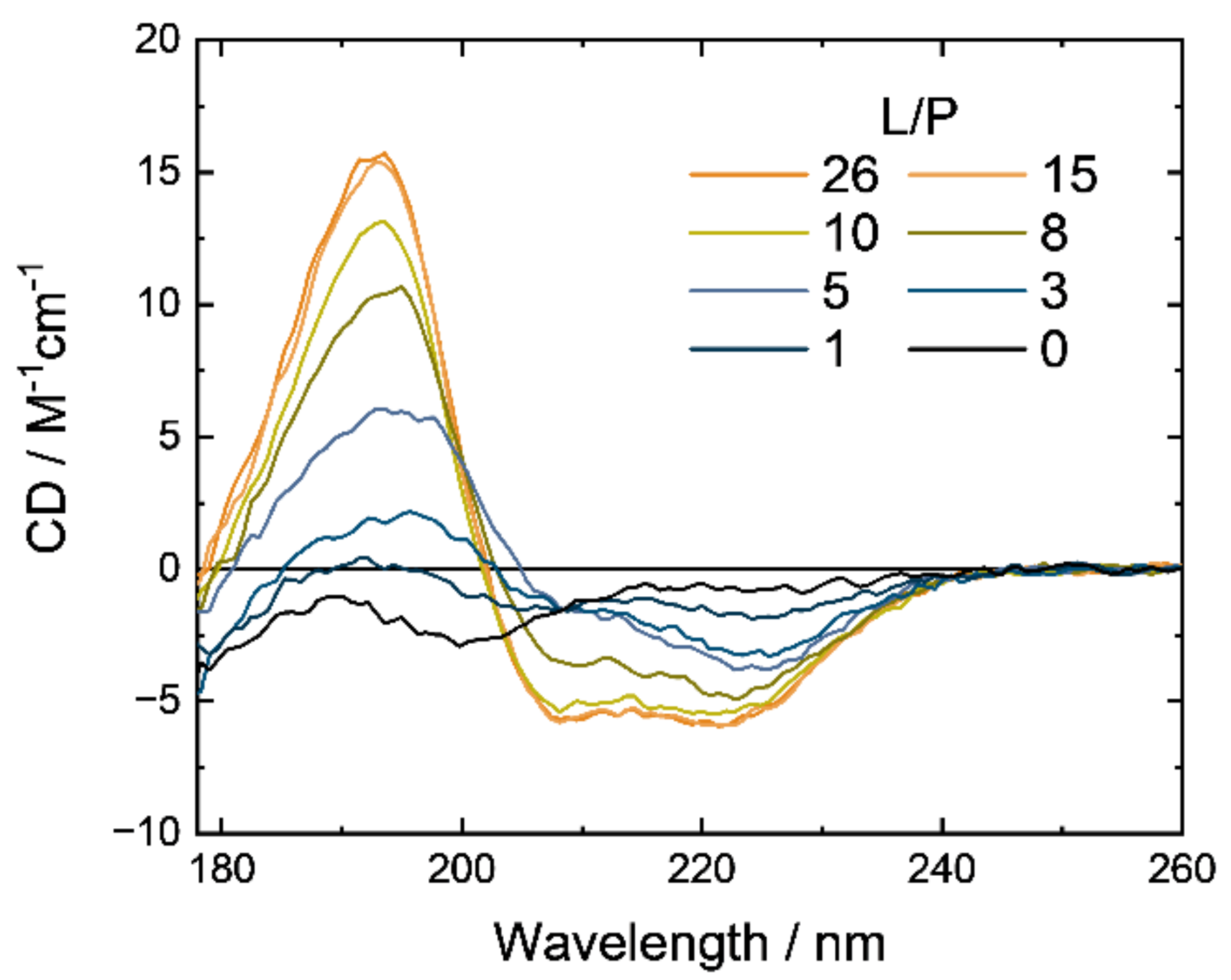
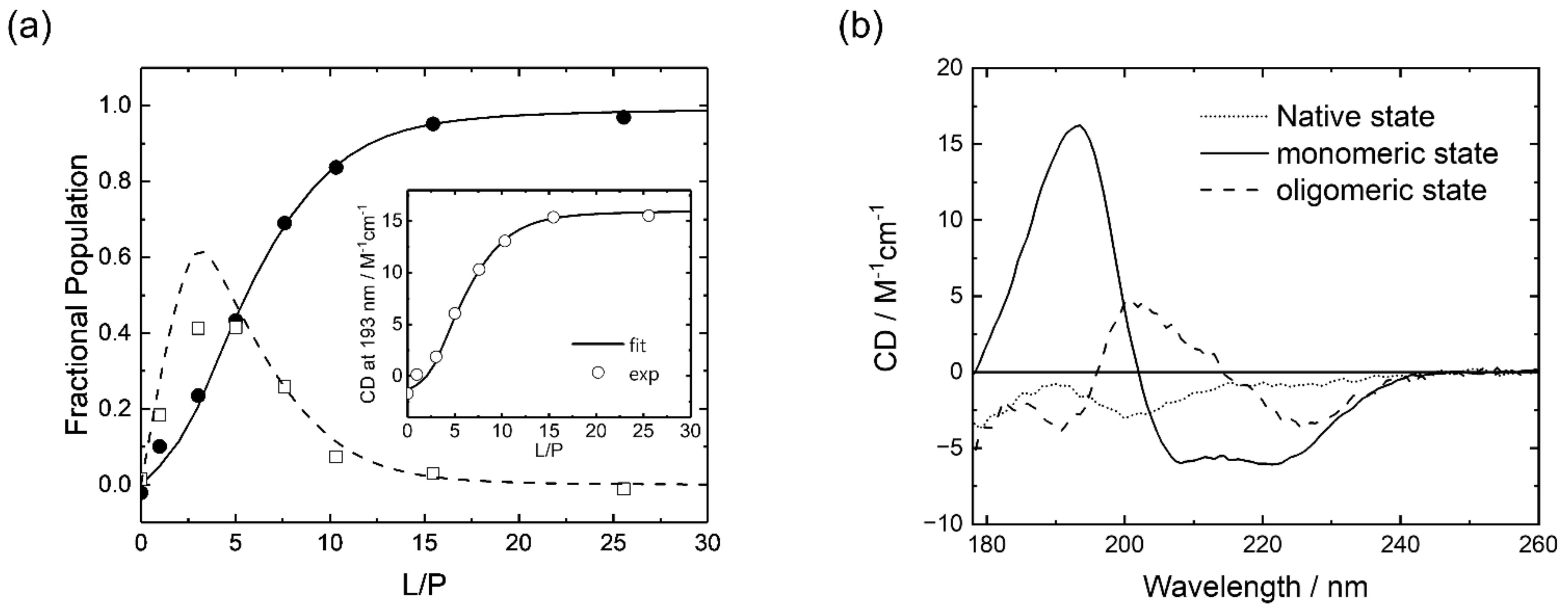
3.1. Synchrotron-Radiation Circular Dichroism
3.2. Synchrotron-Radiation Linear Dichroism
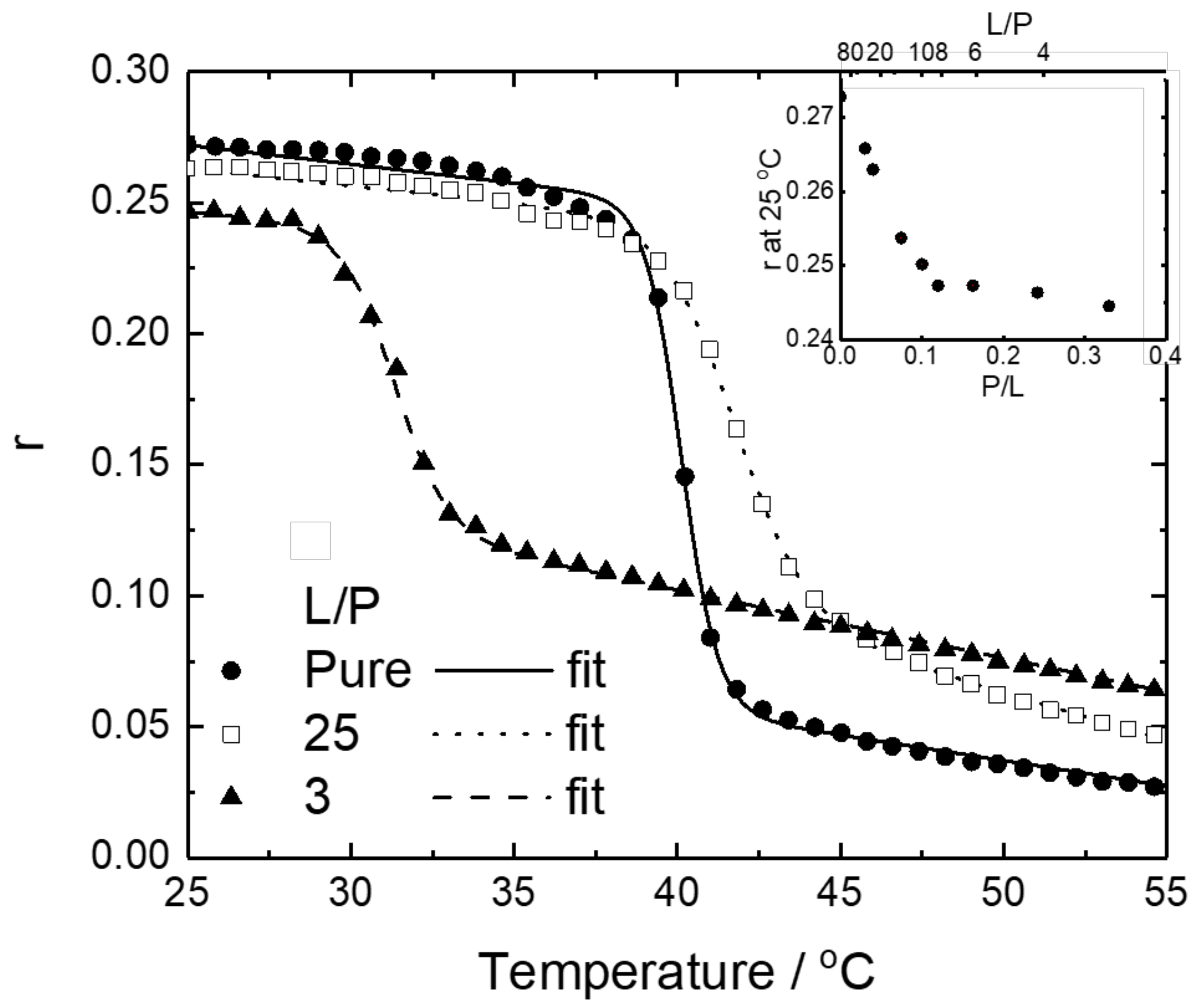
3.3. Fluorescence Anisotropy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hughes, J.M. Preserving the Lifesaving Power of Antimicrobial Agents. JAMA 2011, 305, 1027–1028. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Antimicrobial Peptides of Multicellularorganisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Sahl, H.G. Antimicrobial and Host-Defense Peptides as New Anti-Infective Therapeutic Strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef]
- Melo, M.N.; Ferre, R.; Castanho, M.A.R.B. Antimicrobial Peptides: Linking Partition, Activity and High Membrane-Bound Concentrations. Nat. Rev. Microbiol. 2009, 7, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K. Control of Cell Selectivity of Antimicrobial Peptides. Biochim. Biophys. Acta 2009, 1788, 1687–1692. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Sun, L.C.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q.Y. The Antimicrobial Peptides and Their Potential Clinical Applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar] [PubMed]
- Matsuzaki, K. (Ed.) Antimicrobial Peptides Basics for Clinical Application; Springer: Singapore, 2019; ISBN 978-981-13-3587-7. [Google Scholar]
- Benfield, A.H.; Henriques, S.T. Mode-of-Action of Antimicrobial Peptides: Membrane Disruption vs. Intracellular Mechanisms. Front. Med. Technol. 2020, 2, 610997. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Magainins, a Class of Antimicrobial Peptides from Xenopus Skin: Isolation, Characterization of Two Active Forms, and Partial cDNA Sequence of a Precursor. Proc. Natl. Acad. Sci. USA 1987, 84, 5449–5453. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, K.; Sugishita, K.I.; Ishibe, N.; Ueha, M.; Nakata, S.; Miyajima, K.; Epand, R.M. Relationship of Membrane Curvature to the Formation of Pores by Magainin 2. Biochemistry 1998, 37, 11856–11863. [Google Scholar] [CrossRef]
