
Staphylococcus aureus Protection-Related Type 3 Cell-Mediated Immune Response Elicited by Recombinant Proteins and GM-CSF DNA Vaccine

 , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Production of S. aureus Recombinant Proteins
2.3. Production of GM-CSF DNA Vaccine
2.4. Liposome Preparation and Entrapment of Plasmid DNA
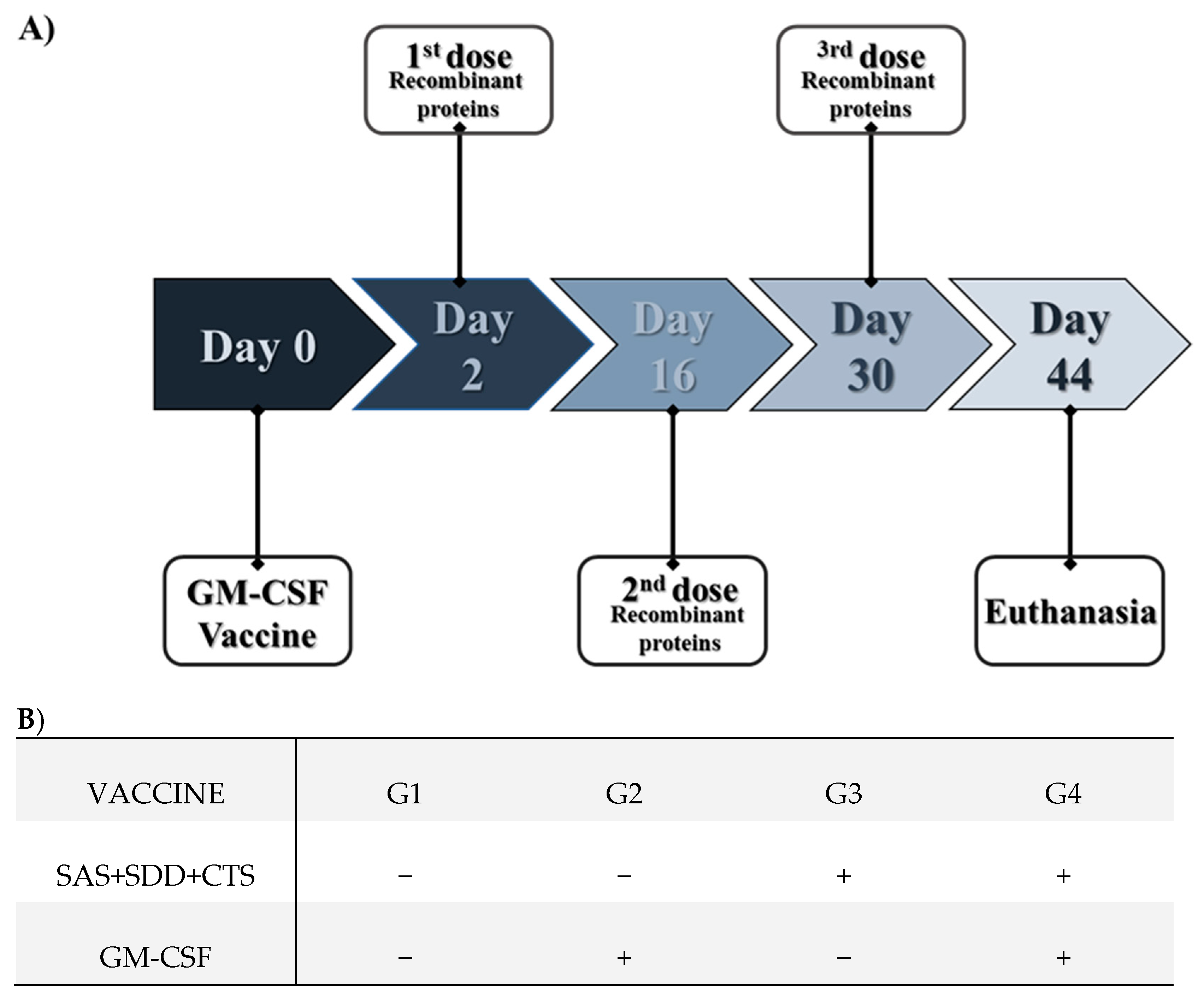
2.5. Animals and Immunization
2.6. Obtaining the Spleen and Lymph Nodes Cells
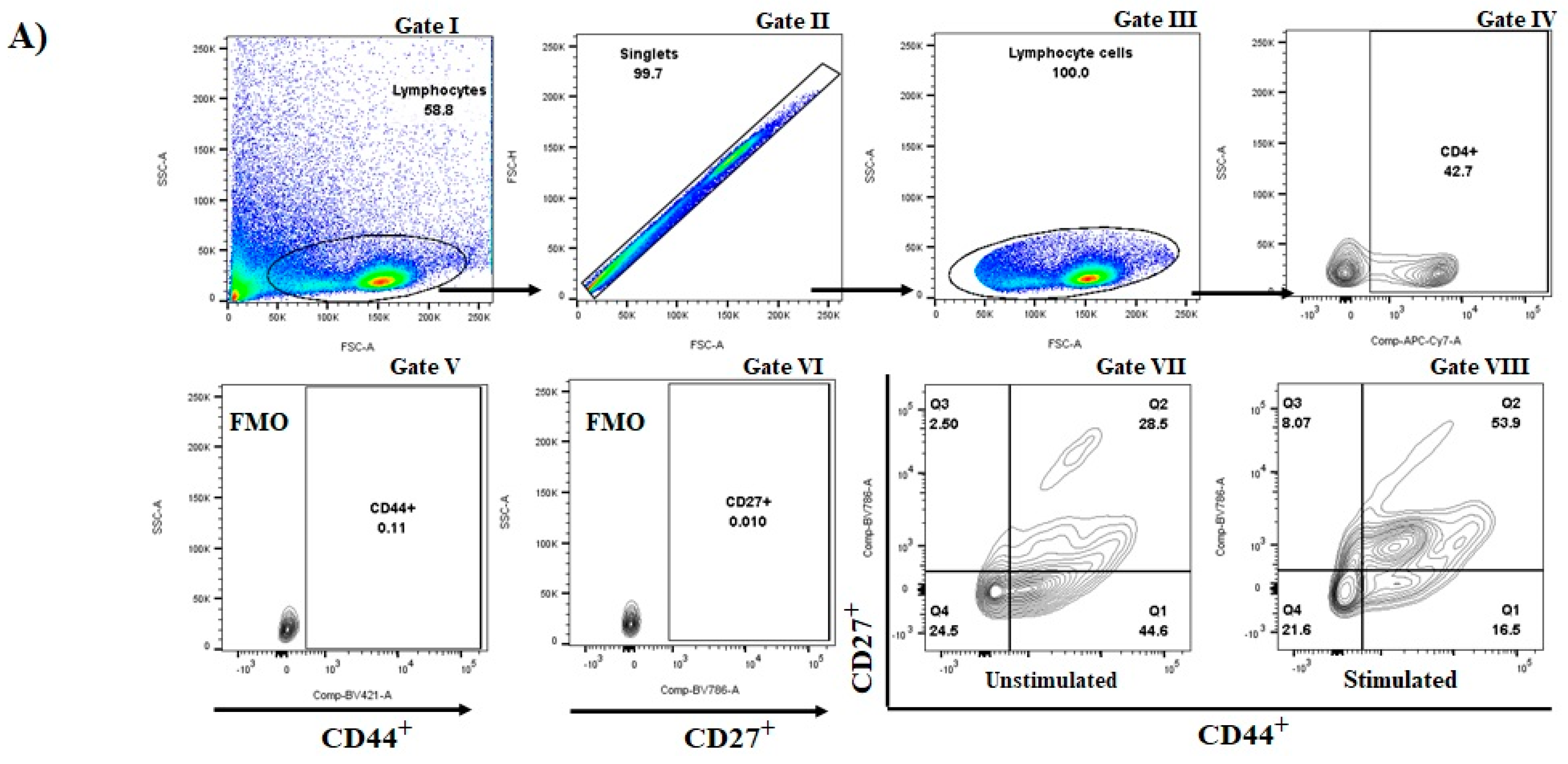
2.7. Immunophenotyping of Ex-Vivo Lymph Nodes Cells
2.8. Preparation of S. aureus Inoculum
2.9. Immunophenotyping and Lymphocyte Proliferation of Cultured Spleen Cells
2.10. Cytokines Measurement
2.11. Statistical Analysis
3. Results
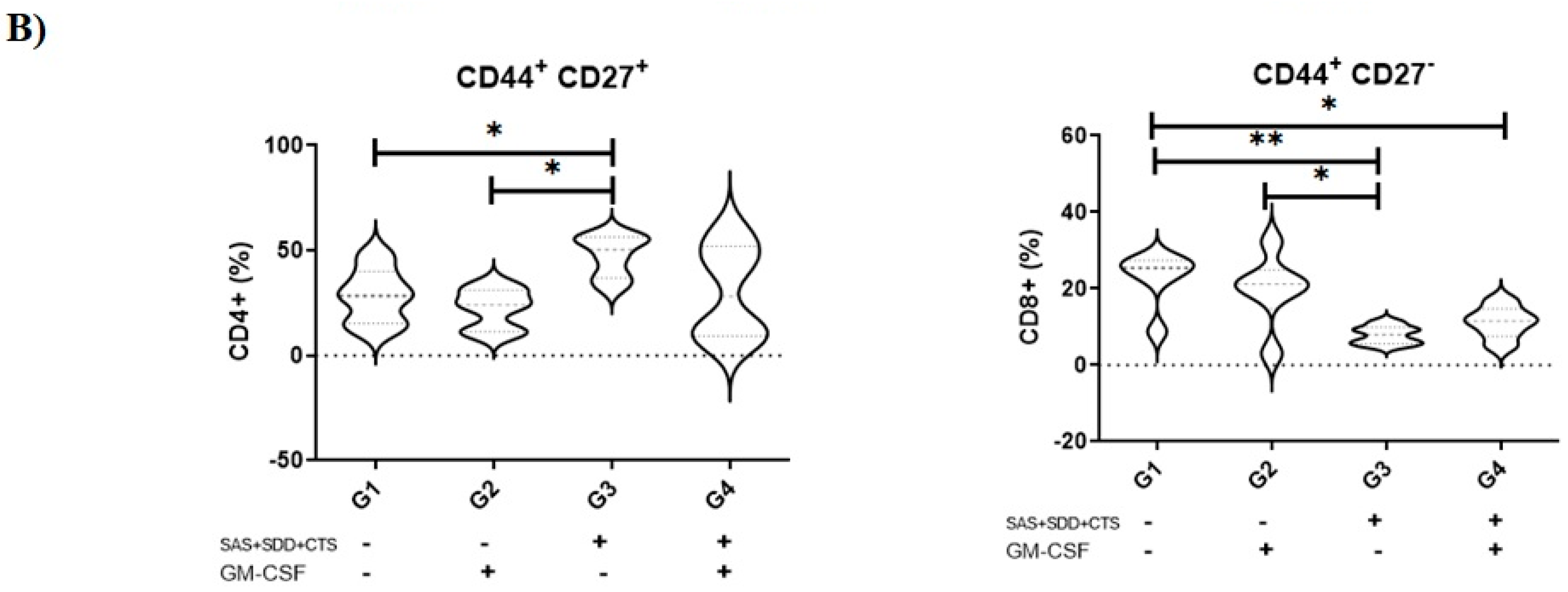
3.1. Immunization with S. aureus Recombinant Proteins Generate Memory Cells in Draining Lymph Nodes
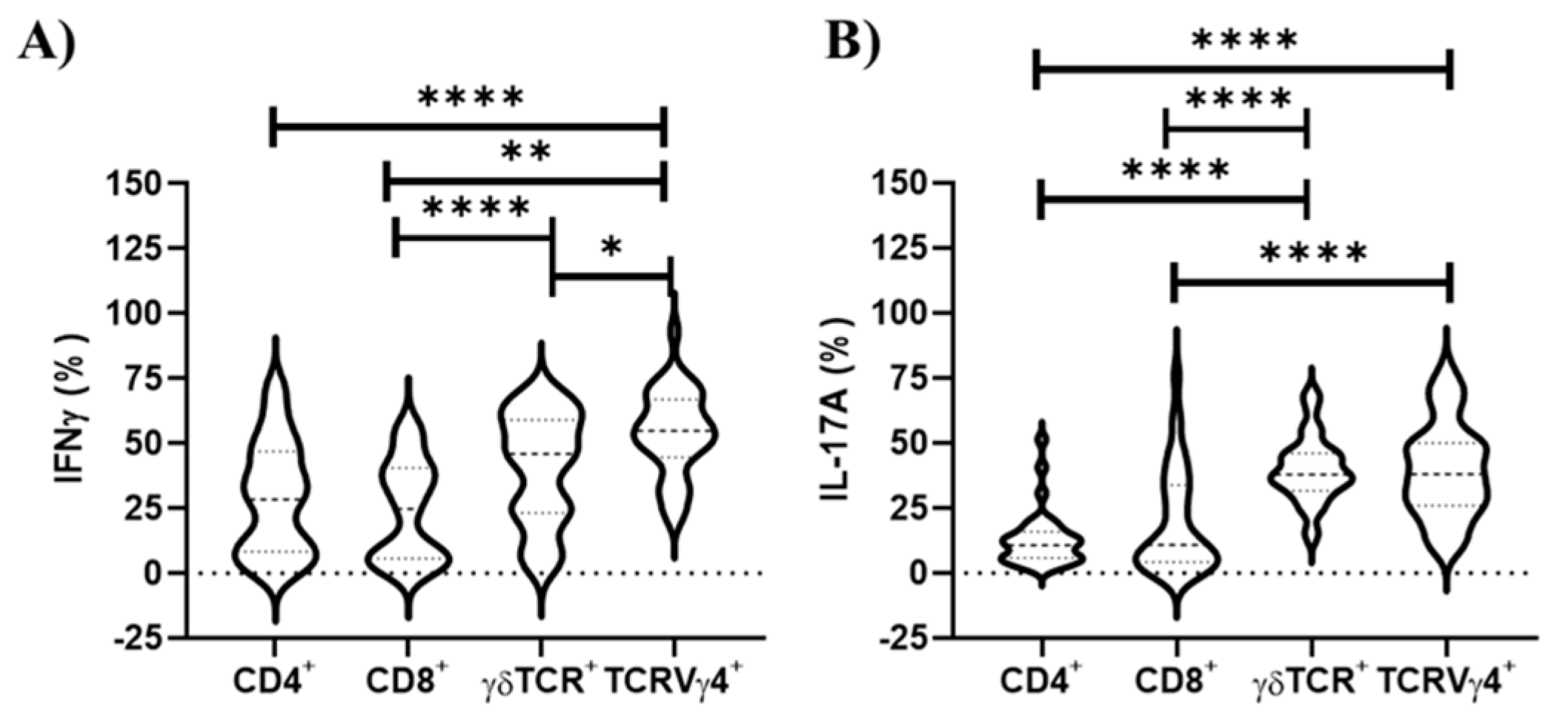
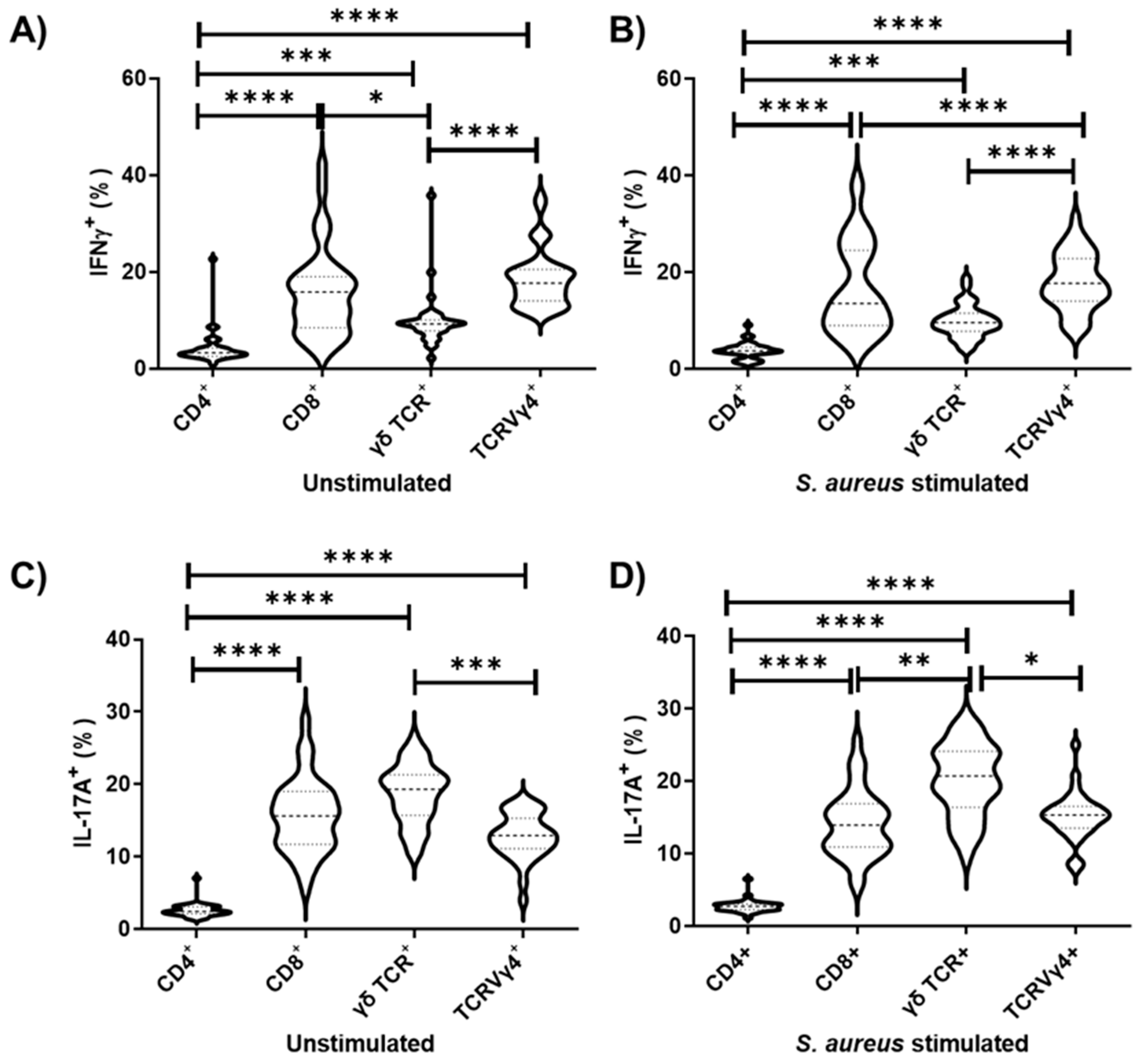
3.2. TCRVγ4+ Cells Are Responsible for the Greatest Contribution for Type 1 and 3 Cell-Mediated Immunities in the Draining Lymph Nodes
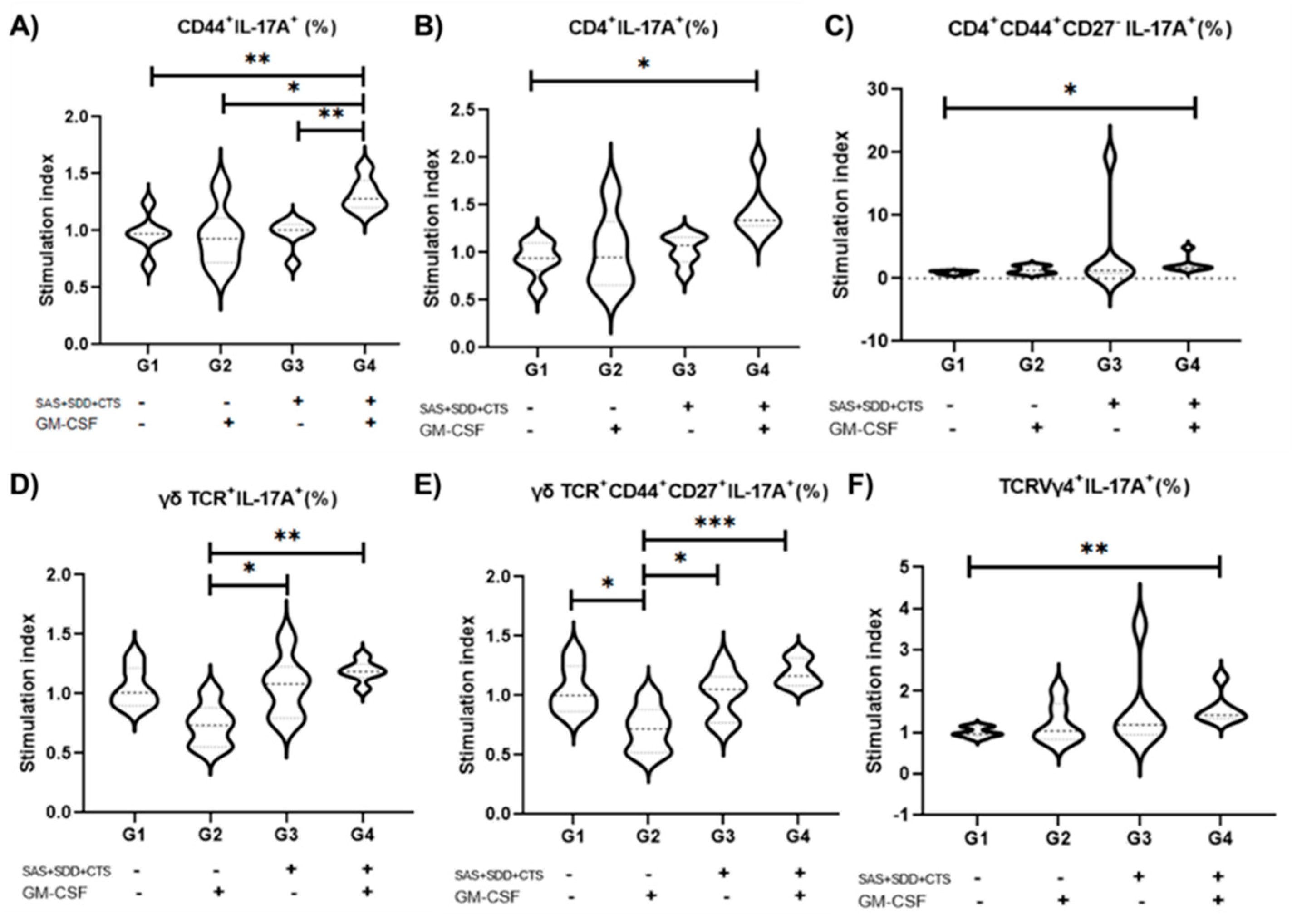
3.3. Immunization with S. aureus Recombinant Proteins Associated with GM-CSF DNA Vaccine Drives Type 3 Recall Immunity That Largely Relies on γδ T and TCRVγ4+ Cells
3.4. TCRVγ4+ Cells and T CD8+ Cells Are the Major Contributors for Type 1 Immunity, While γδ TCR Cells Mainly Support Type 3 Immunity in Splenocyte Cell Culture
3.5. Immunization of Mice with S. aureus Recombinant Proteins Associated with GM-CSF DNA Vaccine DownregulatesTh2 Immunity
3.6. The Combination of S. aureus Recombinant Proteins and GM-CSF Vaccine Induces a Higher Proliferation of Memory T Cells and T CD4+ Cells
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rainard, P.; Foucras, G.; Fitzgerald, J.R.; Watts, J.L.; Koop, G.; Middleton, J.R. Knowledge gaps and research priorities in Staphylococcus aureus mastitis control. Transbound. Emerg. Dis. 2018, 65, 149–165. [Google Scholar] [CrossRef] [Green Version]
- Cunha, A.F.; Andrade, H.M.; Souza, F.N.; Fialho Júnior, L.C.; Rosa, D.L.S.O.; Ramos Sanchez, E.M.; Gidlund, M.; Goto, H.; Brito, M.A.V.P.; Guimarães, A.S.; et al. Comparison of antibody repertories against Staphylococcus aureus in healthy and infected dairy cows with a distinct mastitis history and vaccinated with a polyvalent mastitis vaccine. J. Dairy Sci. 2020, 103, 4588–4605. [Google Scholar] [CrossRef]
