
Mapping Global Prevalence of Acinetobacter baumannii and Recent Vaccine Development to Tackle It


Abstract
:1. Introduction
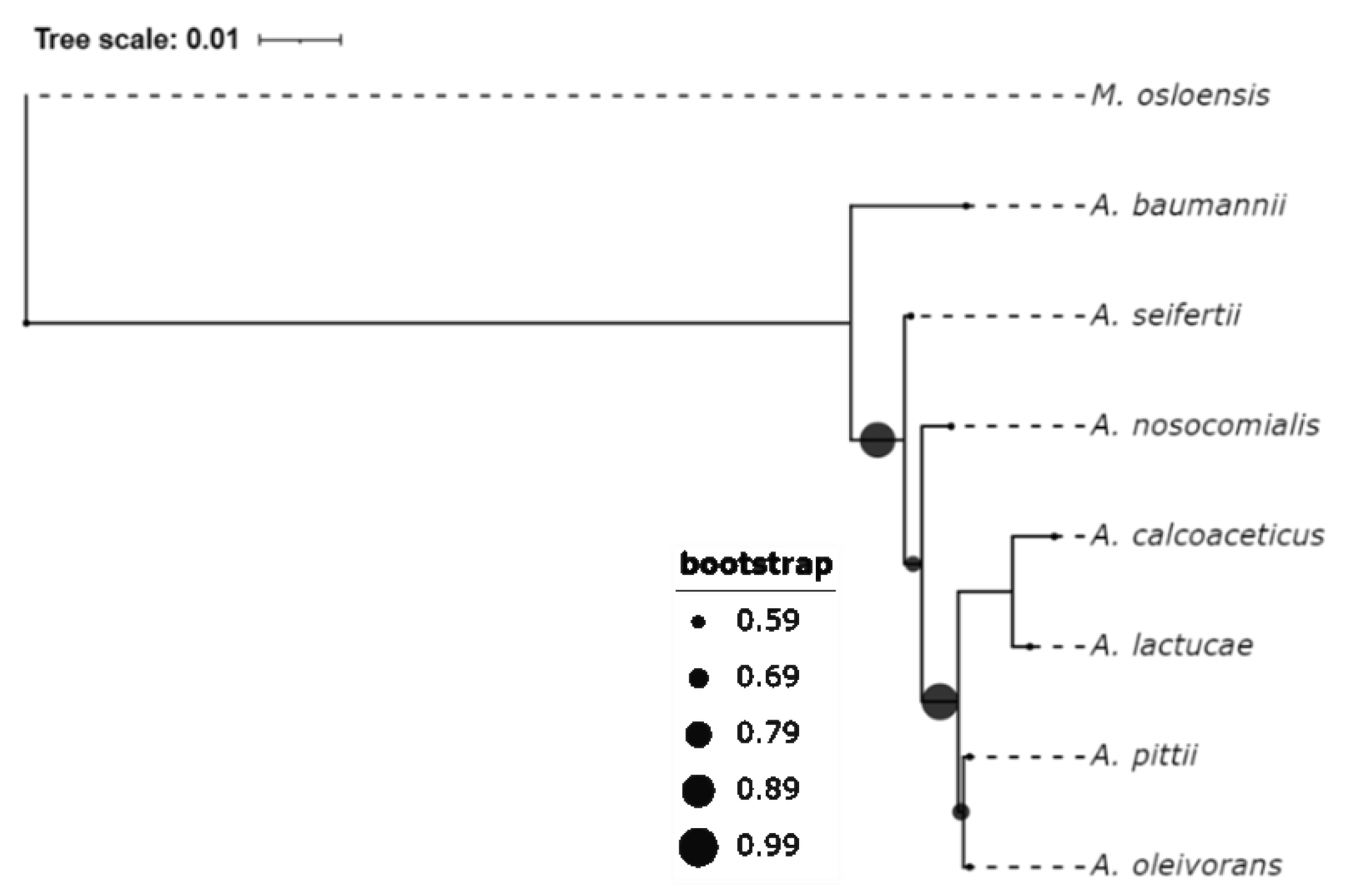
2. A. baumannii Is Ubiquitous
3. Antimicrobial Resistance
4. Clinical Importance
4.1. Susceptible Populations
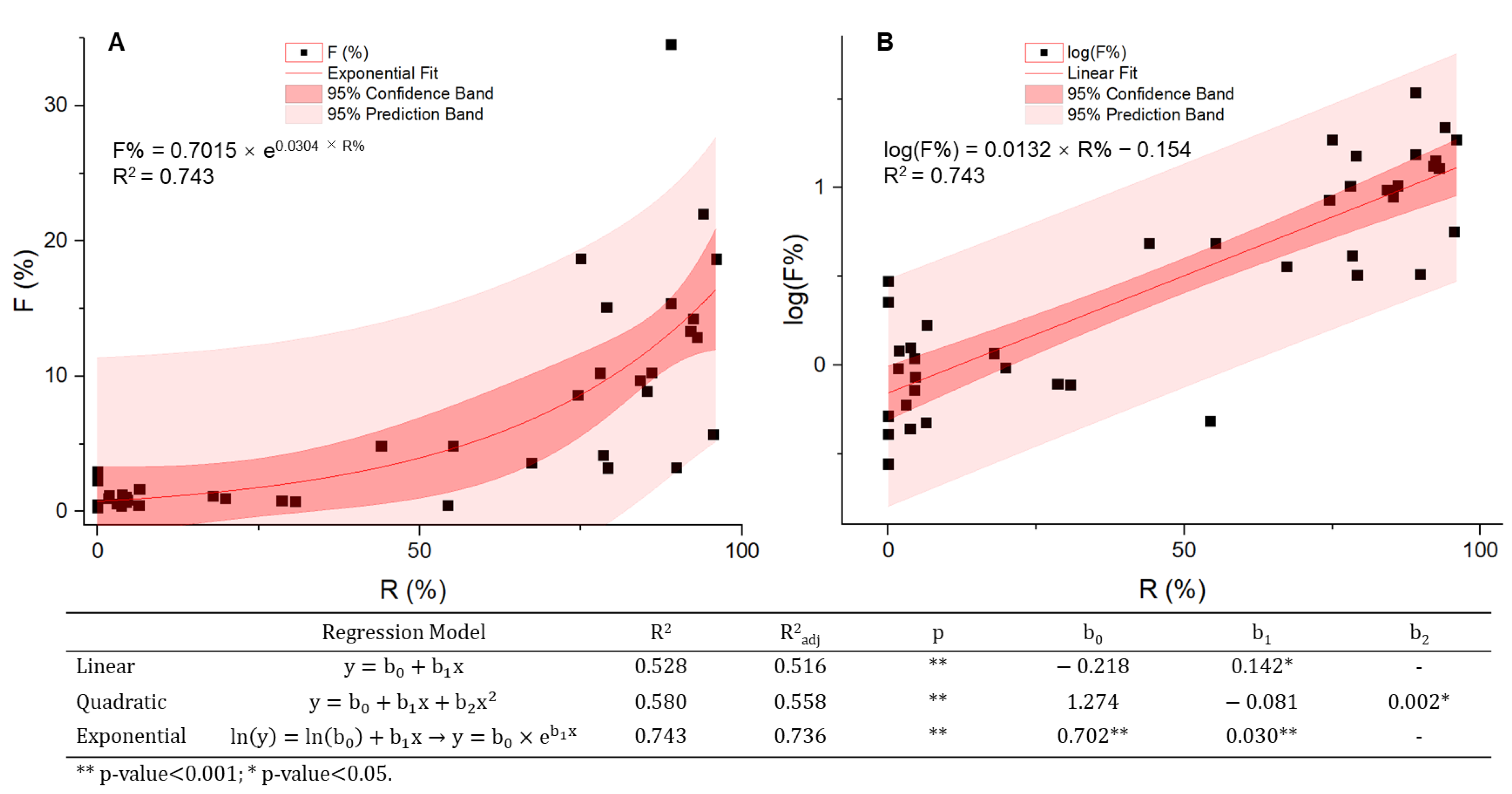
4.2. Prevalence
5. Virulence Factors
5.1. Adhesins and Invasins
5.2. Cytotoxins
5.3. Persistence

{kind=link}
{kind=link}
{kind=link}
| Virulence Factors | Functions Involved | Adhesion | Invasion | Cytotoxicity | Ref |
|---|---|---|---|---|---|
| Ata | type Vc secretion system | + | + | + | [90,91] |
| Bap | type I secretion system | + | [88] | ||
| BasD, BauA | iron acquisition system | + | + | [130] | |
| BfmR/S | two-component regulatory system | + | + | [93,94] | |
| BLP-1, BLP-2 | Bap like protein | + | [131] | ||
| Capsular polysaccharides | outer membrane component | + | [117] | ||
| CdiA/CdiB | type Vb secretion system | + | [132] | ||
| ChoP | phosphorylcholine | + | + | [103] | |
| CipA | plasminogen binding protein | + | [124] | ||
| FhaB/FhaC | type Vb secretion system | + | [89] | ||
| GGT | type II secretion system | + | + | [113] | |
| LOS | outer membrane component | + | [106,107] | ||
| LpsB | LOS production | + | [107] | ||
| LpxA, LpxC, LpxD | LOS production | + | [107,108,109,110] | ||
| NfuA | iron acquisition system | + | + | [101,120,122] | |
| OmpA | porin | + | + | + | [82,83] |
| Omp33-36 | porin | + | + | + | [86,87] |
| paaE | production of toxic epoxide compounds | + | [129,133] | ||
| PLC, PLD | phospholipase | + | [100,101,102] | ||
| TrxA | type 4 pili production | + | [98] | ||
| Type 4 pili | motility apparatus | + | [96] | ||
| T6SS | type VI secretion system | + | + | [114,134] |
6. Anti-A. baumannii Vaccine Development
6.1. Target Populations
6.2. Vaccine Candidates
6.2.1. Whole-Cell Vaccines
6.2.2. Subunit Vaccines
| Platform and Immunogen | Mice Strain | Immunisation Schedule | In Vivo Challenge Study | Immune Responses | Ref. | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Day | Route | Dose | Chall. Strain(s) | Model (Route) | Dose * (CFU) (LD50) | Survival (%) | Survival monitored (days post challenge) | ||||
| Live Attenuated Vaccines | |||||||||||
| Ci79 ΔtrxA | C57BL/6 (4w–6w) | 0, 14 | i.p. / s.c. | 2 × 105 CFU | WT Ci79 | day 28 sepsis (i.p.) | 5 × 106 (10 × LD50) | i.p.—100% s.c.—90% | 28 days | robust humoral responses, minimal cellular immunity | [139] |
| 1 × 106 (2 × LD50) | i.p.—100% s.c.—100% | ||||||||||
| Inactivated Vaccines | |||||||||||
| formalin-killed ATCC 19606 with AlPO4 | C57BL/6 (6w–8w) | 0, 21 | i.m. | 1 × 108 CFU | ATCC 19606 Ab-154 113-16 | day 28 sepsis (i.p.) | 1.6 × 106 (242 × LD50) 3.1 × 106 (6.5 × LD50) 4.1 × 106 (74.5 × LD50) | 100% 100% ~85% | 7 days | significant levels of IgG and IgM, Th2- and Th1-mediated responses | [137] |
| formalin-killed LAC-4 | C57BL/6 BALB/c (8w–12w) | 0, 14, 21 | i.n. | 5 × 107 CFU | LAC-4 | day 42 sepsis (i.p.) pneumonia (i.n.) | 8.4 × 105 (i.p.) 7 × 107 (i.n.) | 100% | 10 days | mucosal immune responses, dominant Th2 responses | [141] |
| formalin-killed IB010 (ATCC 19606 ΔlpxD) with AlPO4 | C57BL/6 (6w–8w) | 0, 14 | i.m. | 1 × 109 CFU | ATCC 19606 Ab-154 | day 21 sepsis (i.p.) | 2.3 × 106 (341 × LD50) 1.1 × 106 (2.2 × LD50) | 100% 100% | 7 days | robust IgG1 and IgG2c responses | [142] |
| Subunit Vaccines | |||||||||||
| rOmpA (ATCC 19606) conjugated with cholera toxin | BALB/c (6w–8w) | 0, 21 | i.n. | 10 μg | strain A strain B strain C strain D strain E | day 28 sepsis (i.p.) | 5 × 108 (2.5 × LD50) 2 × 108 (2 × LD50) 5 × 107 (2 × LD50) 3 × 108 (3 × LD50) 3 × 107 (3 × LD50) | ~50% ~40% ~55% 100% ~70% | 15 days | significant levels of IgG and IgA | [146] |
| rOmpA (ATCC 17978) with Al(OH)3 | diabetic BALB/c (>6 m and 6w–10w) | 0, 21 | s.c. | 3 μg | HUMC 1 | day 35 sepsis (i.v.) | 2 × 107 | ~50% | 28 days | robust humoral responses | [147] |
| rBap (Kh0060) with FA | BALB/c (4w–6w) | 0, 14,28 | not stated | 10 μg | Kh0060 | day 35 sepsis (i.p.) | 1 × 109 (100 × LD50) 1 × 1011 (104 × LD50) 1 × 1013 (106 × LD50) | 100% 80% 60% | 5 days | robust humoral responses | [154] |
| rBauA or rBfnH (ATCC 19606) with FA | BALB/c (6w–8w) | 0, 14,28 | s.c. | 20 μg | ATCC 19606 | day 49 sepsis (i.p.) | 1.5 × 107 (2 × LD50) | ~40% | 7 days | not specified | [155] |
| rBamA (ATCC 19606) with Al(OH)3 | BALB/c (6w–8w) | 0, 14,28 | i.p. | 20 μg | P-562 | day 45 pneumonia (i.n.) | 1 × 109 | ~80% | 7 days | non-neutralising, opsonic IgG | [156] |
| rBamA (ATCC 19606] with FA | BALB/c (6w–8w] | 0, 14, 28, 42 | s.c. | 20 μg | ATCC 19606 | day unknown, sepsis (i.p.) | 2 × 106 (4 × LD50) 7 × 106 (14 × LD50) | 100% 25% | 4 days | not specified | [157] |
