1. Introduction
Zika virus is associated with ~100-fold increased incidence of severe microcephaly and other neurodevelopmental disorders in newborn babies especially when infection occurs during the first trimester [
1,
2,
3]. It is believed that, like other arboviruses, ZIKV elicits a robust protective immune response, making the development of a vaccine highly desirable. However, vaccine trials in the face of an waning epidemic have proven difficult [
4]. Prophylactic or therapeutic passive immunization with anti-ZIKV immune globulin G (IgG) antibodies, including during pregnancy, has been proposed and tested as an option [
ClinicalTrials.gov studies NCT03624946 (IGIV), NCT03443830 (monoclonal antibody)].
Compared to other endogenous serum proteins, IgG molecules, including monoclonal and polyclonal antibody therapies, have a long half-life, ranging from 11–30 days [
5]. The molecular basis of this phenomenon is the interaction between the neonatal Fc receptor (FcRn) and the Fc domain of the antibody. FcRn, a heterodimeric membrane bound receptor expressed in most mammalian cells interacts with IgG and albumin in a pH-dependent manner [
6]. This interaction, which takes place inside acidified endosomes, prevents endosomal degradation and mediates release of IgG molecules into the lumen of blood vessels. The half-life of IgG is decreased (and clearance increased) when the target immunogen is present. It is postulated that Fc gamma (Fcγ) receptors in immune cells play a role in this expedited clearance [
5]. Studies to assess the contributions of FcRn other receptors in this clearance have not been performed.
Human IgG antibodies are transported across the placenta during pregnancy by an active process mediated by the FcRn [
7]. The maternal-to-fetal antibody transfer can provide protection from infections acquired both in utero and in the early post-natal period. Some viruses, in complexes with antibodies, exploit this essential pathway to cross epithelial [
7] or placental [
8] barriers, leading to fetal infection and disease. Recently, it was proposed that ZIKV antibodies may play a role in placental infection by SARS-CoV-2 [
8], by amplifying infection of placental resident immune (Hofbauer) cells [
9]. It is not known if anti-ZIKV antibodies can similarly enhance infection of placental trophoblast or mediate maternal-to-fetal passage of ZIKV through the FcRn antibody transport pathway.
Here we report the results of experiments measuring disposition of ZIKV IC in two mammalian cell lines that express high levels of FcRn. One is an MDCK cell line overexpressing human FcRn [
10], often used as a model to study FcRn-mediated processes in an epithelial polarized monolayer. The other is a syncytium forming human choriocarcinoma cell line [
11], a commonly used model of the syncytiotrophoblast, the placental structure separating the fetus from the maternal circulation. Using these cell lines, we found that intact ZIKV or the soluble fragment of ZIKV envelope glycoprotein (gpE) can enter and passage through these epithelial cell monolayers. The presence of monoclonal antibodies can either inhibit or enhance cellular entry, depending on their concentration and the cell line used. Furthermore, antibodies degrade more quickly in both cell lines when presented as ICs, compared to when there is no immunogen present. The rate of degradation could be related to the physical-chemical properties of these antibodies, such as the levels of protein aggregation in solution.
2. Methods
2.1. Zika Glycoprotein E (gpE) Immunogen and Zika Virus
Soluble, histidine-tagged recombinant gpE, derived from African strain and expressed in insect cells were obtained commercially (Meridian Life Sciences, Memphis, TN, USA).
Zika virus (Puerto Rican strain) used in this study was isolated by CDC from the serum of a ZIKV infected patient who travelled to Puerto Rico in 2015. The complete genome sequence is published (PRVABC59, Gene bank KU501215). Infectious virus was grown in Vero E6 cells (ATCC) after inoculation at low multiplicity of infection (MOI, 0.2–0.4, for the initial virus preparation obtained from CDC) in MEM media supplemented with 2% FBS. Then the virus was purified based on a published protocol [
12]. Briefly, after 3–9 days (based on cytopathic effect) supernatant was harvested after centrifugation at 4000×
g for 15 min. Virus was concentrated by precipitating overnight with 40% PEG 8000 in NTE buffer (10 mM Tris-HCl, pH 8.0, 120 mM NaCl and 1 mM EDTA) at a final concentration of 8% PEG then centrifuged at 14000×
g for 30 min. The pellet was resuspended in NTE buffer and next purified with 25% sucrose cushioning and centrifugation at 30,000×
g for 1.5 h. All the purification steps were performed at 4 °C. The final titer was determined via a standard plaque (TCID
50) assay in Vero E6 cells.
2.2. Antibodies
The following polyclonal sera and monoclonal immune globulin G antibodies (IgG) were purchased from commercial sources. ZENV14-M (mAb14) and ZENV17-M (mAb17) were purchased from Alpha Diagnostic International (San Antonio, TX, USA). mAb14 is a human monoclonal IgG1 anti-ZIKV gpE domain III that does not cross-react with Dengue or other flaviviruses and was reported to enhance ZIKV infection in K562 cells via Fc gamma receptor (product data sheet). mAb17 is a humanized monoclonal IgG1 anti-flavivirus envelope protein domain II that cross-reacts with other flaviviruses. Purified myeloma IgG1 (Kappa) from human plasma was purchased from Sigma (catalog number I5154-1MG) and used as unrelated control for viral entry experiments.
The following antibodies were used for western blots. To detect gpE: mouse-anti-ZIKV gpE (1:5000, BF-1176-56, BioFront Technologies Sarasota, FL, USA), rabbit-anti-ZIKV gpE (1:1000, ZENV11-A, Alpha Diagnostic International), and donkey-anti-mouse IgG conjugated to HRP (1:50000, A90-337P, Bethyl Laboratories Inc., Montgomery, TX, USA); to detect human IgG: goat-anti-human Fc conjugated to HRP (1:20,000, A80-304P, Bethyl Laboratories Inc., Montgomery, TX, USA).
2.3. Cells
BeWo (human choriocarcinoma cell line) clone b30 was provided by Erik Rytting (University of Texas Medical Branch) and Marvin Darby Canine Kidney Cell line 2 (MDCK2) transfected with human FcRn receptor was provided by Richard Blumberg (Harvard University). The cells were passaged (less than 30 passages) in DMEM Glumax® supplemented with 10% fetal bovine serum (FBS), non-essential amino acids (NEAA) and Antibiotic-Antimycotic mixture (AA, Thermo Fisher Scientific, Pittsburgh, PA, USA).