- Richardson, E.J.; Bacigalupe, R.; Harrison, E.M.; Weinert, L.A.; Lycett, S.; Vrieling, M.; Robb, K.; Hoskisson, P.A.; Holden, M.T.G.; Feil, E.J.; et al. Gene exchange drives the ecological success of a multi-host bacterial pathogen. Nat. Ecol. Evol. 2018, 2, 1468–1478. [Google Scholar] [CrossRef]
- Zeng, H.; Yang, F.; Feng, Q.; Zhang, J.; Gu, J.; Jing, H.; Cai, C.; Xu, L.; Yang, X.; Xia, X.; et al. Rapid and broad immune efficacy of a recombinant five-antigen vaccine against Staphylococcus aureus infection in animal models. Vaccines 2020, 8, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montgomery, C.P.; Daniels, M.; Zhao, F.; Alegre, M.L.; Chong, A.S.; Daum, R.S. Protective immunity against recurrent Staphylococcus aureus skin infection requires antibody and interleukin-17A. Infect. Immun. 2014, 82, 2125–2134. [Google Scholar] [CrossRef] [Green Version]
- Armentrout, E.I.; Liu, G.Y.; Martins, G.A. T cell immunity and the quest for protective vaccines against Staphylococcus aureus infection. Microorganisms 2020, 8, 1936. [Google Scholar] [CrossRef] [PubMed]
- Rainard, P.; Cunha, P.; Martins, R.P.; Gilbert, F.B.; Germon, P.; Foucras, G. Type 3 immunity: A perspective for the defense of the mammary gland against infections. Vet. Res. 2020, 51, 129. [Google Scholar] [CrossRef] [PubMed]
- Bekeredjian-Ding, I. Deciphering the significance of the T-cell response to Staphylococcus aureus. Future Microbiol. 2017, 12, 1023–1026. [Google Scholar] [CrossRef]
- Zhang, F.; Ledue, O.; Jun, M.; Goulart, C.; Malley, R.; Lu, Y.J. Protection against Staphylococcus aureus colonization and infection by B-and T-cell-mediated mechanisms. mBio 2018, 9, e01949-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cua, D.J.; Tato, C.M. Innate IL-17-producing cells: The sentinels of the immune system. Nat. Rev. Immunol. 2010, 10, 479–489. [Google Scholar] [CrossRef]
- Goldstein, E.J.C.; Proctor, R.A. Challenges for a universal Staphylococcus aureus vaccine. Clin. Infect. Dis. 2012, 54, 1179–1186. [Google Scholar] [CrossRef] [Green Version]
- Proctor, R.A. Is there a future for a Staphylococcus aureus vaccine? Vaccine 2012, 30, 2921–2927. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.G.; O’Keeffe, K.M.; Lalor, S.J.; Maher, B.M.; Mills, K.H.G.; McLoughlin, R.M. Staphylococcus aureus Infection of Mice Expands a Population of Memory γδ T Cells That Are Protective against Subsequent Infection. J. Immunol. 2014, 192, 3697–3708. [Google Scholar] [CrossRef] [Green Version]