- Bechinger, B.; Zasloff, M.; Opella, S.J. Structure and Orientation of the Antibiotic Peptide Magainin in Membranes by Solid-State Nuclear Magnetic Resonance Spectroscopy. Protein Sci. 1993, 2, 2077–2084. [Google Scholar] [CrossRef] [Green Version]
- Tamba, Y.; Yamazaki, M. Single Giant Unilamellar Vesicle Method Reveals Effect of Antimicrobial Peptide Magainin 2 on Membrane Permeability. Biochemistry 2005, 44, 15823–15833. [Google Scholar] [CrossRef]
- Wenk, M.R.; Seelig, J. Magainin 2 Amide Interaction with Lipid Membranes: Calorimetric Detection of Peptide Binding and Pore Formation. Biochemistry 1998, 37, 3909–3916. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K.; Murase, O.; Tokuda, H.; Funakoshi, S.; Fujii, N.; Miyajima, K. Orientational and Aggregational States of Magainin 2 in Phospholipid Bilayers. Biochemistry 1994, 33, 3342–3349. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Murase, O.; Miyajima, K. Kinetics of Pore Formation by an Antimicrobial Peptide, Magainin 2, in Phospholipid Bilayers. Biochemistry 1995, 34, 12553–12559. [Google Scholar] [CrossRef]
- Schümann, M.; Dathe, M.; Wieprecht, T.; Beyermann, M.; Bienert, M. The Tendency of Magainin to Associate upon Binding to Phospholipid Bilayers. Biochemistry 1997, 36, 4345–4351. [Google Scholar] [CrossRef] [PubMed]
- Gregory, S.M.; Pokorny, A.; Almeida, P.F.F. Magainin 2 Revisited: A Test of the Quantitative Model for the All-or-None Permeabilization of Phospholipid Vesicles. Biophys. J. 2009, 96, 116–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Travkova, O.G.; Moehwald, H.; Brezesinski, G. The Interaction of Antimicrobial Peptides with Membranes. Adv. Colloid Interface Sci. 2017, 247, 521–532. [Google Scholar] [CrossRef]
- Wimley, W.C. Describing the Mechanism of Antimicrobial Peptide Action with the Interfacial Activity Model. ACS Chem. Biol. 2010, 5, 905–917. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.; Mantsch, H.H.; Spencer, J.H. Conformation of Magainin-2 and Related Peptides in Aqueous Solution and Membrane Environments Probed by Fourier Transform Infrared Spectroscopy. Biochemistry 1992, 31, 7289–7293. [Google Scholar] [CrossRef]