Vero E6 cells (ATTC) were used to grow and titer ZIKV. They were passaged in MEM medium supplemented with 10% FBS (2% FBS in experiments with ZIKV), NEAA and AA as above.
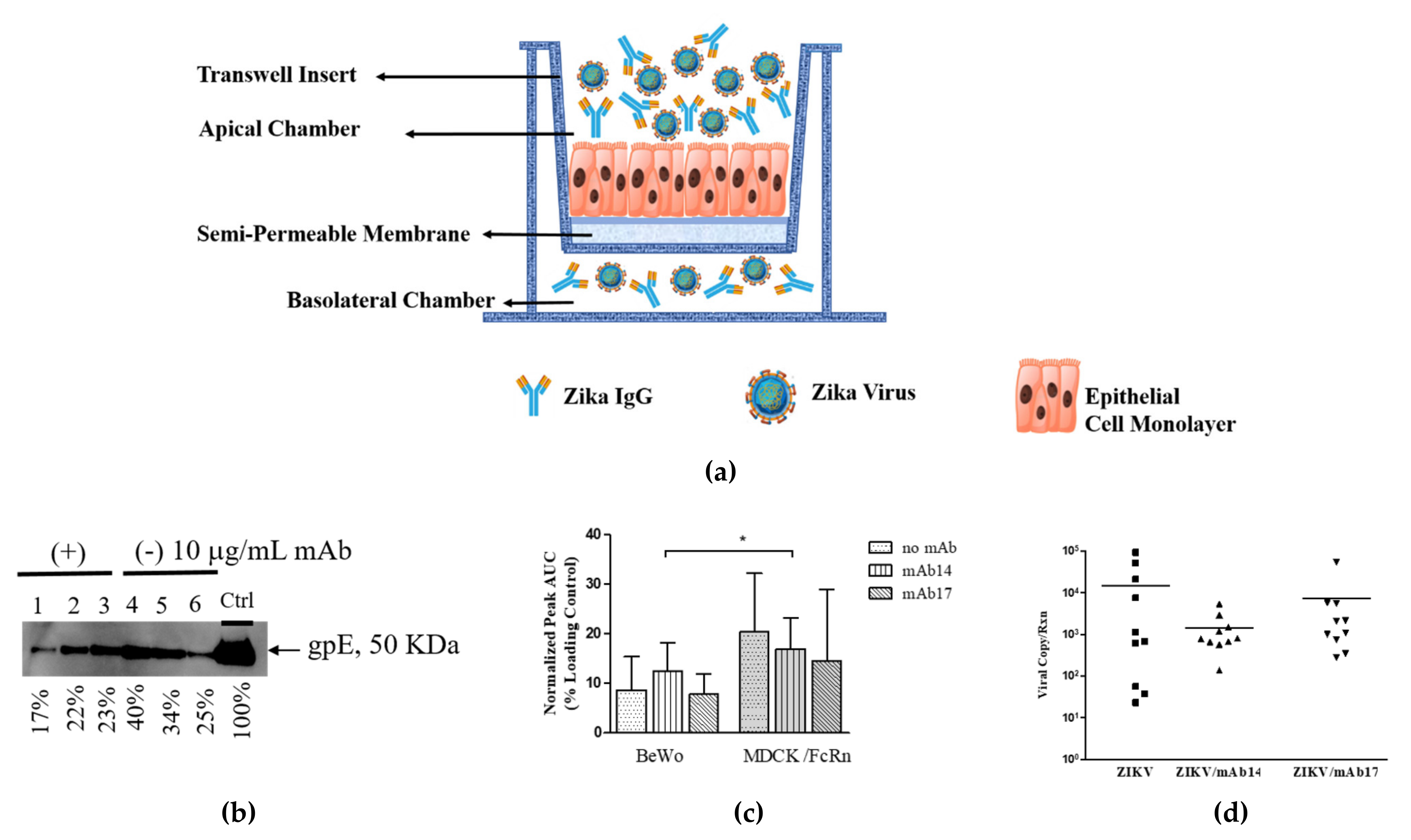
2.4. Assessment of Transcytosis of Immune Complexes (IC) Across Epithelial Cell Layers
Single cell suspensions of MDCK/FcRn (seeding density 1.5 × 10
4 per well) or BeWo (seeding density 1.5 × 10
5 per well) cells were grown in transwell semi-permeable membranes (6.5 mm insert, 0.4 µm pore size, 0.33 cm
2 cell growth area) placed into 24 well plates (Corning, Sigma-Aldrich, St. Louis, MOUSA) in replicates of three per test. Cell growth was monitored by measuring trans epithelial electrical resistance (TEER) via EVOM2 voltohmmeter (World Precision Instruments, Sarasota, FL, USA). When TEER attained 200–250 Ohm for BeWo and 400–450 Ohm for MDCK/FcRn, usually on day 4, the transcytosis experiment was performed. For this gpE alone (10 and 100 µg/mL), IgG-gpE (1:10 and 1:1 weight ratios) IC, or IgG alone (10 and 100 µg/mL) in 0.2 mL pre-warmed Hanks Balanced Salt Solution (HBSS) pH 6.0 were added inside the transwell; 0.2 mL HBSS pH 7.4 was added outside the transwell. IgG-ZIKV IC were generated by mixing 5x10
5 TCID
50 ZIKV (approximate multiplicity of 5 TCID
50 per cell) with 10 µg/mL IgG antibodies inside the transwell, as before. A schematic of the experiment is shown in
Figure 1.
After incubation for 90–120 min at 37 °C, buffer in the basolateral chamber was collected and assessed for the presence of gpE (western blot), ZIKV (PCR), and IgG (ELISA). The wells with increased volume in the basolateral chamber were excluded from measurement due to apparent paracellular transport.