- Mahdavi, M.; Tajik, A.H.; Ebtekar, M.; Rahimi, R.; Adibzadeh, M.M.; Moozarmpour, H.R.; Beikverdi, M.S.; Olfat, S.; Hassan, Z.M.; Choopani, M.; et al. Granulocyte-macrophage colony-stimulating factor, a potent adjuvant for polarization to Th-17 pattern: An experience on HIV-1 vaccine model. Apmis 2017, 125, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Nickerson, S.C.; Owens, W.E.; Watts, J.L. Effects of Recombinant Granulocyte Colony-Stimulating Factor on Staphylococcus aureus Mastitis in Lactating Dairy Cows. J. Dairy Sci. 1989, 72, 3286–3294. [Google Scholar] [CrossRef]
- Carneiro, F.R.G.; Silva, T.C.L.; Alves, A.C.; Haline-Vaz, T.; Gozzo, F.C.; Zanchin, N.I.T. Spectroscopic characterization of the tumor antigen NY-REN-21 and identification of heterodimer formation with SCAND1. Biochem. Biophys. Res. Commun. 2006, 343, 260–268. [Google Scholar] [CrossRef]
- Santos, R.P.; Souza, F.N.; Oliveira, A.C.D.; de Souza Filho, A.F.; Aizawa, J.; Moreno, L.Z.; da Cunha, A.F.; Cortez, A.; Della Libera, A.M.M.P.; Heinemann, M.B.; et al. Molecular typing and antimicrobial susceptibility profile of Staphylococcus aureus isolates recovered from bovine mastitis and nasal samples. Animals 2020, 10, 2143. [Google Scholar] [CrossRef]
- Fotoran, W.L.; Santangelo, R.; de Miranda, B.N.M.; Irvine, D.J.; Wunderlich, G. DNA-Loaded Cationic Liposomes Efficiently Function as a Vaccine against Malarial Proteins. Mol. Ther. Methods Clin. Dev. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Soares, A.; Govender, L.; Hughes, J.; Mavakla, W.; de Kock, M.; Barnard, C.; Pienaar, B.; Janse van Rensburg, E.; Jacobs, G.; Khomba, G.; et al. Novel application of Ki67 to quantify antigen-specific in vitro lymphoproliferation. J. Immunol. Methods 2010, 362, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Souza, F.N.; Piepers, S.; Della Libera, A.M.M.P.; Heinemann, M.B.; Cerqueira, M.M.O.P.; De Vliegher, S. Interaction between bovine-associated coagulase-negative staphylococci species and strains and bovine mammary epithelial cells reflects differences in ecology and epidemiological behavior. J. Dairy Sci. 2016, 99, 2867–2874. [Google Scholar] [CrossRef]