- Hirsh, D.J.; Hammer, J.; Maloy, W.L.; Blazyk, J.; Schaefer, J. Secondary Structure and Location of a Magainin Analogue in Synthetic Phospholipid Bilayers. Biochemistry 1996, 35, 12733–12741. [Google Scholar] [CrossRef]
- Williams, R.W.; Starman, R.; Taylor, K.M.P.; Gable, K.; Beeler, T.; Zasloff, M.; Covell, D. Raman Spectroscopy of Synthetic Antimicrobial Frog Peptides Magainin 2a and PGLa. Biochemistry 1990, 29, 4490–4496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The Expanding Scope of Antimicrobial Peptide Structures and Their Modes of Action. Trends Biotechnol. 2011, 29, 464–472. [Google Scholar] [CrossRef]
- Matsuo, K.; Maki, Y.; Namatame, H.; Taniguchi, M.; Gekko, K. Conformation of Membrane-Bound Proteins Revealed by Vacuum-Ultraviolet Circular-Dichroism and Linear-Dichroism Spectroscopy. Proteins 2016, 84, 349–359. [Google Scholar] [CrossRef]
- Matsuo, K.; Kumashiro, M.; Gekko, K. Characterization of the Mechanism of Interaction Between α1-Acid Glycoprotein and Lipid Membranes by Vacuum-Ultraviolet Circular-Dichroism Spectroscopy. Chirality 2020, 32, 594–604. [Google Scholar] [CrossRef]
- Kumashiro, M.; Izumi, Y.; Matsuo, K. Conformation of Myelin Basic Protein Bound to Phosphatidylinositol Membrane Characterized by Vacuum-Ultraviolet Circular-Dichroism Spectroscopy and Molecular-Dynamics Simulations. Proteins 2021, 89, 1251–1261. [Google Scholar] [CrossRef] [PubMed]
- Hope, M.J.; Bally, M.B.; Webb, G.; Cullis, P.R. Production of Large Unilamellar Vesicles by a Rapid Extrusion Procedure. Characterization of Size Distribution, Trapped Volume and Ability to Maintain a Membrane Potential. Biochim. Biophys. Acta 1985, 812, 55–65. [Google Scholar] [CrossRef]
- Ojima, N.; Sakai, K.; Matsuo, K.; Matsui, T.; Fukazawa, T.; Namatame, H.; Taniguchi, M.; Gekko, K. Vacuum-Ultraviolet Circular Dichroism Spectrophotometer Using Synchrotron Radiation: Optical System and On-Line Performance. Chem. Lett. 2001, 522–523. [Google Scholar] [CrossRef]