To assess transcytosis of intact ZIKV, viral RNA was extracted from the basolateral buffer using QIAamp
® Viral RNA Kit (
www.qiagen.com) following the manufacturer’s instructions. Ten µL of RNA was used as template for RT qPCR. To assess cell-associated IC, cells attached to the membrane were washed 1× with PBS, then lysed with 60 µL 1× NUPAGE LDS sample buffer with reducing agent and loaded into denaturing gel for Western blots. Gels were run using SDS under denaturing conditions (NuPAGE 4–12% Bis-Tris Protein Gels, Thermo Fisher Scientific, Pittsburgh, PA, USA), transferred onto a nitrocellulose mini membrane using iBlot system (Invitrogen) then assessed for the presence of either gpE or IgG using primary and secondary antibodies (see
Antibodies). The same denaturing gels and Western blotting procedure were used to assess gpE transcytosis. A fixed amount of mAb or gpE was loaded in all the gels to serve as an internal normalizer for quantification (see
Data Processing and Statistical Analysis for more details). A commercial kit (ELISA Kit, EI7200-1, AssayPro, St. Charles, MO, USA) was used to quantitate antibody transcytosis. The experiment was repeated on a different day to confirm the results.
2.5. Assessment of ZIKV IC Cell Entry
Viral entry assays were performed as reported in literature [
13], with the modification that crude cell lysates (instead of purified RNA) were used for PCR analysis. Immune complexes were prepared in triplicate by mixing 10 µL virus with 10 µL IgG solution or dilution media (2% FBS in DMEM) and incubated on ice for approximately 1 h. An unrelated IgG1 antibody purified from human myeloma (Sigma-Aldrich, St. Louis, MO, USA) was used as negative control. Pre-formed IC or ZIKV alone were next added onto 5 × 10
5 MDCK/FcRn or BeWo cells (approximate multiplicity of 100 TCID
50 per cell) and incubated at 37 °C for one hour. Viral entry was halted by placing the cells on ice, followed by washing with cold PBS. The plates were next incubated over ice with Protease K (500 ng/mL) (Promega) for one hour, to digest any residual ZIKV attached to cell surfaces. Cells were washed three times with cold PBS and cell pellets were lysed with 200 uL iScript buffer (Bio-Rad) following the manufacturer’s instructions. The lysate was centrifugated at 20,000×
g for three minutes, and supernatants were transferred into fresh tubes. These supernatants were diluted 1:100. RT-qPCR was performed on 4 uL aliquots of each sample, for ZIKV and GAPDH (see below). The experiment was repeated on a different day to confirm results (unrelated IgG controls were run in triplicate, one time only).
2.6. PCR
To assess the amount of ZIKV in purified RNA (transcytosis experiments) or crude cell lysates, one-step quantitative RT PCR was used (TaqMan
TM RNA-to-CT
TM 1-Step Kit from Applied Biosystems,
www.thermofisher.com); primers were chosen to allow for high sensitivity as reported [
14], and were synthesized by Integrated DNA Technologies, Inc. (
www.idtdna.com). Each 50 or 10 uL PCR reaction solution contained 0.5 µM primer, 0.25 µM probe and RT-PCR master mix master as per manufacturer’s instructions. The PCR reaction and analysis were performed in a QuantStudio3 Q-PCR instrument and software from Applied Biosystems (
www.Thermofisher.com). All samples were run in duplicate.