| rTrx-OmpW (ATCC 17978) with alum | ICR (6w–8w) | 0, 14,28 | s.c. | 50 μg | ATCC 17978 | day 49 sepsis (i.p.) | 1 × 106 | 100% | 7 days | opsonic IgG, complements | [158] |
| rTrx-Omp22 (ATCC 17978) | ICR (6w–8w) | 0, 14, 28 | s.c. | 50 μg 20 μg 10 μg 5 μg | Ab1 | day 49 sepsis (i.p.) | 1 × 106 | 100% 33% 33% 0 | 7 days | opsonic IgG, complements | [159] |
| rOmp22 (ATCC 19606) with FA | BALB/c (6w–8w) | 0, 14,21 | s.c. | 20 μg | ATCC 19606 | day 42 sepsis (i.p.) | 2 × 108 | 37.5% | 8 days | not specified | [160] |
| rOmpK (ATCC 19606) with FA | BALB/c (6w–8w) | 0, 14,21 | s.c. | 20 μg | ATCC 19606 | day 42 sepsis (i.p.) | 2 × 108 | 25% | 8 days | not specified | [160] |
| rOmp22-OmpK (ATCC 19606) with FA | BALB/c (6w–8w) | 0, 14,21 | s.c. | 20 μg | ATCC 19606 | day 42 sepsis (i.p.) | 2 × 108 | 67% | 8 days | not specified | [160] |
| rOmp22-OmpK (ATCC 19606) with MF59 adjuvant | BALB/c (6w–8w) | 0, 14,21 | i.t. | 30 μg | ATCC 19606 | day 42 respiratory (i.t.) | 1 × 108 | 83.3% | 10 days | significant levels of IgG and IgA | [161] |
| Multi-Component Vaccines | |||||||||||
| OMC (ATCC 19606) with AlPO4 | C57BL/6 (6w–8w) | 0, 21 | i.m. | 25 μg | ATCC 19606 Ab-154 113-16 | day 35 sepsis (i.p.) | 1 × 106 (151.3 × LD50) 1 × 107 (20.9 × LD 50) 1.5 × 106 (27.3 × LD50) | 100% 80% 100% | 7 days | robust IgG and IgM responses | [136] |
| OMV (ATCC 19606) with AlPO4 | C57BL/6 (6w–8w) | 0, 14 | i.m. | 10 μg | ATCC 19606 Ab-154 113-16 | day 35 sepsis (i.p.) | 4.5 × 105 (71.3 × LD50) 1.7 × 106 (4.3 × LD50) 2 × 106 (39.9 × LD50) | 100% ~90% 100% | 7 days | robust IgG1, IgG2c and IgM responses | [138] |
| ClyA-Omp22 (ATCC 17978) with E. coli derived OMV | ICR (6w–8w) | 0, 14 | s.c. | 5 μg 10 μg 20 μg 50 μg | Ab1 | day 35 sepsis (i.p.) | 1 × 106 | 30% 40% 63.6% 100% | 7 days | opsonic IgG | [162] |
| Nucleic Acid Vaccines | |||||||||||
| pVAX1-ompA (LAC-4) with CpG-ODN adjuvant | C57BL/6 (6w–8w) | 0, 14, 21 | i.m. | 100 μg | LAC-4 SJZ04 SJZ18 SJZ28 | day 28 respiratory (i.t.) | 1.2 × 107 (2 × LD50) 1 × 108 1 × 108 1 × 108 | ~50% ~40% ~60% ~50% | 7 days | robust humoral responses, mixed Th1/Th2/Th17 cellular responses | [163] |
| pBudCE4.1-ompA | BALB/c (6w–8w) | 0, 7, 14, 28 | i.m. | 25 μg | not stated | day 35 respiratory (i.n.) | 1 × 108 | 60% | 15 days | significant levels of IgG and IgM | [164] |
6.2.3. LOS and Conjugate Vaccines
6.2.4. Multi-Component Vaccines
6.2.5. Nucleic Acid Vaccines
6.3. Lessons from A. baumannii Vaccines and Future Directions
6.3.1. Modern Vaccine Technologies
6.3.2. Long-Term Cellular Immunity
6.3.3. Multivalent Protection
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [Green Version]
- Antunes, L.C.; Visca, P.; Towner, K.J. Acinetobacter baumannii: Evolution of a global pathogen. Pathog. Dis. 2014, 71, 292–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulens, S.N.; Yi, S.H.; Walters, M.S.; Jacob, J.T.; Bower, C.; Reno, J.; Wilson, L.; Vaeth, E.; Bamberg, W.; Janelle, S.J.; et al. Carbapenem-nonsusceptible Acinetobacter baumannii, 8 US metropolitan areas, 2012–2015. Emerg. Infect. Dis. 2018, 24, 727–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inchai, J.; Pothirat, C.; Bumroongkit, C.; Limsukon, A.; Khositsakulchai, W.; Liwsrisakun, C. Prognostic factors associated with mortality of drug-resistant Acinetobacter baumannii ventilator-associated pneumonia. J. Intensive Care 2015, 3, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisplinghoff, H.; Paulus, T.; Lugenheim, M.; Stefanik, D.; Higgins, P.G.; Edmond, M.B.; Wenzel, R.P.; Seifert, H. Nosocomial bloodstream infections due to Acinetobacter baumannii, Acinetobacter pittii and Acinetobacter nosocomialis in the United States. J. Infect. 2012, 64, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Metan, G.; Alp, E.; Aygen, B.; Sumerkan, B. Acinetobacter baumannii meningitis in post-neurosurgical patients: Clinical outcome and impact of carbapenem resistance. J. Antimicrob. Chemother. 2007, 60, 197–199. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Wendt, C.; Dietze, B.; Dietz, E.; Rüden, H. Survival of Acinetobacter baumannii on dry surfaces. J. Clin. Microbiol. 1997, 35, 1394–1397. [Google Scholar] [CrossRef] [Green Version]
- Chen, W. Current advances and challenges in the development of Acinetobacter vaccines. Hum. Vaccines Immunother. 2015, 11, 2495–2500. [Google Scholar] [CrossRef] [Green Version]
- Weiss, N.; Kracht, M.; Gleim, D. List of Prokaryotic Names with Standing in Nomenclature-Genus Acinetobacter. 2021. Available online: https://lpsn.dsmz.de/genus/acinetobacter (accessed on 24 May 2021).