- Matsuo, K.; Sakai, K.; Matsushima, Y.; Fukuyama, T.; Gekko, K. Optical Cell with a Temperature-Control Unit for a Vacuum-Ultraviolet Circular Dichroism Spectrophotometer. Anal. Sci. 2003, 19, 129–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreerama, N.; Woody, R.W. Estimation of Protein Secondary Structure from Circular Dichroism Spectra: Comparison of CONTIN, SELCON, and CDSSTR Methods with an Expanded Reference Set. Anal. Biochem. 2000, 287, 252–260. [Google Scholar] [CrossRef]
- Sreerama, N.; Venyaminov, S.Y.U.; Woody, R.W. Estimation of the Number of α-Helical and β-Strand Segments in Proteins Using Circular Dichroism Spectroscopy. Protein Sci. 1999, 8, 370–380. [Google Scholar] [CrossRef]
- Matsuo, K.; Yonehara, R.; Gekko, K. Secondary-Structure Analysis of Proteins by Vacuum-Ultraviolet Circular Dichroism Spectroscopy. J. Biochem. 2004, 135, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Yonehara, R.; Gekko, K. Improved Estimation of the Secondary Structures of Proteins by Vacuum-Ultraviolet Circular Dichroism Spectroscopy. J. Biochem. 2005, 138, 79–88. [Google Scholar] [CrossRef]
- Rodger, A.; Marrington, R.; Geeves, M.A.; Hicks, M.; De Alwis, L.; Halsall, D.J.; Dafforn, T.R. Looking at Long Molecules in Solution: What Happens When They Are Subjected to Couette Flow? Phys. Chem. Chem. Phys. 2006, 8, 3161–3171. [Google Scholar] [CrossRef]
- Lentz, B.R. Use of Fluorescent Probes to Monitor Molecular Order and Motions Within Liposome Bilayers. Chem. Phys. Lipids 1993, 64, 99–116. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd ed.; Springer: Boston, MA, USA, 2006; ISBN 978-0387312781. [Google Scholar]
- Bartlett, G.R. Phosphorus Assay in Column Chromatography. J. Biol. Chem. 1959, 234, 466–468. [Google Scholar] [CrossRef]
- Chatelier, R.C.; Minton, A.P. Adsorption of Globular Proteins on Locally Planar Surfaces: Models for the Effect of Excluded Surface Area and Aggregation of Adsorbed Protein on Adsorption Equilibria. Biophys. J. 1996, 71, 2367–2374. [Google Scholar] [CrossRef] [Green Version]