For transcytosis experiments the levels of ZIKV RNA were quantified using a standard curve, prepared using well-characterized material produced at FDA and harmonized during testing for the development of the WHO International Standard for ZIKV RNA [
15]. Purified RNA from concentrated and titrated ZIKV was used to construct the standard curve. For the viral entry experiments ZIKV RNA was quantified using ∆∆C
t method, with GAPDH as the endogenous control (housekeeping) gene. Control wells containing ZIKV alone were used to establish infection baselines in the absence of IgG,
Table 1.
2.7. Characterization of IC
2.7.1. Surface Plasmon Resonance (SPR) (Biacore)
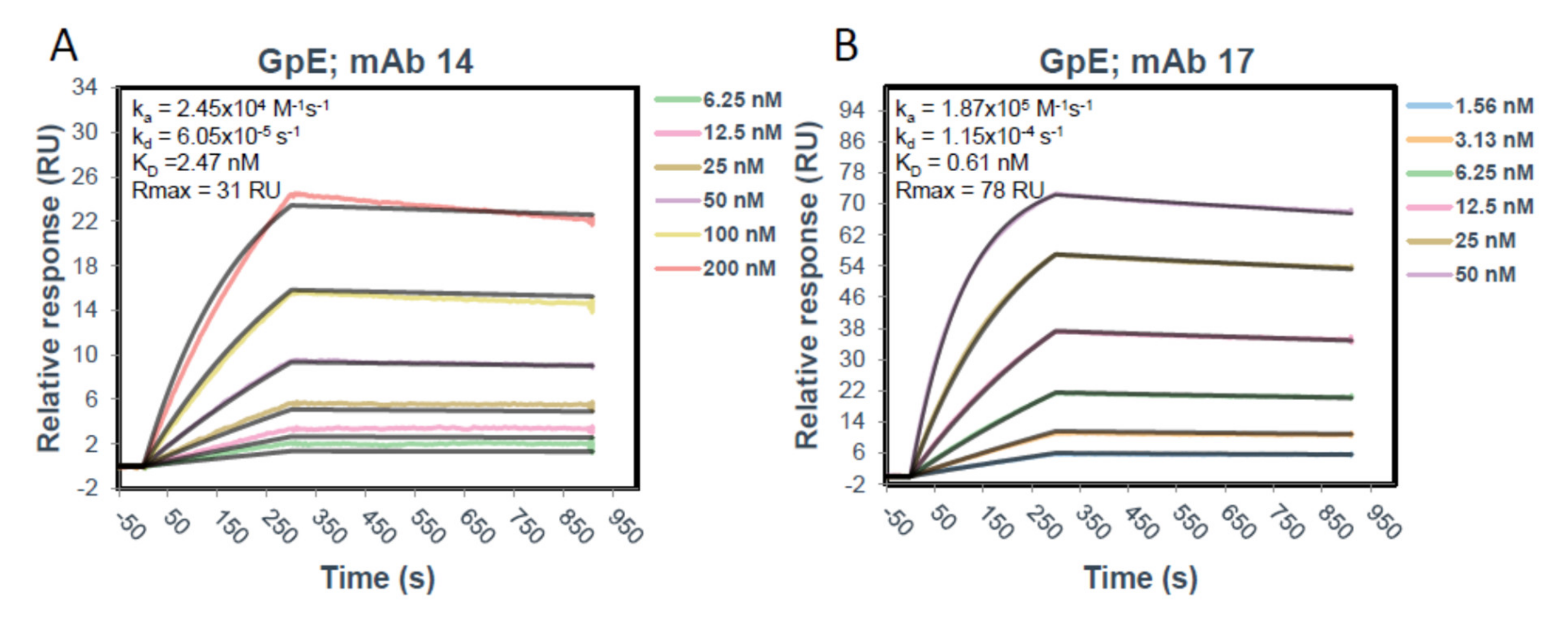
The SPR experiments were performed on a Biacore T200 instrument (Cytiva, Marlborough, MA, USA). A Human Antibody Capture Kit (Cytiva) was used to capture both mAb14 and mAb17 onto a CM5 chip (Cytiva). Briefly, the anti-human IgG (Fc) antibody provided at 0.5 mg/mL in 0.15 M NaCl was diluted to 25 µg/mL in Immobilization buffer (10 mM sodium acetate pH 5.0). Immobilization of antibody onto the CM5 chip was performed using an Amine Coupling Kit (Cytiva) and each following step was performed at 5 µL/min for 7 min. Three flow cells (Fc1, Fc2, and Fc3) of a CM5 chip (Cytiva) were activated with a 1:1 mixture of 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide hydrochloride (EDC) and N-hydroxysuccinimide (NHS) after which the diluted anti-human IgG (Fc) was immobilized saturating the surface at 13,300–14,200 RU. The surface was deactivated using 1.0 M ethanolamine-HCl pH 8.5. Following immobilization of the capture antibody, mAb14 and mAb17 were diluted to 3 µg/mL in 0.01 M HEPES pH 7.4, 0.15 M NaCl, 0.005% v/v Surfactant P20 (HBS-P) and captured on Fc2 and Fc3, respectively, for 40 sec. at a flow rate of 10 µL/min to reach approximately 150 RU, aiming for a Rmax of 50 RU. Fc1 was kept blank as a reference surface for buffer subtraction (capture antibody only). Glycoprotein E was diluted to 200 nM starting concentration in HBS-P and then in 1:2 dilutions (200–3.125 nM) was run over each captured mAb and reference surface at a flow rate of 50 µL/min for 5 min after running 3 startup cycles with HBS-P buffer only over all the surfaces under the same conditions. The mAbs were allowed to dissociate for 10 min after which the surface was regenerated down to the anti-human Fc antibody using 3M magnesium chloride at a 20 µL/min flow rate for 30 s. The resulting sensograms were analyzed using in Biacore Insight Evaluation Software and fit using a 1:1 Langmuir model to obtain kinetics and affinity data.
2.7.2. Dynamic Light Scattering (DLS)
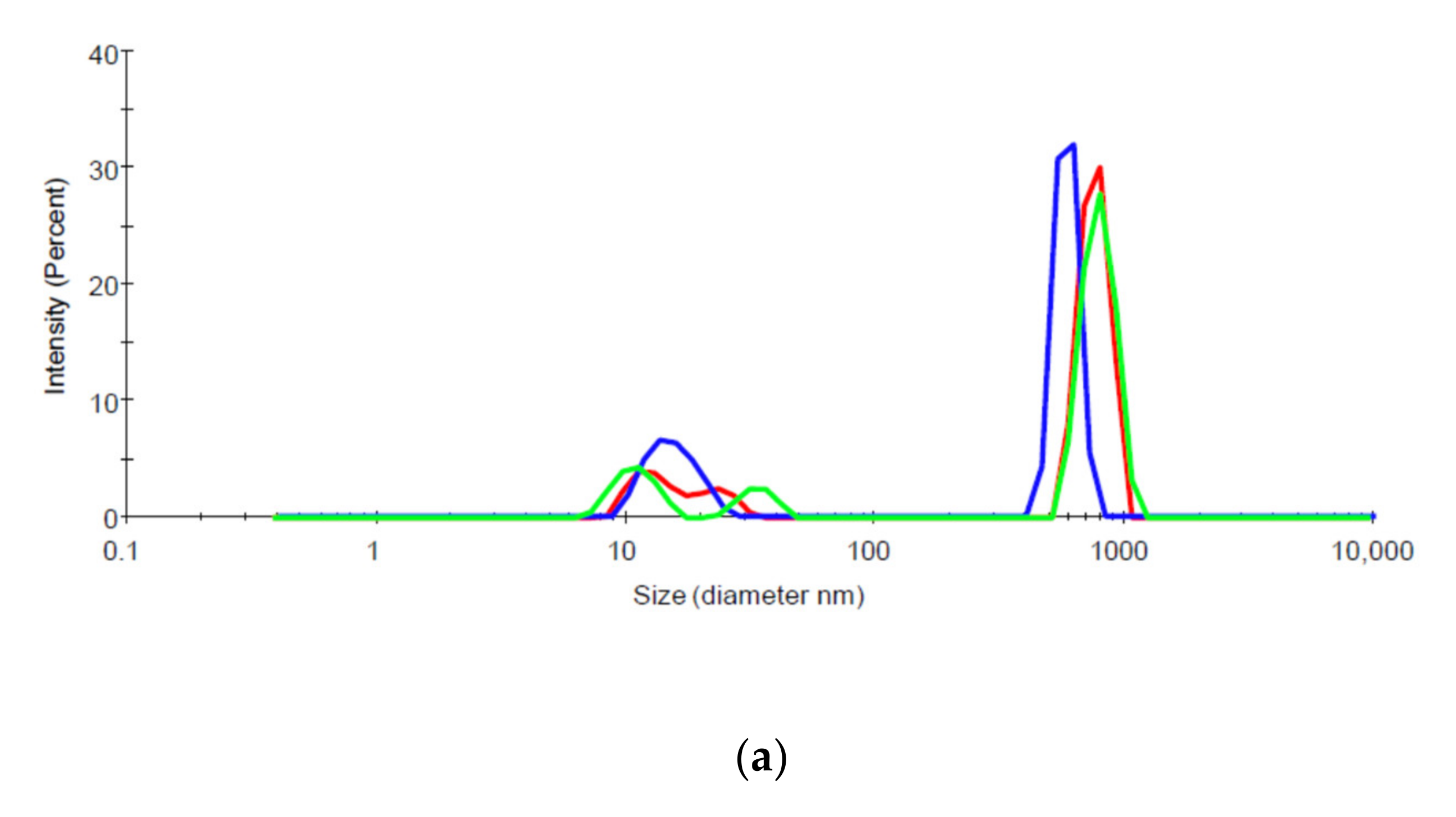
Aggregation of biologic preparations can be assessed by measuring hydrodynamic diameters, which estimates the size and distribution of sizes of particles in solution. A Zetasizer Nano ZS (Malvern Instruments Ltd., Westborough, MA, USA) DLS instrument with 173° detection optics was used to measure estimate the size and size distribution of mAb14, mAb17, gpE, and their 1:1 molar mixtures. The samples were analyzed using a standard protein protocol with the material refractive index (RI) set at 1.45. The dispersant parameters were set based on the RI and viscosity of PBS, which were 1.330 and 0.8882, respectively. The measurement position and attenuator settings were automatically determined by the instrument. Three measurements were performed on each sample.
