Harnessing Cellular Immunity for Vaccination against Respiratory Viruses


Abstract
:1. Introduction
2. History of Vaccine Enhanced Disease
3. Overview of Immune Response to Respiratory Viruses
3.1. T Cell Immunity
3.2. Dendritic Cell Role during Respiratory Viral Infection
3.3. Trained Immunity
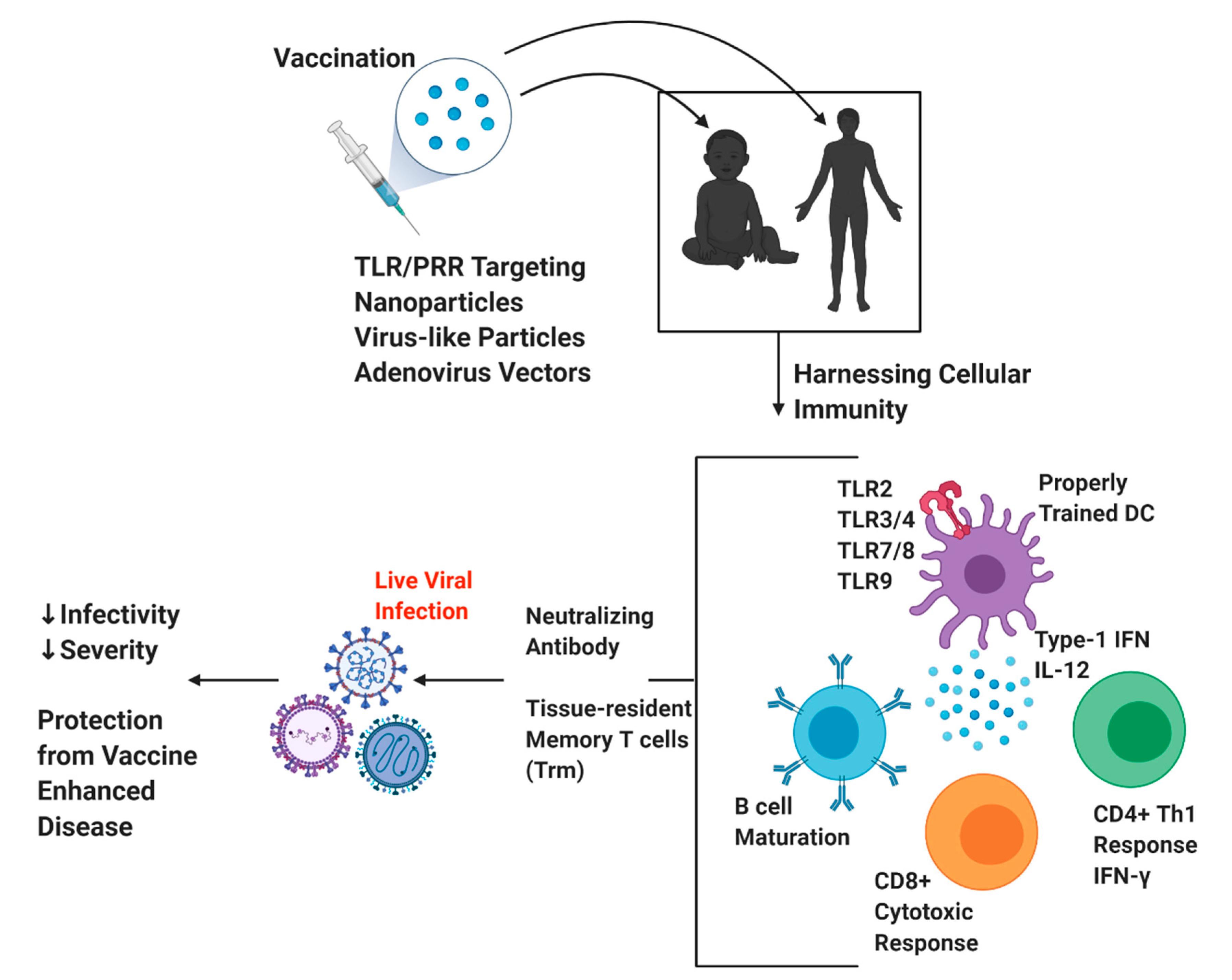
4. Novel Vaccination Approaches against Respiratory Viral Infections
4.1. Targeting T Cells during Vaccine Development
4.2. Importance of Proper TLR/APC Signaling and Vaccine Adjuvanticity
4.3. Current Vaccination Strategies
4.3.1. Nanoparticle-Based Vaccines
4.3.2. Virus-Like Particle Vaccines
4.3.3. Adenoviral Vector Vaccines
4.3.4. Maternal Vaccination for Early-Life Prevention
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Troeger, C.; Forouzanfar, M.; Rao, P.C.; Khalil, I.; Brown, A.; Swartz, S.; Fullman, N.; Mosser, J.; Thompson, R.L.; Reiner, R.C.; et al. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory tract infections in 195 countries: A systematic analysis for the Global Burden of Disease Study 2015. Lancet Infect. Dis. 2017, 17, 1133–1161. [Google Scholar] [CrossRef] [Green Version]
- Nair, H.; Simões, E.A.; Rudan, I.; Gessner, B.D.; Azziz-Baumgartner, E.; Zhang, J.S.F.; Feikin, D.R.; Mackenzie, G.A.; Moiïsi, J.C.; Roca, A.; et al. Global and regional burden of hospital admissions for severe acute lower respiratory infections in young children in 2010: A systematic analysis. Lancet 2013, 381, 1380–1390. [Google Scholar] [CrossRef] [Green Version]
- Alexander-Miller, M.A. Vaccines against Respiratory Viral Pathogens for Use in Neonates: Opportunities and Challenges. J. Immunol. 2014, 193, 5363–5369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borchers, A.T.; Chang, C.; Gershwin, M.E.; Gershwin, L.J. Respiratory Syncytial Virus—A Comprehensive Review. Clin. Rev. Allergy Immunol. 2013, 45, 331–379. [Google Scholar] [CrossRef] [PubMed]
- Ruckwardt, T.J.; Morabito, K.M.; Graham, B.S. Determinants of early life immune responses to RSV infection. Curr. Opin. Virol. 2016, 16, 151–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Liu, L.; Shi, P.; Jiang, G.; Jia, P.; Wang, C.; Wang, L.; Qian, L. Risk factors for acute respiratory syncytial virus infection of lower respiratory tract in hospitalized infants. Zhonghua Chin. J. Pediatr. 2014, 52, 373–377. [Google Scholar]
- Salvati, L.; Biagioni, B.; Vivarelli, E.; Parronchi, P. A gendered magnifying glass on COVID-19. Clin. Mol. Allergy 2020, 18, 1–11. [Google Scholar] [CrossRef]
- Onder, G.; Rezza, G.; Brusaferro, S. Case-Fatality Rate and Characteristics of Patients Dying in Relation to COVID-19 in Italy. JAMA 2020, 323, 1775–1776. [Google Scholar] [CrossRef]
- Lipsky, M.S.; Hung, M. Men and COVID-19: A Pathophysiologic Review. Am. J. Men’s Health 2020, 14. [Google Scholar] [CrossRef]
- Sigurs, N.; Aljassim, F.; Kjellman, B.; Robinson, P.D.; Sigurbergsson, F.; Bjarnason, R.; Gustafsson, P.M. Asthma and allergy patterns over 18 years after severe RSV bronchiolitis in the first year of life. Thorax 2010, 65, 1045–1052. [Google Scholar] [CrossRef] [Green Version]