- Giunchetti, R.C.; Corrêa-Oliveira, R.; Martins-Filho, O.A.; Teixeira-Carvalho, A.; Roatt, B.M.; de Oliveira Aguiar-Soares, R.D.; de Souza, J.V.; das Dores Moreira, N.; Malaquias, L.C.C.; Mota e Castro, L.L.; et al. Immunogenicity of a killed Leishmania vaccine with saponin adjuvant in dogs. Vaccine 2007, 25, 7674–7686. [Google Scholar] [CrossRef]
- Abdeladhim, M.; Ahmed, M.; Marzouki, S.; Hmida, N.B.; Boussoffara, T.; Hamida, N.B.; Salah, A.; Louzir, H. Human cellular immune response to the saliva of Phlebotomus papatasi is mediated by IL-10-producing CD8+ T cells and TH1-Polarized CD4+ lymphocytes. PLoS Negl. Trop. Dis. 2011, 5, e1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duz, A.L.C.; Vieira, P.M.D.A.; Roatt, B.M.; Aguiar-Soares, R.D.O.; de Oliveira Cardoso, J.M.; de Oliveira, F.C.B.; Reis, L.E.S.; Tafuri, W.L.; Veloso, V.M.; Reis, A.B.; et al. The TcI and TcII Trypanosoma cruzi experimental infections induce distinct immune responses and cardiac fibrosis in dogs. Mem. Inst. Oswaldo Cruz 2014, 109, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Mackroth, M.S.; Abel, A.; Steeg, C.; Schulze zur Wiesch, J.; Jacobs, T. Acute Malaria Induces PD1+CTLA4+ Effector T Cells with Cell-Extrinsic Suppressor Function. PLoS Pathog. 2016, 12, e1005909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, S.E.; Zhou, Z.; Landry, L.G.; Anderson, A.M.; Alkanani, A.K.; Fischer, J.; Peakman, M.; Mallone, R.; Campbell, K.; Michels, A.W.; et al. Multiplex T Cell Stimulation Assay Utilizing a T Cell Activation Reporter-Based Detection System. Front. Immunol. 2020, 11, 9–23. [Google Scholar] [CrossRef]
- Schumann, J.; Stanko, K.; Schliesser, U.; Appelt, C.; Sawitzki, B. Differences in CD44 surface expression levels and function discriminates IL-17 and IFN-γ producing helper T cells. PLoS ONE 2015, 10, e0132479. [Google Scholar] [CrossRef] [Green Version]
- van Faassen, H.; Saldanha, M.; Gilbertson, D.; Dudani, R.; Krishnan, L.; Sad, S. Reducing the Stimulation of CD8 + T Cells during Infection with Intracellular Bacteria Promotes Differentiation Primarily into a Central (CD62L high CD44 high) Subset. J. Immunol. 2005, 174, 5341–5350. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, L.; Deschatelets, L.; Stark, F.C.; Gurnani, K.; Sprott, G.D. Archaeosome adjuvant overcomes tolerance to tumor-associated melanoma antigens inducing protective CD8+ T cell responses. Clin. Dev. Immunol. 2010. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.S.; Fowler, V.G.; Shukla, S.K.; Rose, W.E.; Proctor, R.A. Development of a vaccine against Staphylococcus aureus invasive infections: Evidence based on human immunity, genetics and bacterial evasion mechanisms. FEMS Microbiol. Rev. 2020, 44, 123–153. [Google Scholar] [CrossRef] [Green Version]