- Shashilov, V.A.; Lednev, I.K. Advanced Statistical and Numerical Methods for Spectroscopic Characterization of Protein Structural Evolution. Chem. Rev. 2010, 110, 5692–5713. [Google Scholar] [CrossRef]
- Arouri, A.; Dathe, M.; Blume, A. Peptide Induced Demixing in PG/PE Lipid Mixtures: A Mechanism for the Specificity of Antimicrobial Peptides Towards Bacterial Membranes? Biochim. Biophys. Acta 2009, 1788, 650–659. [Google Scholar] [CrossRef] [Green Version]
- Edwards-Gayle, C.J.C.; Barrett, G.; Roy, S.; Castelletto, V.; Seitsonen, J.; Ruokolainen, J.; Hamley, I.W. Selective Antibacterial Activity and Lipid Membrane Interactions of Arginine-Rich Amphiphilic Peptides. ACS Appl. Bio Mater. 2020, 3, 1165–1175. [Google Scholar] [CrossRef]
- Persson, D.; Thorén, P.E.G.; Herner, M.; Lincoln, P.; Nordén, B. Application of a Novel Analysis to Measure the Binding of the Membrane-Translocating Peptide Penetratin to Negatively Charged Liposomes. Biochemistry 2003, 42, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Manavalan, P.; Johnson, W.C. Protein Secondary Structure from Circular Dichroism Spectra. J. Biosci. 1985, 8, 141–149. [Google Scholar] [CrossRef]
- Gorbenko, G.P.; Ioffe, V.M.; Kinnunen, P.K.J. Binding of Lysozyme to Phospholipid Bilayers: Evidence for Protein Aggregation upon Membrane Association. Biophys. J. 2007, 93, 140–153. [Google Scholar] [CrossRef] [Green Version]
- Zuckermann, M.J.; Heimburg, T. Insertion and Pore Formation Driven by Adsorption of Proteins onto Lipid Bilayer Membrane-Water Interfaces. Biophys. J. 2001, 81, 2458–2472. [Google Scholar] [CrossRef] [Green Version]
- Greenfield, N.; Fasman, G.D. Computed Circular Dichroism Spectra for the Evaluation of Protein Conformation. Biochemistry 1969, 8, 4108–4116. [Google Scholar] [CrossRef]
- Gesell, J.; Zasloff, M.; Opella, S.J. Two-Dimensional 1H NMR Experiments Show That the 23-Residue Magainin Antibiotic Peptide Is an α-Helix in Dodecylphosphocholine Micelles, Sodium Dodecylsulfate Micelles, and Trifluoroethanol/Water Solution. J. Biomol. NMR 1997, 9, 127–135. [Google Scholar] [CrossRef]