2.8. Data Processing and Statistical Analysis
ELISA readings were transformed into human IgG concentrations using a five-parameter fit of the standard curve (SoftMax Pro, Molecular Devices, San Jose, CA, USA). The concentration was expressed as a percentage of the IgG concentration added to the input chamber and analyzed using 2-way ANOVA with GraphPad Prism 5 (GraphPad Software, Inc., San Diego, CA, USA) with ICs as the column factor and mAb14 or 17 as the row factor; Bonferroni post-hoc analysis was used to correct for multiple comparisons.
Western blots were scanned using GBOX Mini (Syngene, syngene.com) imaging system. The image files were then analyzed with image processing software ImageJ (open source, version 1.52a). The bands for gpE or mAb were selected and plotted, then the area under the density peak curve (AUC) for each band was calculated using the software tools. Experimental AUCs were divided by the AUC of concurrent loading controls to allow for comparisons between multiple experiments. The differences in normalized AUCs was analyzed with 2-way ANOVA, with Bonferroni correction using GraphPad Prism 5.
ZIKV cellular entry in the presence or absence of mAb was quantified using ∆∆Ct method with GAPDH as the internal control. Briefly, human or canine GAPDH levels were used to calculate ∆Ct for BeWo and MDCK/FcRn cells, respectively. ∆Ct values from the experiments with ZIKV alone were used as the baseline to calculate ∆∆Ct. Difference in cellular ZIKV RNA in the presence versus absence of mAb was then calculated using the expression: Fold Change = 2(−∆∆Ct). The data set was analyzed with 2-way ANOVA as before using GraphPad Prism 5; p < 0.05 was considered significant.
4. Discussion
ZIKV and other flaviviruses enter cells through attachment factors and specific receptors that mediate endocytosis of the virus and subsequent release of the viral contents into the cell [
16]. Receptors include DC-SIGN, the mannose receptor, and members of the TIM and TAM family of phosphatidylserine receptors [