- Sallusto, F.; Geginat, J.; Lanzavecchia, A. Central memory and effector memory T cell subsets: Function, generation, and maintenance. Annu. Rev. Immunol. 2004, 22, 745–763. [Google Scholar] [CrossRef]
- Kedzierska, K.; Curtis, J.M.; Valkenburg, S.A.; Hatton, L.A.; Kiu, H.; Doherty, P.C.; Kedzierski, L. Induction of protective CD4+ T cell-mediated immunity by a Leishmania peptide delivered in recombinant influenza viruses. PLoS ONE 2012, 7, e33161. [Google Scholar] [CrossRef] [Green Version]
- Jameson, S.C.; Masopust, D. Understanding Subset Diversity in T Cell Memory. Immunity 2018, 48, 214–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, M.E.; Bertrand, B.P.; Heim, C.E.; Alqarzaee, A.A.; Chaudhari, S.S.; Aldrich, A.L.; Fey, P.D.; Thomas, V.C.; Kielian, T. Staphylococcus aureus atp synthase promotes biofilm persistence by influencing innate immunity. mBio 2020, 11, e01581-20. [Google Scholar] [CrossRef]
- Liu, L.; Beck, C.; Nøhr-Meldgaard, K.; Peschel, A.; Kretschmer, D.; Ingmer, H.; Vestergaard, M. Inhibition of the ATP synthase sensitizes Staphylococcus aureus towards human antimicrobial peptides. Sci. Rep. 2020, 10, 11391. [Google Scholar] [CrossRef] [PubMed]
- Nagaev, I.; Björkman, J.; Andersson, D.I.; Hughes, D. Biological cost and compensatory evolution in fusidic acid-resistant Staphylococcus aureus. Mol. Microbiol. 2001, 40, 433–439. [Google Scholar] [CrossRef]
- Bharathan, M.; Mullarky, I.K. Targeting mucosal immunity in the battle to develop a mastitis vaccine. J. Mammary Gland. Biol. Neoplasia 2011, 16, 409–419. [Google Scholar] [CrossRef]
- Bröker, B.; Mrochen, D.; Péton, V. The T Cell Response to Staphylococcus aureus. Pathogens 2016, 5, 31. [Google Scholar] [CrossRef]
- Souza, F.N.; Leiva, T.; Rodrigues, R.O.; Gandin Júnior, J.R.; Drago, E.; Gomes, R.C.; Santos, K.R.; Blagitz, M.G.; Della Libera, A.M.M.P.; Vasconcelos, J.L.M.; et al. Lymphocyte proliferative responses in dairy cows supplemented with an immunomodulatory feed additive and administered polyvalent vaccination. Arq. Bras. Med. Vet. Zootec. 2020, 72, 2397–2401. [Google Scholar] [CrossRef]