- Rossau, R.; Van Landschoot, A.; Gillis, M.; De Ley, J. Taxonomy of Moraxellaceae fam. nov., a new bacterial family to accommodate the genera Moraxella, Acinetobacter, and Psychrobacter and related organisms. Int. J. Syst. Bacteriol. 1991, 41, 310–319. [Google Scholar] [CrossRef]
- Smith, M.G.; Gianoulis, T.A.; Pukatzki, S.; Mekalanos, J.J.; Ornston, L.N.; Gerstein, M.; Snyder, M. New insights into Acinetobacter baumannii pathogenesis revealed by high-density pyrosequencing and transposon mutagenesis. Genes Dev. 2007, 21, 601–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemec, A.; Krizova, L.; Maixnerova, M.; van der Reijden, T.J.; Deschaght, P.; Passet, V.; Vaneechoutte, M.; Brisse, S.; Dijkshoorn, L. Genotypic and phenotypic characterization of the Acinetobacter calcoaceticus—Acinetobacter baumannii complex with the proposal of Acinetobacter pittii sp. nov. (formerly Acinetobacter genomic species 3) and Acinetobacter nosocomialis sp. nov. (formerly Acinetobacter genomic species 13TU). Res. Microbiol. 2011, 162, 393–404. [Google Scholar] [CrossRef]
- Nemec, A.; Krizova, L.; Maixnerova, M.; Sedo, O.; Brisse, S.; Higgins, P.G. Acinetobacter seifertii sp. nov., a member of the Acinetobacter calcoaceticus–Acinetobacter baumannii complex isolated from human clinical specimens. Int. J. Syst. Evol. Microbiol. 2015, 65, 934–942. [Google Scholar] [CrossRef] [Green Version]
- Cosgaya, C.; Marí-Almirall, A.; Van Assche, A.; Fernández-Orth, D.; Mosqueda, N.; Telli, M.; Huys, G.; Higgins, P.G.; Seifert, H.; Lievens, B.; et al. Acinetobacter dijkshoorniae sp. nov., a new member of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex mainly recovered from clinical samples in different countries. Int. J. Syst. Evol. Microbiol. 2016, 66, 4105–4111. [Google Scholar] [CrossRef]
- Rooney, A.; Dunlap, C.; Flor-Weiler, L.B. Acinetobacter lactucae sp. nov., isolated from iceberg lettuce (Asteraceae: Lactuca sativa). Int. J. Syst. Evol. Microbiol. 2016, 66, 3566–3572. [Google Scholar] [CrossRef] [PubMed]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Genet. 2018, 16, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Boo, T.W.; Walsh, F.; Crowley, B. Molecular characterization of carbapenem-resistant Acinetobacter species in an Irish university hospital: Predominance of Acinetobacter genomic species 3. J. Med. Microbiol. 2009, 58, 209–216. [Google Scholar] [CrossRef]
- Zhang, H.Z.; Zhang, J.S.; Qiao, L. The Acinetobacter baumannii group: A systemic review. World J. Emerg. Med. 2013, 4, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.C.; Huang, Y.T.; Tan, C.K.; Kuo, Y.W.; Liao, C.H.; Lee, P.I.; Hsueh, P.-R. Acinetobacter baumannii and Acinetobacter genospecies 13TU and 3 bacteraemia: Comparison of clinical features, prognostic factors and outcomes. J. Antimicrob. Chemother. 2011, 66, 1839–1846. [Google Scholar] [CrossRef]
- Kang, Y.S.; Jung, J.; Jeon, C.O.; Park, W. Acinetobacter oleivorans sp. nov. is capable of adhering to and growing on diesel-oil. J. Microbiol. 2011, 49, 29–34. [Google Scholar] [CrossRef]
- Hare, J.M.; Bradley, J.A.; Lin, C.L.; Elam, T.J. Diverse responses to UV light exposure in Acinetobacter include the capacity for DNA damage-induced mutagenesis in the opportunistic pathogens Acinetobacter baumannii and Acinetobacter ursingii. Microbiology 2012, 158, 601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateo-Estrada, V.; Grana-Miraglia, L.; Lopez-Leal, G.; Castillo-Ramirez, S. Phylogenomics reveals clear cases of misclassification and genus-wide phylogenetic markers for Acinetobacter. Genome Biol. Evol. 2019, 11, 2531–2541. [Google Scholar] [CrossRef] [PubMed]
- Beijerinck, M. Pigmenten als oxydatieproducten gevormd door bacterien. Versl K. Akad Wetensch Amst. 1911, 19, 1092–1103. [Google Scholar]
- Al Atrouni, A.; Joly-Guillou, M.-L.; Hamze, M.; Kempf, M. Reservoirs of non-baumannii Acinetobacter Species. Front. Microbiol. 2016, 7, 49. [Google Scholar] [CrossRef] [Green Version]
- Seifert, H.; Dijkshoorn, L.; Gerner-Smidt, P.; Pelzer, N.; Tjernberg, I.; Vaneechoutte, M. Distribution of Acinetobacter species on human skin: Comparison of phenotypic and genotypic identification methods. J. Clin. Microbiol. 1997, 35, 2819–2825. [Google Scholar] [CrossRef] [Green Version]
- La Scola, B.; Raoult, D. Acinetobacter baumannii in human body louse. Emerg. Infect. Dis. 2004, 10, 1671. [Google Scholar] [CrossRef]
- Kempf, M.; Abdissa, A.; Diatta, G.; Trape, J.-F.; Angelakis, E.; Mediannikov, O.; La Scola, B.; Raoult, D. Detection of Acinetobacter baumannii in human head and body lice from Ethiopia and identification of new genotypes. Int. J. Infect. Dis. 2012, 16, e680–e683. [Google Scholar] [CrossRef] [Green Version]
- Bouvresse, S.; Socolovschi, C.; Berdjane, Z.; Durand, R.; Izri, A.; Raoult, D.; Chosidow, O.; Brouqui, P. No evidence of Bartonella quintana but detection of Acinetobacter baumannii in head lice from elementary schoolchildren in Paris. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 475–477. [Google Scholar] [CrossRef]
- Fournier, P.E.; Vallenet, D.; Barbe, V.; Audic, S.; Ogata, H.; Poirel, L.; Richet, H.; Robert, C.; Mangenot, S.; Abergel, C.; et al. Comparative genomics of multidrug resistance in Acinetobacter baumannii. PLoS Genet. 2006, 2, e7. [Google Scholar] [CrossRef] [Green Version]
- Eveillard, M.; Kempf, M.; Belmonte, O.; Pailhories, H.; Joly-Guillou, M.L. Reservoirs of Acinetobacter baumannii out-side the hospital and potential involvement in emerging human community-acquired infections. Int. J. Infect. Dis. 2013, 17, e802–e805. [Google Scholar] [CrossRef] [Green Version]
- Byrne-Bailey, K.G.; Gaze, W.H.; Kay, P.; Boxall, A.B.; Hawkey, P.M.; Wellington, E.M. Prevalence of sulfonamide resistance genes in bacterial isolates from manured agricultural soils and pig slurry in the United Kingdom. Antimicrob. Agents Chemother. 2009, 53, 696–702. [Google Scholar] [CrossRef] [Green Version]
- Furlan, J.; Pitondo-Silva, A.; Stehling, E.G. New STs in multidrug-resistant Acinetobacter baumannii harbouring β -lactamases encoding genes isolated from Brazilian soils. J. Appl. Microbiol. 2018, 125, 506–512. [Google Scholar] [CrossRef]
- Girlich, D.; Poirel, L.; Nordmann, P. First isolation of the blaOXA-23 carbapenemase gene from an environmental Acinetobacter baumannii isolate. Antimicrob. Agents Chemother. 2009, 54, 578–579. [Google Scholar] [CrossRef] [Green Version]
- Higgins, P.G.; Hrenovic, J.; Seifert, H.; Dekic, S. Characterization of Acinetobacter baumannii from water and sludge line of secondary wastewater treatment plant. Water Res. 2018, 140, 261–267. [Google Scholar] [CrossRef]
- Kittinger, C.; Kirschner, A.; Lipp, M.; Baumert, R.; Mascher, F.; Farnleitner, A.H.; Zarfel, G.E. Antibiotic resistance of Acinetobacter spp. isolates from the river Danube: Susceptibility stays high. Int. J. Environ. Res. Public Health 2017, 15, 52. [Google Scholar] [CrossRef] [Green Version]
- Rafei, R.; Hamze, M.; Pailhoriès, H.; Eveillard, M.; Marsollier, L.; Joly-Guillou, M.-L.; Dabboussi, F.; Kempf, M. Extrahuman epidemiology of Acinetobacter baumannii in lebanon. Appl. Environ. Microbiol. 2015, 81, 2359–2367. [Google Scholar] [CrossRef] [Green Version]
- Houang, E.T.; Chu, Y.; Leung, C.; Chu, K.; Berlau, J.; Ng, K.; Cheng, A. Epidemiology and infection control implications of Acinetobacter spp. in Hong Kong. J. Clin. Microbiol. 2001, 39, 228–234. [Google Scholar] [CrossRef] [Green Version]