- Vermeer, L.S.; Marquette, A.; Schoup, M.; Fenard, D.; Galy, A.; Bechinger, B. Simultaneous Analysis of Secondary Structure and Light Scattering from Circular Dichroism Titrations: Application to Vectofusin-1. Sci. Rep. 2016, 6, 39450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, A.J.; Wallace, B.A. Circular Dichroism Spectroscopy of Membrane Proteins. Chem. Soc. Rev. 2016, 45, 4859–4872. [Google Scholar] [CrossRef] [Green Version]
- Micsonai, A.; Wien, F.; Kernya, L.; Lee, Y.H.; Goto, Y.; Réfrégiers, M.; Kardos, J. Accurate Secondary Structure Prediction and Fold Recognition for Circular Dichroism Spectroscopy. Proc. Natl. Acad. Sci. USA 2015, 112, E3095–E3103. [Google Scholar] [CrossRef] [Green Version]
- Rodger, A.; Nordén, B.; Dafforn, T. Linear Dichroism and Circular Dichroism: A Textbook on Polarized-Light Spectroscopy; Royal Society of Chemistry: Cambridge, UK, 2010; ISBN 978-1847559029. [Google Scholar]
- Rodger, A.; Rajendra, J.; Marrington, R.; Ardhammar, M.; Nordén, B.; Hirst, J.D.; Gilbert, A.T.B.; Dafforn, T.R.; Halsall, D.J.; Woolhead, C.A.; et al. Flow Oriented Linear Dichroism to Probe Protein Orientation in Membrane Environments. Phys. Chem. Chem. Phys. 2002, 4, 4051–4057. [Google Scholar] [CrossRef]
- Lentz, B.R. Membrane “Fluidity” as Detected by Diphenylhexatriene Probes. Chem. Phys. Lipids 1989, 50, 171–190. [Google Scholar] [CrossRef]
- Jang, H.; Ma, B.; Lal, R.; Nussinov, R. Models of Toxic β-Sheet Channels of protegrin-1 Suggest a Common Subunit Organization Motif Shared with Toxic Alzheimer β-Amyloid Ion Channels. Biophys. J. 2008, 95, 4631–4642. [Google Scholar] [CrossRef] [Green Version]
- Ciudad, S.; Puig, E.; Botzanowski, T.; Meigooni, M.; Arango, A.S.; Do, J.; Mayzel, M.; Bayoumi, M.; Chaignepain, S.; Maglia, G.; et al. Aβ(1–42) Tetramer and Octamer Structures Reveal Edge Conductivity Pores as a Mechanism for Membrane Damage. Nat. Commun. 2020, 11, 3014. [Google Scholar] [CrossRef] [PubMed]