- Baldwin, C.L.; Yirsaw, A.; Gillespie, A.; Le Page, L.; Zhang, F.; Damani-Yokota, P.; Telfer, J.C. γδ T cells in livestock: Responses to pathogens and vaccine potential. Transbound. Emerg. Dis. 2020, 67, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Buza, J.; Kiros, T.; Zerihun, A.; Abraham, I.; Ameni, G. Vaccination of calves with Mycobacteria bovis Bacilli Calmete Guerin (BCG) induced rapid increase in the proportion of peripheral blood γδ T cells. Vet. Immunol. Immunopathol. 2009, 130, 251–255. [Google Scholar] [CrossRef]
- Kabelitz, D. γδ T-cells: Cross-talk between innate and adaptive immunity. Cell. Mol. Life Sci. 2011, 68, 2331–2333. [Google Scholar] [CrossRef]
- Spellberg, B.; Daum, R. A new view on development of a Staphylococcus aureus vaccine: Insights from mice and men. Hum. Vaccin. 2010, 6, 857–859. [Google Scholar] [CrossRef] [Green Version]
- Jing, X.; Cao, D.; Liu, H.; Wang, X.; Chen, X.Z.; Chen, D. Pivotal Role of IL-17-producing γδ T cells in Mouse Chronic Mastitis Experimentally Induced with Staphylococcus aureus. Asian J. Anim. Vet. Adv. 2012, 7, 1266–1278. [Google Scholar] [CrossRef] [Green Version]
- Paape, M.J.; Shafer-Weaver, K.; Capuco, A.V.; Van Oostveldt, K.; Burvenich, C. Immune surveillance of mammary tissue by phagocytic cells. Adv. Exp. Med. Biol. 2000, 480, 259–277. [Google Scholar] [CrossRef]
- Cho, J.S.; Pietras, E.M.; Garcia, N.C.; Ramos, R.I.; Farzam, D.M.; Monroe, H.R.; Magorien, J.E.; Blauvelt, A.; Kolls, J.K.; Cheung, A.L.; et al. IL-17 is essential for host defense against cutaneous Staphylococcus aureus infection in mice. J. Clin. Investig. 2010, 120, 1762–1773. [Google Scholar] [CrossRef] [Green Version]
- Maher, B.M.; Mulcahy, M.E.; Murphy, A.G.; Wilk, M.; O’Keeffe, K.M.; Geoghegan, J.A.; Lavelle, E.C.; McLoughlin, R.M. Nlrp-3-driven interleukin 17 production by γδT cells controls infection outcomes during Staphylococcus aureus surgical site infection. Infect. Immun. 2013, 81, 4478–4489. [Google Scholar] [CrossRef] [Green Version]
- Marchitto, M.C.; Dillen, C.A.; Liu, H.; Miller, R.J.; Archer, N.K.; Ortines, R.V.; Alphonse, M.P.; Marusina, A.I.; Merleev, A.A.; Wang, Y.; et al. Clonal Vγ6+Vδ4+ T cells promote IL-17–mediated immunity against Staphylococcus aureus skin infection. Proc. Natl. Acad. Sci. USA 2019, 166, 10917–10926. [Google Scholar] [CrossRef] [Green Version]