- Hrenovic, J.; Durn, G.; Goic-Barisic, I.; Kovacic, A. Occurrence of an environmental Acinetobacter baumannii strain similar to a clinical isolate in paleosol from Croatia. Appl. Environ. Microbiol. 2014, 80, 2860–2866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klotz, P.; Higgins, P.G.; Schaubmar, A.R.; Failing, K.; Leidner, U.; Seifert, H.; Scheufen, S.; Semmler, T.; Ewers, C. Seasonal occurrence and carbapenem susceptibility of bovine Acinetobacter baumannii in Germany. Front. Microbiol. 2019, 10, 272. [Google Scholar] [CrossRef] [Green Version]
- Al Bayssari, C.; Dabboussi, F.; Hamze, M.; Rolain, J.-M. Emergence of carbapenemase-producing Pseudomonas aeruginosa and Acinetobacter baumannii in livestock animals in Lebanon. J. Antimicrob. Chemother. 2015, 70, 950–951. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Berçot, B.; Millemann, Y.; Bonnin, R.A.; Pannaux, G.; Nordmann, P. Carbapenemase-producing Acinetobacter spp. in cattle, France. Emerg. Infect. Dis. 2012, 18, 523. [Google Scholar] [CrossRef]
- Wilharm, G.; Skiebe, E.; Higgins, P.G.; Poppel, M.T.; Blaschke, U.; Leser, S.; Heider, C.; Heindorf, M.; Brauner, P.; Jäckel, U.; et al. Relatedness of wildlife and livestock avian isolates of the nosocomial pathogen Acinetobacter baumannii to lineages spread in hospitals worldwide. Environ. Microbiol. 2017, 19, 4349–4364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamouda, A.; Findlay, J.; Al Hassan, L.; Amyes, S.G. Epidemiology of Acinetobacter baumannii of animal origin. Int. J. Antimicrob. Agents 2011, 38, 314–318. [Google Scholar] [CrossRef]
- Łopińska, A.; Indykiewicz, P.; Skiebe, E.; Pfeifer, Y.; Trček, J.; Jerzak, L.; Minias, P.; Nowakowski, J.; Ledwoń, M.; Betleja, J.; et al. Low occurrence of Acinetobacter baumannii in gulls and songbirds. Pol. J. Microbiol. 2020, 69, 85–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Sobouti, B.; Mirshekar, M.; Fallah, S.; Tabaei, A.; Mehrabadi, J.F.; Darbandi, A. Pan drug-resistant Acinetobacter baumannii causing nosocomial infections among burnt children. Med. J. Islam. Repub. Iran. 2020, 34, 24. [Google Scholar] [PubMed]
- Goic-Barisic, I.; Music, M.S.; Kovacic, A.; Tonkic, M.; Hrenovic, J. Pan drug-resistant environmental isolate of Acinetobacter baumannii from Croatia. Microb. Drug Resist. 2017, 23, 494–496. [Google Scholar] [CrossRef] [Green Version]
- Quinones, D.; Carvajal, I.; Perez, Y.; Hart, M.; Perez, J.; Garcia, S.; Salazar, D.; Ghosh, S.; Kawaguchiya, M.; Aung, M.S.; et al. High prevalence of bla OXA-23 in Acinetobacter spp. and detection of bla NDM-1 in A. soli in Cuba: Report from national surveillance program (2010–2012). New Microbes New Infect. 2015, 7, 52–56. [Google Scholar] [CrossRef] [Green Version]
- Boluki, E.; Kazemian, H.; Peeridogaheh, H.; Alikhani, M.Y.; Shahabi, S.; Beytollahi, L.; Ghorbanzadeh, R. Antimicrobial activity of photodynamic therapy in combination with colistin against a pan-drug resistant Acinetobacter baumannii isolated from burn patient. Photodiagn. Photodyn. Ther. 2017, 18, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Al-Kadmy, I.M.S.; Ibrahim, S.A.; Al-Saryi, N.; Aziz, S.N.; Besinis, A.; Hetta, H.F. Prevalence of genes involved in colistin resistance in Acinetobacter baumannii: First report from Iraq. Microb. Drug Resist. 2020, 26, 616–622. [Google Scholar] [CrossRef]
- Nowak, J.; Zander, E.; Stefanik, D.; Higgins, P.G.; Roca, I.; Vila, J.; McConnell, M.J.; Cisneros, J.M.; Seifert, H. High incidence of pandrug-resistant Acinetobacter baumannii isolates collected from patients with ventilator-associated pneumonia in Greece, Italy and Spain as part of the MagicBullet clinical trial. J. Antimicrob. Chemother. 2017, 72, 3277–3282. [Google Scholar] [CrossRef]
- Falagas, M.E.; Bliziotis, I.A.; Siempos, I.I. Attributable mortality of Acinetobacter baumannii infections in critically ill patients: A systematic review of matched cohort and case-control studies. Crit. Care 2006, 10, R48. [Google Scholar] [CrossRef] [Green Version]
- Zhanel, G.G.; Clark, N.M.; Lynch, J.P. Infections Due to Acinetobacter baumannii in the ICU: Treatment options. Semin. Respir. Crit. Care Med. 2017, 38, 311–325. [Google Scholar] [CrossRef] [Green Version]
- Scott, P.; Deye, G.; Srinivasan, A.; Murray, C.; Moran, K.; Hulten, E.; Fishbain, J.; Craft, D.; Riddell, S.; Lindler, L.; et al. An outbreak of multidrug-resistant Acinetobacter baumannii-calcoaceticus complex infection in the US military health care system associated with military operations in Iraq. Clin. Infect. Dis. 2007, 44, 1577–1584. [Google Scholar] [CrossRef] [Green Version]
- Öncül, O.; Keskin, Ö.; Acar, H.; Küçükardalı, Y.; Evrenkaya, R.; Atasoyu, E.; Top, C.; Nalbant, S.; Özkan, S.; Emekdaş, G.; et al. Hospital-acquired infections following the 1999 Marmara earthquake. J. Hosp. Infect. 2002, 51, 47–51. [Google Scholar] [CrossRef]
- Rafei, R.; Dabboussi, F.; Hamze, M.; Eveillard, M.; Lemarie, C.; Mallat, H.; Rolain, J.-M.; Joly-Guillou, M.-L.; Kempf, M. First report of blaNDM-1-producing Acinetobacter baumannii isolated in Lebanon from civilians wounded during the Syrian war. Int. J. Infect. Dis. 2014, 21, 21–23. [Google Scholar] [CrossRef] [Green Version]
- Tao, C.; Kang, M.; Chen, Z.; Xie, Y.; Fan, H.; Qin, L.; Ma, Y. Microbiologic study of the pathogens isolated from wound culture among Wenchuan earthquake survivors. Diagn. Microbiol. Infect. Dis. 2009, 63, 268–270. [Google Scholar] [CrossRef]
- Lai, C.-C.; Wang, C.-Y.; Hsueh, P.-R. Co-infections among patients with COVID-19: The need for combination therapy with non-anti-SARS-CoV-2 agents? J. Microbiol. Immunol. Infect. 2020, 53, 505–512. [Google Scholar] [CrossRef]
- Sharifipour, E.; Shams, S.; Esmkhani, M.; Khodadadi, J.; Fotouhi-Ardakani, R.; Koohpaei, A.; Doosti, Z.; Golzari, S.E. Evaluation of bacterial co-infections of the respiratory tract in COVID-19 patients admitted to ICU. BMC Infect. Dis. 2020, 20, 1–7. [Google Scholar] [CrossRef]
- Perez, S.; Innes, G.K.; Walters, M.S.; Mehr, J.; Arias, J.; Greeley, R.; Chew, D. Increase in hospital-acquired carbapenem-resistant Acinetobacter baumannii infection and colonization in an acute care hospital during a surge in COVID-19 admissions—New Jersey, February–July 2020. Morb. Mortal. Wkly. Rep. 2020, 69, 1827. [Google Scholar] [CrossRef]
- Chen, X.; Liao, B.; Cheng, L.; Peng, X.; Xu, X.; Li, Y.; Hu, T.; Li, J.; Zhou, X.; Ren, B. The microbial coinfection in COVID-19. Appl. Microbiol. Biotechnol. 2020, 104, 1–9. [Google Scholar] [CrossRef]
- Anstey, N.M.; Currie, B.J.; Hassell, M.; Palmer, D.; Dwyer, B.; Seifert, H. Community-Acquired Bacteremic Acinetobacter Pneumonia in tropical Australia is caused by diverse strains of Acinetobacter baumannii, with carriage in the throat in at-risk groups. J. Clin. Microbiol. 2002, 40, 685–686. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.-Z.; Hsueh, P.-R.; Lee, L.-N.; Yu, C.-J.; Yang, P.-C.; Luh, K.-T. Severe community-acquired pneumonia due to Acinetobacter baumannii. Chest 2001, 120, 1072–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, W.-S.; Chu, C.-M.; Tsang, K.-Y.; Lo, F.-H.; Lo, K.-F.; Ho, P.-L. Fulminant community-acquired Acinetobacter baumannii pneumonia as a distinct clinical syndrome. Chest 2006, 129, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Ong, C.W.; Lye, D.C.; Khoo, K.L.; Chua, G.S.W.; Yeoh, S.F.; Leo, Y.S.; Tambyah, P.A.; Chua, A.C. Severe community-acquired Acinetobacter baumannii pneumonia: An emerging highly lethal infectious disease in the Asia–Pacific. Respirology 2009, 14, 1200–1205. [Google Scholar] [CrossRef]