- Nair, H.; Nokes, D.J.; Gessner, B.D.; Dherani, M.; Madhi, S.A.; Singleton, R.J.; O’Brien, K.L.; Roca, A.; Wright, P.F.; Bruce, N.; et al. Global burden of acute lower respiratory infections due to respiratory syncytial virus in young children: A systematic review and meta-analysis. Lancet Lond. Engl. 2010, 375, 1545–1555. [Google Scholar] [CrossRef] [Green Version]
- Jackson, D.J.; Gern, J.E.; Lemanske, R.F. The contributions of allergic sensitization and respiratory pathogens to asthma inception. J. Allergy Clin. Immunol. 2016, 137, 659–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busse, W.W.; Lemanske, R.F.; Gern, J.E. Role of viral respiratory infections in asthma and asthma exacerbations. Lancet Lond. Engl. 2010, 376, 826–834. [Google Scholar] [CrossRef]
- Marshall, M. The lasting misery of coronavirus long-haulers. Nature 2020, 585, 339–341. [Google Scholar] [CrossRef] [PubMed]
- Understanding the long-term health effects of COVID-19. EClinicalMedicine 2020, 26, 100586. [CrossRef] [PubMed]
- Fulginiti, V.A.; Eller, J.J.; Sieber, O.F.; Joyner, J.W.; Minamitani, M.; Meiklejohn, G. Respiratory virus immunization. I. A field trial of two inactivated respiratory virus vaccines; an aqueous trivalent parainfluenza virus vaccine and an alum-precipitated respiratory syncytial virus vaccine. Am. J. Epidemiol. 1969, 89, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Kapikian, A.Z.; Mitchell, R.H.; Chanock, R.M.; Shvedoff, R.A.; Stewart, C.E. An epidemiologic study of altered clinical reactivity to respiratory syncytial (RS) virus infection in children previously vaccinated with an inactivated RS virus vaccine. Am. J. Epidemiol. 1969, 89, 405–421. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Canchola, J.G.; Brandt, C.D.; Pyles, G.; Chanock, R.M.; Jensen, K.; Parrott, R.H. Respiratory syncytial virus disease in infants despite prior administration of antigenic inactivated vaccine. Am. J. Epidemiol. 1969, 89, 422–434. [Google Scholar] [CrossRef]
- Smatti, M.K.; Al Thani, A.A.; Yassine, H.M. Viral-Induced Enhanced Disease Illness. Front. Microbiol. 2018, 9, 2991. [Google Scholar] [CrossRef] [Green Version]
- Acosta, P.L.; Caballero, M.T.; Polack, F.P. Brief History and Characterization of Enhanced Respiratory Syncytial Virus Disease. Clin. Vaccine Immunol. 2016, 23, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Varga, S.M. Fixing a failed vaccine. Nat. Med. 2009, 15, 21–22. [Google Scholar] [CrossRef] [PubMed]
- Schuler, C.F.; Malinczak, C.-A.; Best, S.K.K.; Morris, S.B.; Rasky, A.J.; Ptaschinski, C.; Lukacs, N.W.; Fonseca, W. Inhibition of uric acid or IL-1β ameliorates respiratory syncytial virus immunopathology and development of asthma. Allergy 2020, 75, 2279–2293. [Google Scholar] [CrossRef] [PubMed]