- Rainard, P.; Cunha, P.; Bougarn, S.; Fromageau, A.; Rossignol, C.; Gilbert, F.B.; Berthon, P. T Helper 17-Associated Cytokines Are Produced during Antigen-Specific Inflammation in the Mammary Gland. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Kehrli, M.E.; Cullor, J.S.; Nickerson, S.C. Immunobiology of Hematopoietic Colony-Stimulating Factors: Potential Application to Disease Prevention in the Bovine. J. Dairy Sci. 1991, 74, 4399–4412. [Google Scholar] [CrossRef]
- Bougarn, S.; Cunha, P.; Gilbert, F.B.; Harmache, A.; Foucras, G.; Rainard, P. Staphylococcal-associated molecular patterns enhance expression of immune defense genes induced by IL-17 in mammary epithelial cells. Cytokine 2011, 56, 749–759. [Google Scholar] [CrossRef]
- Warren, T.L.; Weiner, G.J. Uses of granulocyte-macrophage colony-stimulating factor in vaccine development. Curr. Opin. Hematol. 2000, 7, 168–173. [Google Scholar] [CrossRef]
- Stabel, J.R.; Kehrli, M.E.; Thurston, J.R.; Goff, J.P.; Boone, T.C. Granulocyte Colony-Stimulating Factor Effects on Lymphocytes and Immunoglobulin Concentrations in Periparturient Cows. J. Dairy Sci. 1991, 74, 3755–3762. [Google Scholar] [CrossRef]
- Kimura, K.; Goff, J.P.; Canning, P.; Wang, C.; Roth, J.A. Effect of recombinant bovine granulocyte colony-stimulating factor covalently bound to polyethylene glycol injection on neutrophil number and function in periparturient dairy cows. J. Dairy Sci. 2014, 97, 4842–4851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutton, C.E.; Lalor, S.J.; Sweeney, C.M.; Brereton, C.F.; Lavelle, E.C.; Mills, K.H.G. Interleukin-1 and IL-23 Induce Innate IL-17 Production from γδ T Cells, Amplifying Th17 Responses and Autoimmunity. Immunity 2009, 31, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Korn, T.; Petermann, F. Development and function of interleukin 17-producing γδ T cells. Ann. N. Y. Acad. Sci. 2012, 1247, 34–45. [Google Scholar] [CrossRef]
- Dillen, C.A.; Pinsker, B.L.; Marusina, A.I.; Merleev, A.A.; Farber, O.N.; Liu, H.; Archer, N.K.; Lee, D.B.; Wang, Y.; Ortines, R.V.; et al. Clonally expanded γδ T cells protect against Staphylococcus aureus skin reinfection. J. Clin. Investig. 2018, 128, 1026–1042. [Google Scholar] [CrossRef]
- Cooper, A.J.R.; Lalor, S.J.; McLoughlin, R.M. Activation of Human Vδ2 + γδ T Cells by Staphylococcus aureus Promotes Enhanced Anti-Staphylococcal Adaptive Immunity. J. Immunol. 2020, 205, 1039–1049. [Google Scholar] [CrossRef]