17]. Antibodies against ZIKV can protect via various well described mechanisms, including blockage of cell entry through direct binding to envelope glycoprotein and subsequent destruction of pathogens by effector cells. However, it is possible for virus-antibody complexes to enter immune cells through various Fc receptors, be carried as cargo in endosomes, followed by endosomal acidification and membrane fusion, which permits release of viral RNA into the cytoplasm and subsequent viral replication. This mechanism has been identified as the reason for increased disease severity following Dengue virus (DENV) infections in subjects with existing immunity to DENV of a different serotype [
18,
19]. The phenomenon, referred to as antibody dependent enhancement (ADE), has been proposed to occur for ZIKV in vitro and in animal studies [
20,
21]. It is thus important that, before being tested in clinical trials, antibody preparations targeting ZIKV be assessed for their ability to block infection without the possibility of ADE. Such tests are often performed in animal models of infection and in cell lines that express Fc-gamma receptors. Although not often discussed, the FcRn, an Fc-receptor highly expressed in placenta, could also mediate ADE. We set out to test this hypothesis using a syncytium-forming human trophoblast cell line and dog kidney epithelial cells engineered to overexpress human FcRn.
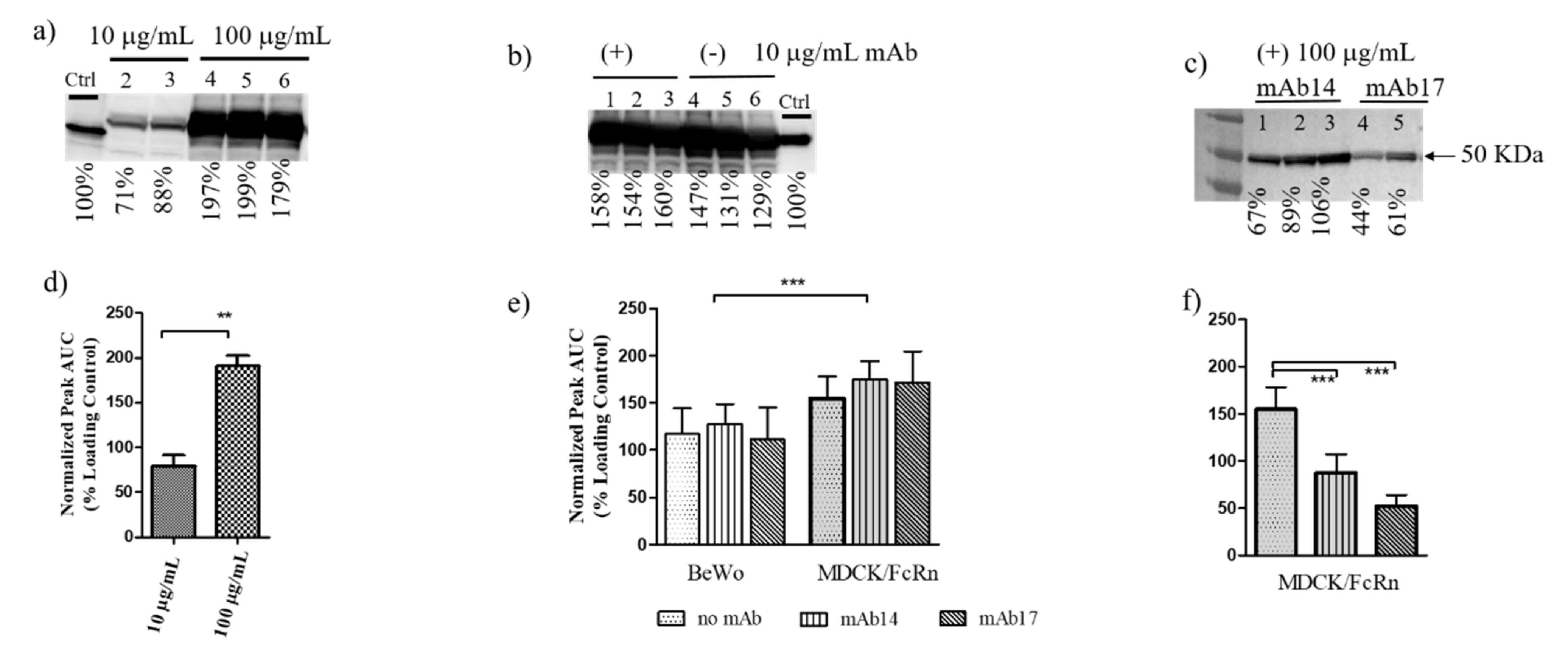
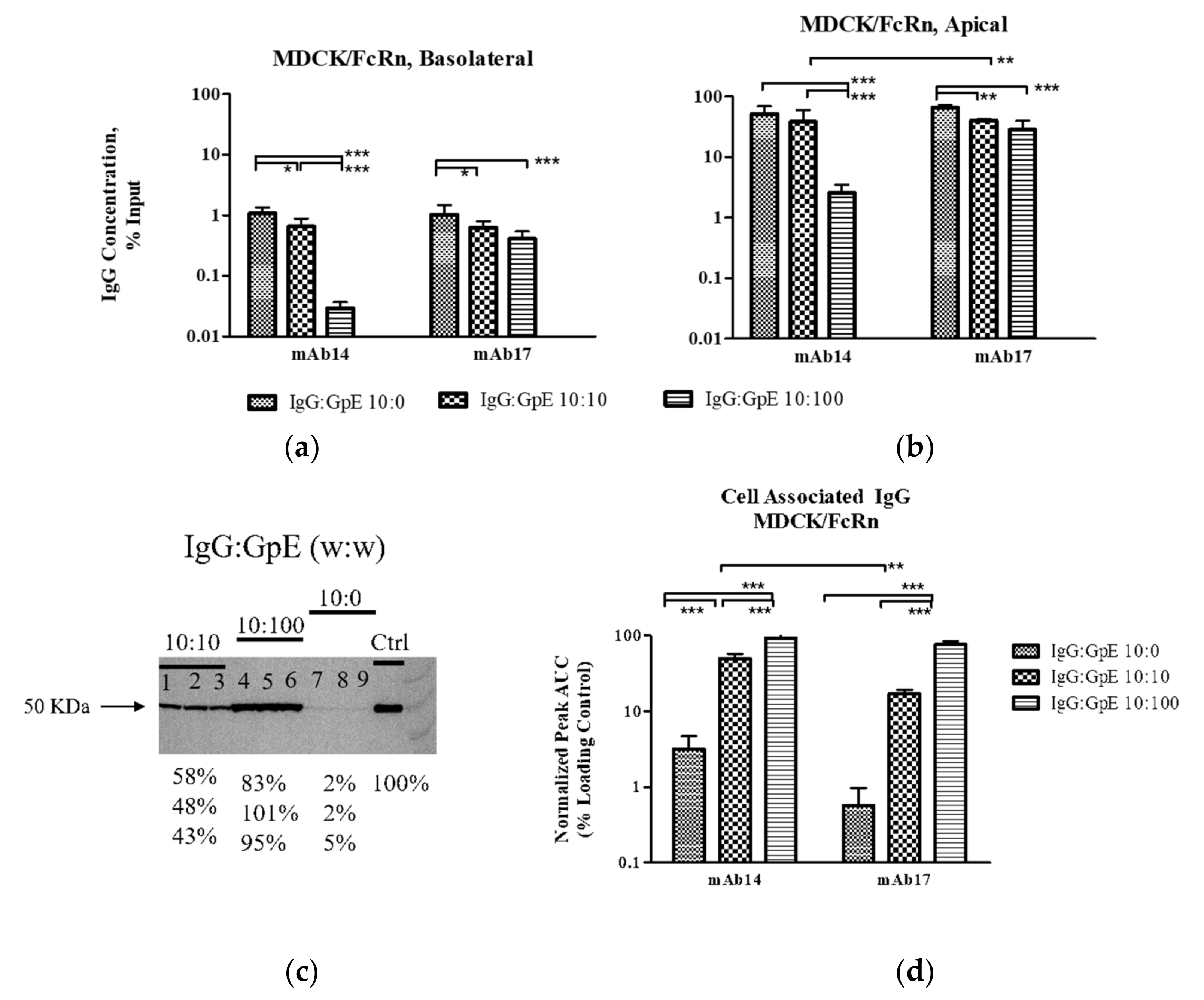
We showed that ZIKV envelope glycoprotein alone or in complex with IgG can be transported through MDCK/FcRn and BeWo epithelial cells (
Figure 1 and
Figure 2). Although not surprising given these cells’ permissiveness to infection by ZIKV [
22,
23,
24], to our knowledge this is the first time this observation has been made. Both cell entry and transcytosis of gpE is greater in MDCK compared to BeWo cells (
Figure 1c and
Figure 2e), which correlates with reduced susceptibility of the latter to ZIKV infection [
24]. Intact virus was also transcytosed in MDCK cells (
Figure 1d), as it has been reported for the HULEC-5a endothelial cell line [
21]. It is not known if such passages through cell monolayers over a short time (transcytosis was assessed at 90 min) occurs for other viruses. Polarized entry and release of ZIKV was shown in Caco-2 cell monolayers [
25], indicating that directional transport is a feature of this virus and could play a role in viral uptake and infectivity.