- Dexter, C.; Murray, G.L.; Paulsen, I.T.; Peleg, A.Y. Community-acquired Acinetobacter baumannii: Clinical characteris-tics, epidemiology and pathogenesis. Expert Rev. Anti-Infect. Ther. 2015, 13, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, D.N.; Elbourne, L.D.H.; Hassan, K.A.; Eijkelkamp, B.A.; Tetu, S.G.; Brown, M.H.; Shah, B.S.; Peleg, A.; Mabbutt, B.C.; Paulsen, I.T. The complete genome and phenome of a community-acquired Acinetobacter baumannii. PLoS ONE 2013, 8, e58628. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wu, X.; Tang, J.; Zhang, L.; Jia, X. Trends and development in the antibiotic-resistance of Acinetobacter baumannii: A scientometric research study (1991–2019). Infect. Drug Resist. 2020, 13, 3195–3208. [Google Scholar] [CrossRef] [PubMed]
- Central Asian and European Surveillance of Antimicrobial Resistance Annual Report 2019; WHO Regional Office for Europe: Geneva, Switzerland, 2019.
- Surveillance of Antimicrobial Resistance in Europe 2018: Annual Report of the European Antimicrobial Resistance Surveillance Network (EARS-Net); European Centre for Disease Prevention and Control: Solna, Sweden, 2019.
- Lob, S.H.; Hoban, D.J.; Sahm, D.F.; Badal, R.E. Regional differences and trends in antimicrobial susceptibility of Acinetobacter baumannii. Int. J. Antimicrob. Agents 2016, 47, 317–323. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Acinetobacter baumannii infections among patients at military medical facilities treating injured US service members, 2002–2004. MMWR Morb. Mortal. Wkly. Rep. 2004, 53, 1063.
- Moradi, J.; Hashemi, F.B.; Bahador, A. Antibiotic resistance of Acinetobacter baumannii in Iran: A systemic review of the published literature. Osong Public Health Res. Perspect. 2015, 6, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Bolourchi, N.; Azizi, O.; Tabrizi, A.M.A.; Esmaeili, S.; Fereshteh, S.; Badmasti, F. Clonal relatedness of Acinetobacter baumannii isolated from the middle east: A systematic review. Rev. Med. Microbiol. 2020. [Google Scholar] [CrossRef]
- Griffith, M.E.; Ceremuga, J.M.; Ellis, M.W.; Guymon, C.H.; Hospenthal, D.R.; Murray, C.K. Acinetobacter skin colonization of US army soldiers. Infect. Control Hosp. Epidemiol. 2006, 27, 659–661. [Google Scholar] [CrossRef]
- Griffith, M.E.; Lazarus, D.R.; Mann, P.B.; Boger, J.A.; Hospenthal, D.R.; Murray, C.K. Acinetobacter skin carriage among US army soldiers deployed in Iraq. Infect. Control Hosp. Epidemiol. 2007, 28, 720–722. [Google Scholar] [CrossRef] [PubMed]
- Keen, E.F., 3rd; Robinson, B.J.; Hospenthal, D.R.; Aldous, W.K.; Wolf, S.E.; Chung, K.K.; Murray, C.K. Prevalence of multidrug-resistant organisms recovered at a military burn center. Burns 2010, 36, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Echols, R.; Magee, G.; Ferreira, J.C.A.; Morgan, G.; Ariyasu, M.; Sawada, T.; Nagata, T.D. Prevalence of carbapenem-resistant gram-negative infections in the United States predominated by Acinetobacter baumannii and Pseudomonas aeruginosa. Open Forum Infect. Dis. 2017, 4, ofx176. [Google Scholar] [CrossRef] [PubMed]
- Manikal, V.M.; Landman, D.; Saurina, G.; Oydna, E.; Lal, H.; Quale, J. Endemic carbapenem-resistant Acinetobacter species in Brooklyn, New York: Citywide prevalence, interinstitutional spread, and relation to antibiotic usage. Clin. Infect. Dis. 2000, 31, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Yang, J.; Hu, R.; Hu, X.; Chen, K. Acinetobacter baumannii infection and IL-17 mediated immunity. Mediat. Inflamm. 2016, 2016, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.H.; Lee, J.S.; Lee, Y.C.; Park, T.I.; Lee, J.C. Acinetobacter baumannii invades epithelial cells and outer mem-brane protein A mediates interactions with epithelial cells. BMC Microbiol. 2008, 8, 216. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.H.; Hyun, S.H.; Lee, J.Y.; Lee, J.S.; Lee, Y.S.; Kim, S.A.; Chae, J.P.; Yoo, S.M.; Lee, J.C. Acinetobacter baumannii outer membrane protein A targets the nucleus and induces cytotoxicity. Cell. Microbiol. 2008, 10, 309–319. [Google Scholar] [CrossRef]
- Choi, C.H.; Lee, E.Y.; Lee, Y.C.; Park, T.I.; Kim, H.J.; Hyun, S.H.; Kim, S.A.; Lee, S.-K.; Lee, J.C. Outer membrane protein 38 of Acinetobacter baumannii localizes to the mitochondria and induces apoptosis of epithelial cells. Cell. Microbiol. 2005, 7, 1127–1138. [Google Scholar] [CrossRef]
- Choi, J.; Park, Y.; Kim, C.; Park, Y.; Yoon, H.; Shin, S.; Kim, Y.A.; Song, Y.; Yong, D.; Lee, K. Mortality risk factors of Acinetobacter baumannii bacteraemia. Intern. Med. J. 2005, 35, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Smani, Y.; Dominguez-Herrera, J.; Pachón, J. Association of the outer membrane protein Omp33 with fitness and virulence of Acinetobacter baumannii. J. Infect. Dis. 2013, 208, 1561–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rumbo, C.; Tomás, M.; Moreira, E.F.; Soares, N.C.; Carvajal, M.; Santillana, E.; Beceiro, A.; Romero, A.; Bou, G. The Acinetobacter baumannii Omp33–36 porin is a virulence factor that induces apoptosis and modulates autophagy in human cells. Infect. Immun. 2014, 82, 4666–4680. [Google Scholar] [CrossRef] [Green Version]
- Brossard, K.A.; Campagnari, A.A. The Acinetobacter baumannii biofilm-associated protein plays a role in adherence to human epithelial cells. Infect. Immun. 2011, 80, 228–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, A.; Merino, M.; Rumbo-Feal, S.; Alvarez-Fraga, L.; Vallejo, J.A.; Beceiro, A.; Ohneck, E.J.; Mateos, J.; Fernandez-Puente, P.; Actis, L.A.; et al. The FhaB/FhaC two-partner secretion system is involved in adhesion of Acinetobacter baumannii AbH12O-A2 strain. Virulence 2017, 8, 959–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weidensdorfer, M.; Chae, J.I.; Makobe, C.; Stahl, J.; Averhoff, B.; Muller, V.; Schurmann, C.; Brandes, R.P.; Wilharm, G.; Ballhorn, W.; et al. Analysis of endothelial adherence of Bartonella henselae and Acinetobacter baumannii using a dynamic human ex vivo infection model. Infect. Immun. 2015, 84, 711–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentancor, L.V.; Camacho-Peiro, A.; Bozkurt-Guzel, C.; Pier, G.B.; Maira-Litran, T. Identification of Ata, a multifunctional trimeric autotransporter of Acinetobacter baumannii. J. Bacteriol. 2012, 194, 3950–3960. [Google Scholar] [CrossRef] [Green Version]
- Weidensdorfer, M.; Ishikawa, M.; Hori, K.; Linke, D.; Djahanschiri, B.; Iruegas, R.; Ebersberger, I.; Riedel-Christ, S.; Enders, G.; Leukert, L.; et al. The Acinetobacter trimeric autotransporter adhesin Ata controls key virulence traits of Acinetobacter baumannii. Virulence 2019, 10, 68–81. [Google Scholar] [CrossRef] [Green Version]
- Liou, M.-L.; Soo, P.-C.; Ling, S.-R.; Kuo, H.-Y.; Tang, C.Y.; Chang, K.-C. The sensor kinase BfmS mediates virulence in Acinetobacter baumannii. J. Microbiol. Immunol. Infect. 2014, 47, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Tomaras, A.P.; Flagler, M.J.; Dorsey, C.W.; Gaddy, J.A.; Actis, L.A. Characterization of a two-component regulatory system from Acinetobacter baumannii that controls biofilm formation and cellular morphology. Microbiology 2008, 154, 3398–3409. [Google Scholar] [CrossRef] [Green Version]
- Wood, C.R.; Ohneck, E.J.; Edelmann, R.E.; Actis, L.A. A light-regulated type I pilus contributes to Acinetobacter baumannii biofilm, motility, and virulence functions. Infect. Immun. 2018, 86, e00442-18. [Google Scholar] [CrossRef] [Green Version]