- Schluns, K.S.; Lefrançois, L. Cytokine control of memory T-cell development and survival. Nat. Rev. Immunol. 2003, 3, 269–279. [Google Scholar] [CrossRef]
- Woodland, D.L.; Kohlmeier, J.E. Migration, maintenance and recall of memory T cells in peripheral tissues. Nat. Rev. Immunol. 2009, 9, 153–161. [Google Scholar] [CrossRef]
- Ahlers, J.D.; Belyakov, I.M. Memories that last forever: Strategies for optimizing vaccine T-cell memory. Blood 2010, 115, 1678–1689. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mAbs | Fluorescent Probes | Target | Clone | Host | Concentration (mg mL−1) | Cat. n. |
|---|---|---|---|---|---|---|
| Anti-CD4 1 | APC-Cy7 | Mouse | GK1.5 | Rat | 0.2 | 552051 |
| Anti-CD8 1 | BV510 | Mouse | 53-6.7 | Rat | 0.2 | 563068 |
| Anti-CD44 1 | BV421 | Mouse | IM7 | Rat | 0.2 | 563970 |
| Anti-CD27 1 | BV750 | Mouse | LG.3A10 | Hamster | 0.2 | 747399 |
| Anti-CD19 1 | PE-Cy7 | Mouse | 1D3 | Rat | 0.2 | 552854 |
| Anti-γδ TCR 1 | BV650 | Mouse | GL3 | Hamster | 0.2 | 563993 |
| Anti-TCRVγ4 1 | FITC | Mouse | GL2 | Hamster | 0.2 | 552143 |
| Anti-IL-17° 1 | Alexa 700 | Mouse | TC11-18H10 | Rat | 0.2 | 560820 |
| Anti-IFN-γ 1 | PE | Mouse | XMG1.2 | Rat | 0.2 | 554412 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, K.R.; Souza, F.N.; Ramos-Sanchez, E.M.; Batista, C.F.; Reis, L.C.; Fotoran, W.F.; Heinemann, M.B.; Goto, H.; Gidlund, M.; Cunha, A.F.; et al. Staphylococcus aureus Protection-Related Type 3 Cell-Mediated Immune Response Elicited by Recombinant Proteins and GM-CSF DNA Vaccine. Vaccines 2021, 9, 899. https://doi.org/10.3390/vaccines9080899
Santos KR, Souza FN, Ramos-Sanchez EM, Batista CF, Reis LC, Fotoran WF, Heinemann MB, Goto H, Gidlund M, Cunha AF, et al. Staphylococcus aureus Protection-Related Type 3 Cell-Mediated Immune Response Elicited by Recombinant Proteins and GM-CSF DNA Vaccine. Vaccines. 2021; 9(8):899. https://doi.org/10.3390/vaccines9080899
Chicago/Turabian StyleSantos, Kamila R., Fernando N. Souza, Eduardo M. Ramos-Sanchez, Camila F. Batista, Luiza C. Reis, Wesley F. Fotoran, Marcos B. Heinemann, Hiro Goto, Magnus Gidlund, Adriano F. Cunha, and et al. 2021. "Staphylococcus aureus Protection-Related Type 3 Cell-Mediated Immune Response Elicited by Recombinant Proteins and GM-CSF DNA Vaccine" Vaccines 9, no. 8: 899. https://doi.org/10.3390/vaccines9080899