Given that transcytosis in MDCK/FcRn and BeWo cells occurs for both gpE and intact ZIKV, it is likely that the process is mediated (at least in part) by ZIKV receptors that directly interact with gpE. In such a case, the addition of anti-gpE IgG would compete with receptor binding and thus cause a reduction of cell entry and transcytosis. Indeed, while the addition of sub-stochiometric amounts (10 µg/mL) of mAb14 and mAb17 resulted in non-significant changes in transcytosis and cell entry of gpE (
Figure 1b,c), approximate stochiometric amounts (100 µg/mL) significantly reduced entry (mAb14 and mAb17,
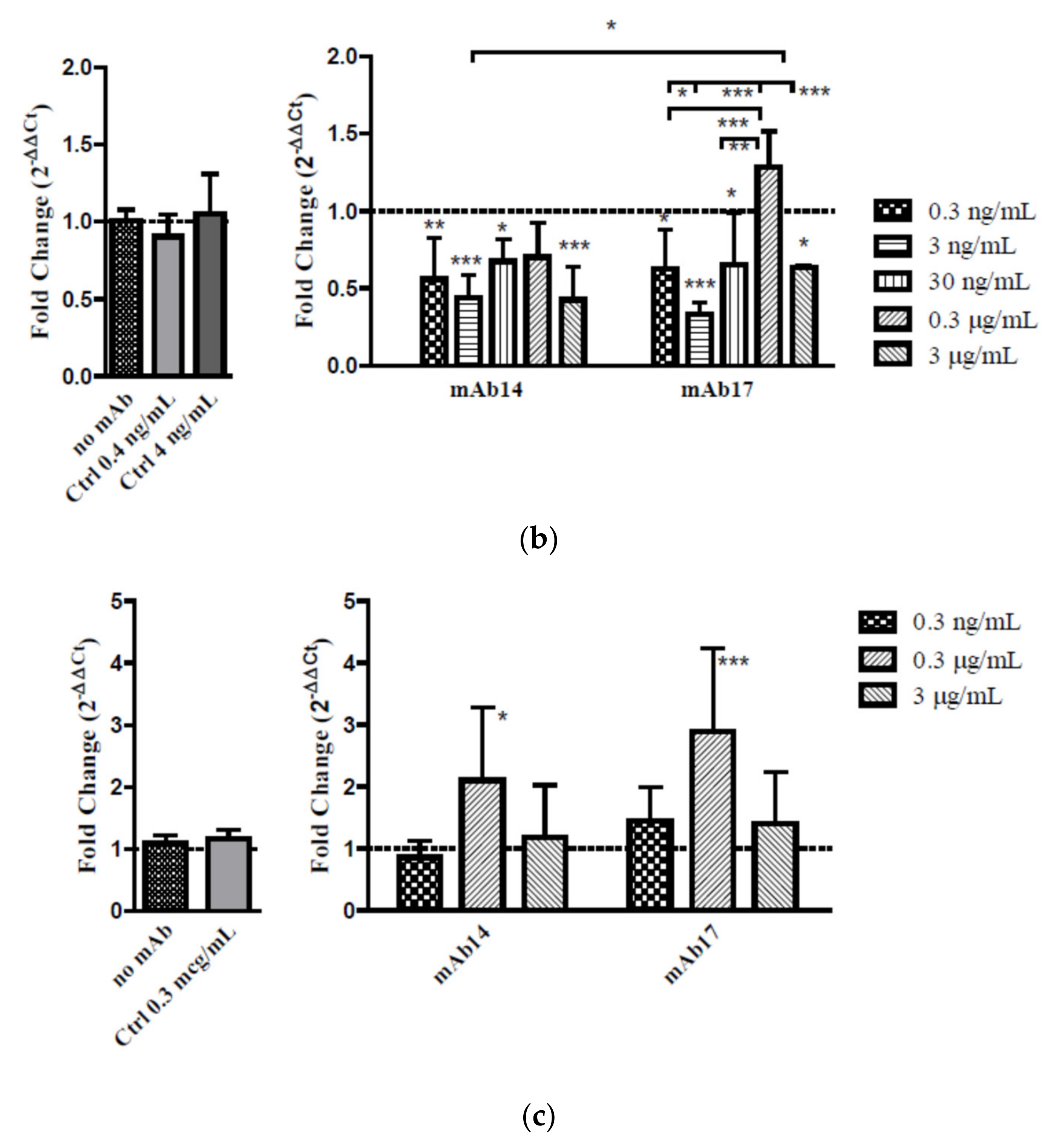
Figure 2e) in MDCK/FcRn cells. To better address the effect of the IgG concentration on viral entry, we performed carefully controlled experiments of ZIKV entry in cell suspension (
Figure 4). We diluted mAb14 and 17 over a wide range, from 0.3 ng/mL to 3 µg/mL and, for each dilution measured the effect in reducing or enhancing viral entry. These studies showed that, in cells that overexpress FcRn, a stepwise increase in IgG concentration first decreased then increased viral entry. A second reduction of viral entry was seen at a high concentration of IgG. An analogous bimodal behavior was recently described for MERS coronavirus in presence of Mersmab1.
A similar concentration dependence of ZIKV entry was seen in BeWo cells. The lowest (0.3 ng/mL) and highest (3 µg/mL) concentrations tested had little effect on viral entry, whereas intermediate concentration (0.3 µg/mL) resulted in enhanced viral entry. The lack of significant inhibition of viral entry in BeWo cells comports with lower infectivity of ZIKV in these cells [
24] and likely results from lower levels of gpE-specific receptors in these cells. On the other hand, unlike in MDCK/FcRn cells, the enhancement of viral entry in BeWo cells at intermediate concentrations was statistically significant, of higher magnitude, and occurred for both mAbs.
The antibodies we used have an affinity for gpE equal to 2.5 and 0.6 nM (
Figure 5), thus would be expected to bind gpE and block viral entry through gpE specific receptors even at very low concentrations. While a 50–60% reduction of viral entry at low concentrations in MDCK/FcRn cells was not surprising, it was unexpected that increasing antibody concentrations abrogated this inhibitory effect. We postulate that raising IgG concentrations would result in more immune complexes and higher cell entry via antibody Fc mediated FcRn receptor binding. This would negate some of the Fab mediated blockade of ZIKV entry. That this increased viral entry occurred at intermediate IgG concentrations would be in agreement with the µM-level affinity of the IgG for human FcRn [
26,
27]. At yet higher concentrations of IgG, it is conceivable that excess unbound antibodies would compete with ZIKV IC for entry through FcRn receptor, thus resulting in a second reduction of cellular entry. It is possible that other mechanisms, such as increased degradation could also be involved.