- Eijkelkamp, B.A.; Stroeher, U.H.; Hassan, K.A.; Papadimitrious, M.S.; Paulsen, I.T.; Brown, M.H. Adherence and motility characteristics of clinical Acinetobacter baumannii isolates. FEMS Microbiol. Lett. 2011, 323, 44–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eijkelkamp, B.A.; Stroeher, U.H.; Hassan, K.A.; Elbourne, L.D.; Paulsen, I.T.; Brown, M.H. H-NS plays a role in expression of Acinetobacter baumannii virulence features. Infect. Immun. 2013, 81, 2574–2583. [Google Scholar] [CrossRef] [Green Version]
- de Breij, A.; Gaddy, J.; van der Meer, J.; Koning, R.; Koster, A.; van den Broek, P.; Actis, L.; Nibbering, P.; Dijkshoorn, L. CsuA/BABCDE-dependent pili are not involved in the adherence of Acinetobacter baumannii ATCC19606(T) to human airway epithelial cells and their inflammatory response. Res. Microbiol. 2009, 160, 213–218. [Google Scholar] [CrossRef] [PubMed]
- May, H.C.; Yu, J.-J.; Zhang, H.; Wang, Y.; Cap, A.P.; Chambers, J.P.; Guentzel, M.N.; Arulanandam, B.P. Thioredoxin-A is a virulence factor and mediator of the type IV pilus system in Acinetobacter baumannii. PLoS ONE 2019, 14, e0218505. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.C.; Hood, I.; Boyd, K.L.; Olson, P.D.; Morrison, J.M.; Carson, S.; Sayood, K.; Iwen, P.C.; Skaar, E.P.; Dunman, P.M. Inactivation of phospholipase D diminishes Acinetobacter baumannii pathogenesis. Infect. Immun. 2010, 78, 1952–1962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiester, S.E.; Arivett, B.A.; Schmidt, R.E.; Beckett, A.C.; Ticak, T.; Carrier, M.V.; Ghosh, R.; Ohneck, E.J.; Metz, M.L.; Jeffries, M.; et al. Iron-regulated phospholipase C activity contributes to the cytolytic activity and virulence of Acinetobacter baumannii. PLoS ONE 2016, 11, e0167068. [Google Scholar] [CrossRef] [Green Version]
- Stahl, J.; Bergmann, H.; Göttig, S.; Ebersberger, I.; Averhoff, B. Acinetobacter baumannii virulence is mediated by the concerted action of three phospholipases D. PLoS ONE 2015, 10, e0138360. [Google Scholar] [CrossRef]
- Smani, Y.; Docobo-Pérez, F.; López-Rojas, R.; Domínguez-Herrera, J.; Ibáñez-Martínez, J.; Pachón, J. Platelet-activating factor receptor initiates contact of Acinetobacter baumannii expressing phosphorylcholine with host cells. J. Biol. Chem. 2012, 287, 26901–26910. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.M.; Azzouz, M.; Häggendal, B.; Säljö, K.; Malmi, H.; Zavialov, A.; Teneberg, S. Glycosphingolipids recognized by Acinetobacter baumannii. Microorganisms 2020, 8, 612. [Google Scholar] [CrossRef]
- Lee, C.R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, antibiotic resistance mechanisms, and prospective treatment options. Front. Cell Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef] [Green Version]
- Erridge, C.; Moncayo-Nieto, O.L.; Morgan, R.; Young, M.; Poxton, I.R. Acinetobacter baumannii lipopolysaccharides are potent stimulators of human monocyte activation via Toll-like receptor 4 signalling. J. Med. Microbiol. 2007, 56, 165–171. [Google Scholar] [CrossRef]
- Luke, N.R.; Sauberan, S.L.; Russo, T.A.; Beanan, J.M.; Olson, R.; Loehfelm, T.W.; Cox, A.D.; Michael, F.S.; Vinogradov, E.V.; Campagnari, A.A. Identification and characterization of a glycosyltransferase involved in Acinetobacter baumannii lipopolysaccharide core biosynthesis. Infect. Immun. 2010, 78, 2017–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Tan, B.; Pantapalangkoor, P.; Ho, T.; Baquir, B.; Tomaras, A.; Montgomery, J.I.; Barbacci, E.G.; Hujer, K.; Bonomo, R.A. Inhibition of LpxC protects mice from resistant Acinetobacter baumannii by modulating inflammation and enhancing phagocytosis. MBio 2012, 3, e00312-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McQueary, C.N.; Kirkup, B.C.; Si, Y.; Barlow, M.; Actis, L.A.; Craft, D.W.; Zurawski, D.V. Extracellular stress and lipopolysaccharide modulate Acinetobacter baumannii surface-associated motility. J. Microbiol. 2012, 50, 434–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McConnell, M.J.; Actis, L.A.; Pachón, J. Acinetobacter baumannii: Human infections, factors contributing to pathogenesis and animal models. FEMS Microbiol. Rev. 2013, 37, 130–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.-W.; Lee, S.-H.; Lee, D.-H. Relationship of serum gamma-glutamyltransferase levels with pulmonary function and chronic obstructive pulmonary disease. Lung 2014, 192, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Ermis, H.; Celik, M.R.; Gulbas, G.; Tavli, D.; Aytemur, Z.A. Relationship between serum γ-glutamyltransferase levels and acute exacerbation of chronic obstructive pulmonary disease. Pol. Arch. Intern. Med. 2013, 123, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Elhossieny, N.M.; Elhezawy, N.B.; Sayed, R.M.; Khattab, M.S.; el Far, M.Y.; Attia, A.S. Gamma-glutamyl transferase is a novel virulence factor of Acinetobacter baumannii that induces alveolar wall destruction and renal damage in systemic pathology. J. Infect. Dis. 2020, 222, 871–879. [Google Scholar] [CrossRef]
- Carruthers, M.D.; Nicholson, P.A.; Tracy, E.N.; Munson, R.S., Jr. Acinetobacter baumannii utilizes a type VI secretion system for bacterial competition. PLoS ONE 2013, 8, e59388. [Google Scholar] [CrossRef]
- Bannister, B.A.; Begg, N.T.; Gillespie, S.H. Infectious Disease, 2nd ed.; Blackwell Science: Hoboken, NJ, USA, 2000. [Google Scholar]
- Wyres, K.L.; Cahill, S.M.; Holt, K.E.; Hall, R.M.; Kenyon, J.J. Identification of Acinetobacter baumannii loci for capsular polysaccharide (KL) and lipooligosaccharide outer core (OCL) synthesis in genome assemblies using curated reference databases compatible with Kaptive. Microb. Genom. 2020, 6, e000339. [Google Scholar] [CrossRef]
- Russo, T.A.; Luke, N.R.; Beanan, J.M.; Olson, R.; Sauberan, S.L.; Macdonald, U.; Schultz, L.W.; Umland, T.C.; Campagnari, A.A. The K1 capsular polysaccharide of Acinetobacter baumannii Strain 307-0294 is a major virulence factor. Infect. Immun. 2010, 78, 3993–4000. [Google Scholar] [CrossRef] [Green Version]
- Iwashkiw, J.A.; Seper, A.; Weber, B.S.; Scott, N.E.; Vinogradov, E.; Stratilo, C.; Reiz, B.; Cordwell, S.J.; Whittal, R.; Schild, S. Identification of a general O-linked protein glycosylation system in Acinetobacter baumannii and its role in virulence and biofilm formation. PLoS Pathog. 2012, 8, e1002758. [Google Scholar] [CrossRef]
- Lees-Miller, R.G.; Iwashkiw, J.A.; Scott, N.E.; Seper, A.; Vinogradov, E.; Schild, S.; Feldman, M.F. A common pathway for O-linked protein-glycosylation and synthesis of capsule in Acinetobacter baumannii. Mol. Microbiol. 2013, 89, 816–830. [Google Scholar] [CrossRef]
- Zimbler, D.L.; Park, T.M.; Arivett, B.A.; Penwell, W.F.; Greer, S.M.; Woodruff, T.M.; Tierney, D.L.; Actis, L.A. Stress response and virulence functions of the Acinetobacter baumannii NfuA FeS scaffold protein. J. Bacteriol. 2012, 194, 2884–2893. [Google Scholar] [CrossRef] [Green Version]
- Juttukonda, L.J.; Chazin, W.J.; Skaar, E.P. Acinetobacter baumannii coordinates urea metabolism with metal import to resist host-mediated metal limitation. mBio 2016, 7, e01475-16. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, B.L.; Skaar, E.P. The contribution of nutrient metal acquisition and metabolism to Acinetobacter baumannii survival within the host. Front. Cell. Infect. Microbiol. 2013, 3, 95. [Google Scholar] [CrossRef]