- Rzeźnicka, I.I.; Pandey, R.; Schleeger, M.; Bonn, M.; Weidner, T. Formation of Lysozyme Oligomers at Model Cell Membranes Monitored with Sum Frequency Generation Spectroscopy. Langmuir 2014, 30, 7736–7744. [Google Scholar] [CrossRef] [PubMed]
- Papahadjopoulos, D.; Moscarello, M.; Eylar, E.H.; Isac, T. Effects of proteins on the thermotropic phase transitions of phospholipid membranes. Biochim. Biophys. Acta Biomembr. 1975, 401, 317–335. [Google Scholar] [CrossRef]
- Quinn, P.J. The Fluidity of Cell Membranes and Its Regulation. Prog. Biophys. Mol. Biol. 1981, 38, 1–104. [Google Scholar] [CrossRef]
- Veld, G.I.; Driessen, A.J.M.; Konings, W.N. Bacterial Solute Transport Proteins in Their Lipid Environment. FEMS Microbiol. Rev. 1993, 12, 293–314. [Google Scholar] [CrossRef] [PubMed]
- Hazel, J.R.; Williams, E.E. The Role of Alterations in Membrane Lipid Composition in Enabling Physiological Adaptation of Organisms to Their Physical Environment. Prog. Lipid Res. 1990, 29, 167–227. [Google Scholar] [CrossRef]
- Avitabile, C.; D’Andrea, L.D.; Romanelli, A. Circular Dichroism Studies on the Interactions of Antimicrobial Peptides with Bacterial Cells. Sci. Rep. 2014, 4, 4293. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| L/P | Tm [°C] | ΔH [kcal/mol] |
|---|---|---|
| Only liposome | 40.12 ± 0.02 | 351 ± 7 |
| 25 | 41.71 ± 0.02 | 180 ± 4 |
| 3 | 31.31 ± 0.03 | 231 ± 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumashiro, M.; Tsuji, R.; Suenaga, S.; Matsuo, K. Formation of β-Strand Oligomers of Antimicrobial Peptide Magainin 2 Contributes to Disruption of Phospholipid Membrane. Membranes 2022, 12, 131. https://doi.org/10.3390/membranes12020131
Kumashiro M, Tsuji R, Suenaga S, Matsuo K. Formation of β-Strand Oligomers of Antimicrobial Peptide Magainin 2 Contributes to Disruption of Phospholipid Membrane. Membranes. 2022; 12(2):131. https://doi.org/10.3390/membranes12020131
Chicago/Turabian StyleKumashiro, Munehiro, Ryoga Tsuji, Shoma Suenaga, and Koichi Matsuo. 2022. "Formation of β-Strand Oligomers of Antimicrobial Peptide Magainin 2 Contributes to Disruption of Phospholipid Membrane" Membranes 12, no. 2: 131. https://doi.org/10.3390/membranes12020131