Our experiments suggest that, at specific concentrations, even high affinity IgG antibodies may not reduce and could enhance ZIKV entry in FcRn bearing cells. Such enhancement was recently demonstrated in vivo and in vitro [
21]. Specifically, pre-existing DENV antibodies resulted in enhanced fetal disease in wild type mice but not FcRn
-/- mice. In addition, high concentrations (200–500 µg) of anti-flavivirus cross-reactive antibody together with ZIKV on the apical side of endothelial cells resulted in enhanced infection of trophoblast cells on the basolateral site. Our in vitro studies point to increased ZIKV viral entry in placental cells in the presence of an anti-flavivirus antibody (mAb17) as well as an anti-ZIKV specific monoclonal antibody, mAb14. Notably, mAb14 has been shown to enhance infection of K562 immune cells (Alpha Diagnostic International, Product Data Sheet), leading us to suspect that an Fc-mediated viral entry process underlies the enhancement in both Fc-gamma and FcRn bearing cells. Our data suggests that the concentration of cross-reacting and ZIKV-specific antibodies plays a critical role in inhibition versus viral entry, which underscores the importance of in vitro and in vivo testing of any antibody based prophylactic or therapeutic treatment. More studies are needed to better understand the process of ZIKV transcytosis, antibody mediated enhancement of viral entry and the relevance of these findings in ZIKV infection and placental transport in vitro and in vivo.
Another interesting finding emerged when we assessed disposition of the antibody element of the ICs. Increasing amounts of gpE were associated with decreased amounts of IgG transcytosis and a simultaneous depletion of the IgG in the input reservoir. This observation supports the possibility that more mAb are channeled towards degradation pathways rather than undergoing sorting and recycling as is the case for un-complexed antibodies. Furthermore, mAb14, was depleted faster than mAb17. This antibody had both lower affinity for gpE (
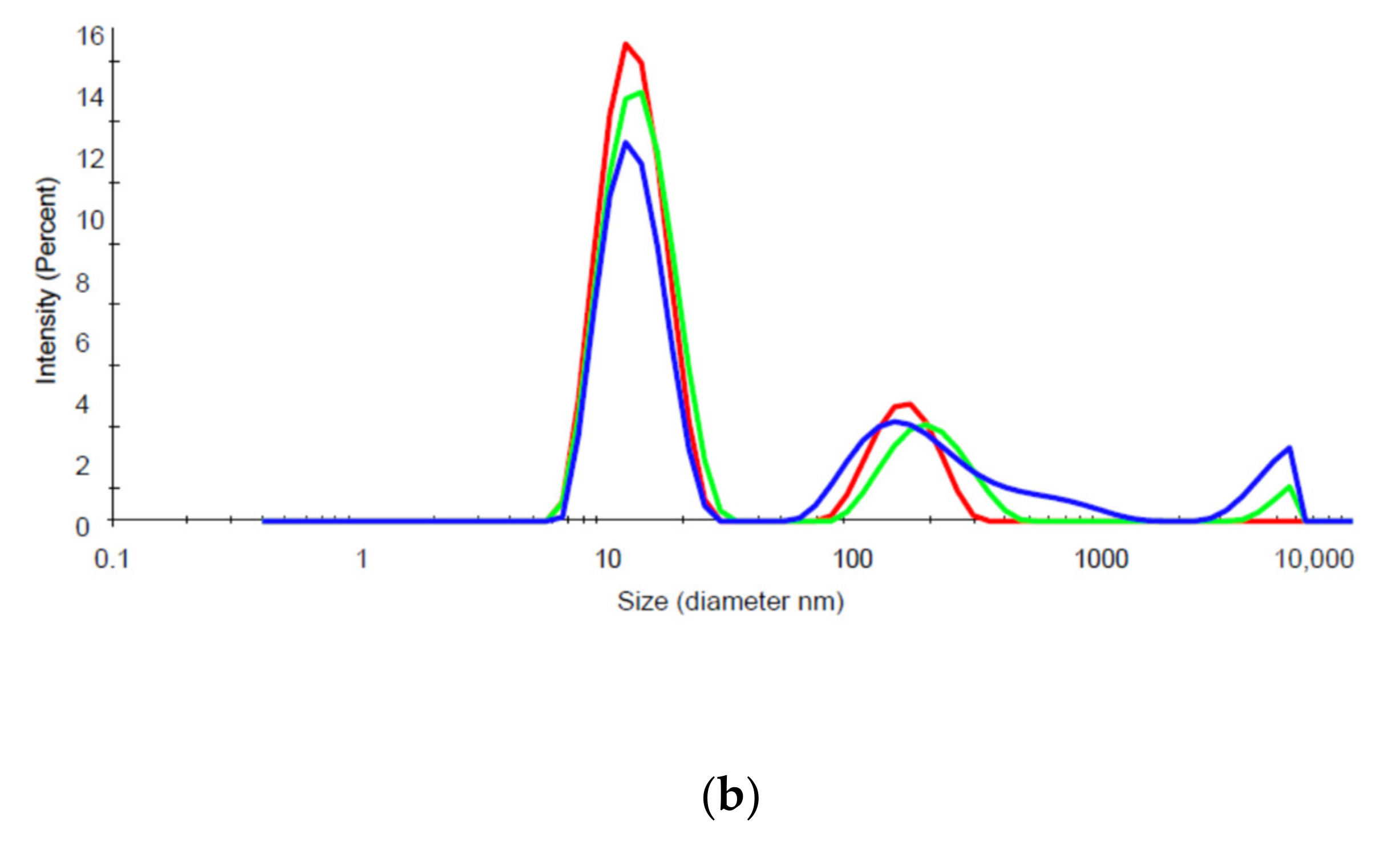
Figure 5) and greater aggregation in solution (
Figure 6). It is possible that these less favorable physicochemical properties of mAb14 play a role in its degradation.
FcRn is crucial in maintaining IgG homeostasis in circulation [
6] and its blockade has been implicated in reduction of plasma IGG and autoimmune complexes [
28]. It has been recently proposed that in vivo half-life of mAb correlates with transcytosis in cells that overexpress FcRn [
29]. Although mechanistic studies are scarce, it is believed that the degradation of immune complexes occurs inside vacuoles, i.e., endosomes and lysosomes following pinocytosis or via receptor-mediated entry [
30]. An alternative pathway could be through proteasomes, perhaps through a process mediated by TRIM21, an intra-cellular Fc receptor and ubiquitin ligase. In this pathway, the antigen/antibody complex passes through the membrane of the endosome and enters the cytoplasm [
31]. In our study we observed that larger IC undergo less transcytosis and seem to be degraded faster than the un-complexed or less aggregated antibody. We postulate that an increased degradation of such larger complexes may be due to the engagement of several cellular pathways for proteolysis, for example lysosome and proteasome pathways. Larger aggregates would engage the lysosome pathway more readily and effectively than smaller ones, thus promoting a faster degradation. More studies are needed to better understand these pathways, and their significance in virus and antibody clearance in vivo.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}