- Nairn, B.L.; Lonergan, Z.; Wang, J.; Braymer, J.; Zhang, Y.; Calcutt, M.W.; Lisher, J.P.; Gilston, B.A.; Chazin, W.J.; De Crécy-Lagard, V.; et al. The response of Acinetobacter baumannii to zinc starvation. Cell Host Microbe 2016, 19, 826–836. [Google Scholar] [CrossRef] [Green Version]
- Koenigs, A.; Stahl, J.; Averhoff, B.; Göttig, S.; Wichelhaus, T.A.; Wallich, R.; Zipfel, P.F.; Kraiczy, P. CipA of Acinetobacter baumannii is a novel plasminogen binding and complement inhibitory protein. J. Infect. Dis. 2016, 213, 1388–1399. [Google Scholar] [CrossRef] [Green Version]
- Elhosseiny, N.M.; Amin, M.A.; Yassin, A.S.; Attia, A.S. Acinetobacter baumannii universal stress protein A plays a pivotal role in stress response and is essential for pneumonia and sepsis pathogenesis. Int. J. Med Microbiol. 2015, 305, 114–123. [Google Scholar] [CrossRef]
- Gebhardt, M.J.; Gallagher, L.A.; Jacobson, R.K.; Usacheva, E.A.; Peterson, L.R.; Zurawski, D.V.; Shuman, H.A. Joint transcritional control of virulence and resistance to antibiotic and environmental stress in Acinetobacter baumannii. MBio 2015, 6, e01660-15. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, A.; McClean, S. The role of universal stress proteins in bacterial infections. Curr. Med. Chem. 2017, 24, 3970–3979. [Google Scholar] [CrossRef]
- Sun, D.; Crowell, S.A.; Harding, C.M.; De Silva, P.M.; Harrison, A.; Fernando, D.M.; Mason, K.M.; Santana, E.; Loewen, P.C.; Kumar, A.; et al. KatG and KatE confer Acinetobacter resistance to hydrogen peroxide but sensitize bacteria to killing by phagocytic respiratory burst. Life Sci. 2016, 148, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, G.L.; Peleg, A.; Doi, Y. Acinetobacter baumannii: Evolution of Antimicrobial Resistance—Treatment Options. Semin. Respir. Crit. Care Med. 2015, 36, 085–098. [Google Scholar] [CrossRef] [Green Version]
- Gaddy, J.A.; Arivett, B.A.; McConnell, M.J.; Lopez-Rojas, R.; Pachon, J.; Actis, L.A. Role of acinetobactin-mediated iron acquisition functions in the interaction of Acinetobacter baumannii strain ATCC 19606T with human lung epithelial cells, Galleria mellonella caterpillars, and mice. Infect. Immun. 2012, 80, 1015–1024. [Google Scholar] [CrossRef] [Green Version]
- De Gregorio, E.; Del Franco, M.; Martinucci, M.; Roscetto, E.; Zarrilli, R.; Di Nocera, P.P. Biofilm-associated proteins: News from Acinetobacter. BMC Genom. 2015, 16, 933. [Google Scholar] [CrossRef] [Green Version]
- Roussin, M.; Rabarioelina, S.; Cluzeau, L.; Cayron, J.; Lesterlin, C.; Salcedo, S.P.; Bigot, S. Identification of a contact-dependent growth inhibition (CDI) system that reduces biofilm formation and host cell adhesion of Acinetobacter baumannii DSM30011 strain. Front. Microbiol. 2019, 10, 2450. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira, G.M.; Kostoulias, X.; Khoo, C.; Aibinu, I.; Qu, Y.; Traven, A.; Peleg, A. A global virulence regulator in Acinetobacter baumannii and its control of the phenylacetic acid catabolic pathway. J. Infect. Dis. 2014, 210, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Repizo, G.D.; Gagné, S.; Foucault-Grunenwald, M.-L.; Borges, V.; Charpentier, X.; Limansky, A.S.; Gomes, J.P.; Viale, A.M.; Salcedo, S.P. Differential role of the T6SS in Acinetobacter baumannii virulence. PLoS ONE 2015, 10, e0138265. [Google Scholar] [CrossRef] [Green Version]
- Seder, R.A.; Hill, A.V.S. Vaccines against intracellular infections requiring cellular immunity. Nat. Cell Biol. 2000, 406, 793–798. [Google Scholar] [CrossRef]
- McConnell, M.J.; Domínguez-Herrera, J.; Smani, Y.; López-Rojas, R.; Docobo-Pérez, F.; Pachón, J. Vaccination with outer membrane complexes elicits rapid protective immunity to multidrug-resistant Acinetobacter baumannii. Infect. Immun. 2010, 79, 518–526. [Google Scholar] [CrossRef] [Green Version]
- McConnell, M.J.; Pachón, J. Active and passive immunization against Acinetobacter baumannii using an inactivated whole cell vaccine. Vaccine 2010, 29, 1–5. [Google Scholar] [CrossRef]
- McConnell, M.J.; Rumbo, C.; Bou, G.; Pachón, J. Outer membrane vesicles as an acellular vaccine against Acinetobacter baumannii. Vaccine 2011, 29, 5705–5710. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, S.; Ketter, P.M.; Yu, J.-J.; Grimm, R.C.; May, H.C.; Cap, A.P.; Chambers, J.P.; Guentzel, M.N.; Arulanan-dam, B.P. Vaccination with a live attenuated Acinetobacter baumannii deficient in thioredoxin provides protection against systemic Acinetobacter infection. Vaccine 2017, 35, 3387–3394. [Google Scholar] [CrossRef]
- Shu, M.H.; MatRahim, N.; NorAmdan, N.; Pang, S.P.; Hashim, S.H.; Phoon, W.H.; AbuBakar, S. An Inactivated antibiotic exposed whole-cell vaccine enhances bactericidal activities against multidrug-resistant Acinetobacter baumannii. Sci. Rep. 2016, 6, 22332. [Google Scholar] [CrossRef] [Green Version]
- KuoLee, R.; Harris, G.; Yan, H.; Xu, H.H.; Conlan, W.J.; Patel, G.B.; Chen, W. Intranasal immunization protects against Acinetobacter baumannii-associated pneumonia in mice. Vaccine 2015, 33, 260–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Quintanilla, M.; Pulido, M.R.; Pachón, J.; McConnell, M.J. Immunization with lipopolysaccharide-deficient whole cells provides protective immunity in an experimental mouse model of Acinetobacter baumannii Infection. PLoS ONE 2014, 9, e114410. [Google Scholar] [CrossRef] [Green Version]
- Wizemann, T.M.; Adamou, J.E.; Langermann, S. Adhesins as targets for vaccine development. Emerg. Infect. Dis. 1999, 5, 395–403. [Google Scholar] [CrossRef]
- Jahangiri, A.; Rasooli, I.; Owlia, P.; Fooladi, A.A.I.; Salimian, J. In silico design of an immunogen against Acinetobacter baumannii based on a novel model for native structure of outer membrane protein A. Microb. Pathog. 2017, 105, 201–210. [Google Scholar] [CrossRef]
- Nie, D.; Hu, Y.; Chen, Z.; Li, M.; Hou, Z.; Luo, X.; Mao, X.; Xue, X. Outer membrane protein A (OmpA) as a potential therapeutic target for Acinetobacter baumannii infection. J. Biomed. Sci. 2020, 27, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yang, T.; Cao, J.; Sun, J.; Dai, W.; Zhang, L. Mucosal immunization with purified OmpA elicited protective immunity against infections caused by multidrug-resistant Acinetobacter baumannii. Microb. Pathog. 2016, 96, 20–25. [Google Scholar] [CrossRef]
- Luo, G.; Lin, L.; Ibrahim, A.S.; Baquir, B.; Pantapalangkoor, P.; Bonomo, R.A.; Doi, Y.; Adams, M.D.; Russo, T.A.; Spellberg, B. Active and Passive Immunization protects against lethal, extreme drug resistant-Acinetobacter baumannii infection. PLoS ONE 2012, 7, e29446. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Tan, B.; Pantapalangkoor, P.; Ho, T.; Hujer, A.M.; Taracila, M.A.; Bonomo, R.A.; Spellberg, B. Acinetobacter baumannii rOmpA vaccine dose alters immune polarization and immunodominant epitopes. Vaccine 2013, 31, 313–318. [Google Scholar] [CrossRef] [Green Version]
- Islam, A.H.M.S.; Singh, K.-K.B.; Ismail, A. Demonstration of an outer membrane protein that is antigenically specific for Acinetobacter baumannii. Diagn. Microbiol. Infect. Dis. 2011, 69, 38–44. [Google Scholar] [CrossRef]
- Jahangiri, A.; Rasooli, I.; Owlia, P.; Fooladi, A.A.I.; Salimian, J. Highly conserved exposed immunogenic peptides of Omp34 against Acinetobacter baumannii: An innovative approach. J. Microbiol. Methods 2018, 144, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Bentancor, L.V.; Routray, A.; Bozkurt-Guzel, C.; Camacho-Peiro, A.; Pier, G.B.; Maira-Litran, T. Evaluation of the trimeric autotransporter Ata as a vaccine candidate against Acinetobacter baumannii infections. Infect. Immun. 2012, 80, 3381–3388. [Google Scholar] [CrossRef] [Green Version]
- Bentancor, L.V.; O’Malley, J.M.; Bozkurt-Guzel, C.; Pier, G.B.; Maira-Litran, T. Poly-N-acetyl-beta-(1–6)-glucosamine is a target for protective immunity against Acinetobacter baumannii infections. Infect. Immun. 2012, 80, 651–656. [Google Scholar] [CrossRef] [Green Version]
- Russo, T.A.; Beanan, J.M.; Olson, R.; Macdonald, U.; Cox, A.D.; Michael, F.S.; Vinogradov, E.V.; Spellberg, B.; Luke-Marshall, N.R.; Campagnari, A.A. The K1 capsular polysaccharide from Acinetobacter baumannii is a potential therapeutic target via passive immunization. Infect. Immun. 2013, 81, 915–922. [Google Scholar] [CrossRef] [Green Version]
- Fattahian, Y.; Rasooli, I.; Gargari, S.L.M.; Rahbar, M.R.; Astaneh, S.D.; Amani, J. Protection against Acinetobacter baumannii infection via its functional deprivation of biofilm associated protein (Bap). Microb. Pathog. 2011, 51, 402–406. [Google Scholar] [CrossRef]
- Aghajani, Z.; Rasooli, I.; Gargari, S.L.M. Exploitation of two siderophore receptors, BauA and BfnH, for protection against Acinetobacter baumannii infection. Apmis 2019, 127, 753–763. [Google Scholar] [CrossRef]
- Huang, W.; Yao, Y.; Wang, S.; Xia, Y.; Yang, X.; Long, Q.; Sun, W.; Liu, C.; Li, Y.; Chu, X.; et al. Immunization with a 22-kDa outer membrane protein elicits protective immunity to multidrug-resistant Acinetobacter baumannii. Sci. Rep. 2016, 6, 20724. [Google Scholar] [CrossRef]
- Casey, W.T.; Spink, N.; Cia, F.; Collins, C.; Romano, M.; Berisio, R.; Bancroft, G.J.; McClean, S. Identification of an OmpW homologue in Burkholderia pseudomallei, a protective vaccine antigen against melioidosis. Vaccine 2016, 34, 2616–2621. [Google Scholar] [CrossRef]
- Singh, R.; Capalash, N.; Sharma, P. Immunoprotective potential of BamA, the outer membrane protein assembly factor, against MDR Acinetobacter baumannii. Sci. Rep. 2017, 7, 12411. [Google Scholar] [CrossRef] [PubMed]
- Tohidinia, M.; Moshtaghioun, S.M.; Sefid, F.; Falahati, A. Functional exposed amino acids of Caro analysis as a potential vaccine candidate in Acinetobacter baumannii. Int. J. Pept. Res. Ther. 2019, 26, 1185–1197. [Google Scholar] [CrossRef]
- Catel-Ferreira, M.; Marti, S.; Guillon, L.; Jara, L.; Coadou, G.; Molle, V.; Bouffartigues, E.; Bou, G.; Shalk, I.; Jouenne, T. The outer membrane porin OmpW of Acinetobacter baumannii is involved in iron uptake and colistin binding. FEBS Lett. 2016, 590, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Wang, S.; Yao, Y.; Xia, Y.; Yang, X.; Long, Q.; Sun, W.; Liu, C.; Li, Y.; Ma, Y. OmpW is a potential target for eliciting protective immunity against Acinetobacter baumannii infections. Vaccine 2015, 33, 4479–4485. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.J.; Ren, S.; Xie, Y.E. Evaluation of the protective efficacy of a fused OmpK/Omp22 protein vaccine candidate against Acinetobacter baumannii infection in mice. Biomed. Environ. Sci. 2018, 31, 155–158. [Google Scholar]
- Yang, A.; Yang, H.; Guo, S.; Xie, Y. MF59 adjuvant enhances the immunogenicity and protective immunity of the OmpK/Omp22 fusion protein from Acineterbacter baumannii through intratracheal inoculation in mice. Scand. J. Immunol. 2019, 90, e12769. [Google Scholar] [CrossRef]
- García-Patiño, M.G.; García-Contreras, R.; Licona-Limón, P. The immune response against Acinetobacter baumannii, an emerging pathogen in nosocomial infections. Front. Immunol. 2017, 8, 441. [Google Scholar] [CrossRef] [Green Version]
- Henderson, J.C.; Zimmerman, S.M.; Crofts, A.; Boll, J.M.; Kuhns, L.G.; Herrera, C.M.; Trent, M.S. The power of asymmetry: Architecture and assembly of the gram-negative outer membrane lipid bilayer. Annu. Rev. Microbiol. 2016, 70, 255–278. [Google Scholar] [CrossRef]
- Tan, K.; Li, R.; Huang, X.; Liu, Q. Outer membrane vesicles: Current status and future direction of these novel vaccine adjuvants. Front. Microbiol. 2018, 9, 783. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Wang, S.; Yao, Y.; Xia, Y.; Yang, X.; Li, K.; Sun, P.; Liu, C.; Sun, W.; Bai, H.; et al. Employing Escherichia coli-derived outer membrane vesicles as an antigen delivery platform elicits protective immunity against Acinetobacter baumannii infection. Sci. Rep. 2016, 6, 37242. [Google Scholar] [CrossRef]
- Lei, L.; Yang, F.; Zou, J.; Jing, H.; Zhang, J.; Xu, W.; Zou, Q.; Zhang, J.; Wang, X. DNA vaccine encoding OmpA and Pal from Acinetobacter baumannii efficiently protects mice against pulmonary infection. Mol. Biol. Rep. 2019, 46, 5397–5408. [Google Scholar] [CrossRef]
- Ansari, H.; Tahmasebi-Birgani, M.; Bijanzadeh, M.; Doosti, A.; Kargar, M. Study of the immunogenicity of outer membrane protein A (OmpA) gene from Acinetobacter baumannii as DNA vaccine candidate in vivo. Iran. J. Basic Med. Sci. 2019, 22, 669–675. [Google Scholar]
- Rasooli, I.; Abdolhamidi, R.; Jahangiri, A.; Astaneh, S.D.A. Outer membrane protein, Oma87 prevents Acinetobacter baumannii infection. Int. J. Pept. Res. Ther. 2020, 26, 2653–2660. [Google Scholar] [CrossRef]
- Hounsome, J.D.; Baillie, S.; Noofeli, M.; Riboldi-Tunnicliffe, A.; Burchmore, R.J.; Isaacs, N.W.; Davies, R.L. Outerembrane protein A of bovine and ovine isolates of Mannheimia haemolytica is surface exposed and contains host species-specific epitopes. Infect. Immun. 2011, 79, 4332–4341. [Google Scholar] [CrossRef] [PubMed] [Green Version]



| Susceptible Group 1 | Examples | Vaccination Indication |
|---|---|---|
| Inpatients with immune deficiencies or compromising health conditions. | People with hematologic disorders, neutropenia, immune regulatory abnormalities, or receiving splenectomy, organ transplantation; patients kept in ICU or long-term hospitalisation, particularly those intubated. | Ideally vaccination before or soon after being admitted to hospital, especially in medical institutions with historical nosocomial outbreaks. |
| People who are more likely to suffer trauma or receive untimely, inappropriate wound treatments. | Soldiers or peacekeepers serving in ‘hot’ conflict regions; survivors from natural disasters and war; residents of temporary refugee settlements; employees with high-risk jobs such as logging, mining, fishing, emergency response. | Troops assigned to regions with high prevalence of A. baumannii should be vaccinated before deployment; large-scale vaccination in advance, unlikely, thus injured people with severe open wounds should be vaccinated as soon as possible after receiving first aid treatment. |
| People exposed to a high risk of community transmission. | Vulnerable people, e.g., the elderly, in communities with high skin-colonisation or environmental isolation rate, e.g., tropical areas in Asia–Pacific. | People may require vaccination more voluntarily. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, C.; McClean, S. Mapping Global Prevalence of Acinetobacter baumannii and Recent Vaccine Development to Tackle It. Vaccines 2021, 9, 570. https://doi.org/10.3390/vaccines9060570
Ma C, McClean S. Mapping Global Prevalence of Acinetobacter baumannii and Recent Vaccine Development to Tackle It. Vaccines. 2021; 9(6):570. https://doi.org/10.3390/vaccines9060570
Chicago/Turabian StyleMa, Chaoying, and Siobhán McClean. 2021. "Mapping Global Prevalence of Acinetobacter baumannii and Recent Vaccine Development to Tackle It" Vaccines 9, no. 6: 570. https://doi.org/10.3390/vaccines9060570