- Malinczak, C.-A.; Fonseca, W.; Rasky, A.J.; Ptaschinski, C.; Morris, S.; Ziegler, S.F.; Lukacs, N.W. Sex-associated TSLP-induced immune alterations following early-life RSV infection leads to enhanced allergic disease. Mucosal Immunol. 2019, 12, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Openshaw, P.J.M.; Tregoning, J.S. Immune Responses and Disease Enhancement during Respiratory Syncytial Virus Infection. Clin. Microbiol. Rev. 2005, 18, 541–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kool, M.; Pétrilli, V.; Smedt, T.D.; Rolaz, A.; Hammad, H.; van Nimwegen, M.; Bergen, I.M.; Castillo, R.; Lambrecht, B.N.; Tschopp, J. Cutting Edge: Alum Adjuvant Stimulates Inflammatory Dendritic Cells through Activation of the NALP3 Inflammasome. J. Immunol. 2008, 181, 3755–3759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosmann, T.R.; Sad, S. The expanding universe of T-cell subsets: Th1, Th2 and more. Immunol. Today 1996, 17, 138–146. [Google Scholar] [CrossRef]
- Zhu, J.; Yamane, H.; Paul, W.E. Differentiation of Effector CD4 T Cell Populations. Annu. Rev. Immunol. 2010, 28, 445–489. [Google Scholar] [CrossRef] [Green Version]
- Swain, S.L.; McKinstry, K.K.; Strutt, T.M. Expanding roles for CD4+ T cells in immunity to viruses. Nat. Rev. Immunol. 2012, 12, 136–148. [Google Scholar] [CrossRef]
- Kang, S.; Brown, H.M.; Hwang, S. Direct Antiviral Mechanisms of Interferon-Gamma. Immune Netw. 2018, 18, e33. [Google Scholar] [CrossRef]
- Romagnani, S. Th1/Th2 cells. Inflamm. Bowel Dis. 1999, 5, 285–294. [Google Scholar] [CrossRef]
- Guglani, L.; Khader, S.A. Th17 cytokines in mucosal immunity and inflammation. Curr. Opin. HIV AIDS 2010, 5, 120–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, L.; Sagfors, A.M.; Openshaw, P.J.M.; Culley, F.J. Immunity to RSV in Early-Life. Front. Immunol. 2014, 5, 466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Lindell, D.M.; Berlin, A.A.; Morris, S.B.; Shanley, T.P.; Hershenson, M.B.; Lukacs, N.W. IL-17-induced pulmonary pathogenesis during respiratory viral infection and exacerbation of allergic disease. Am. J. Pathol. 2011, 179, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Lotz, M.T.; Peebles, R.S. Mechanisms of Respiratory Syncytial Virus Modulation of Airway Immune Responses. Curr. Allergy Asthma Rep. 2012, 12, 380–387. [Google Scholar] [CrossRef] [Green Version]
- Jeyanathan, M.; Afkhami, S.; Smaill, F.; Miller, M.S.; Lichty, B.D.; Xing, Z. Immunological considerations for COVID-19 vaccine strategies. Nat. Rev. Immunol. 2020, 20, 615–632. [Google Scholar] [CrossRef]
- Sette, A.; Moutaftsi, M.; Moyron-Quiroz, J.; McCausland, M.M.; Davies, D.H.; Johnston, R.J.; Peters, B.; Benhnia, R.M.; Hoffmann, J.; Su, H.-P.; et al. Selective CD4 T cell help for antibody responses to a large viral pathogen: Deterministic linkage of specificities. Immunity 2008, 28, 847–858. [Google Scholar] [CrossRef] [Green Version]
- Arunachalam, P.S.; Charles, T.P.; Joag, V.; Bollimpelli, V.S.; Scott, M.K.D.; Wimmers, F.; Burton, S.L.; Labranche, C.C.; Petitdemange, C.; Gangadhara, S.; et al. T cell-inducing vaccine durably prevents mucosal SHIV infection even with lower neutralizing antibody titers. Nat. Med. 2020, 26, 932–940. [Google Scholar] [CrossRef]
- Oh, H.-L.J.; Gan, S.K.-E.; Bertoletti, A.; Tan, Y.-J. Understanding the T cell immune response in SARS coronavirus infection. Emerg. Microbes Infect. 2012, 1, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Spellberg, B.; Edwards, J.E. Type 1/Type 2 Immunity in Infectious Diseases. Clin. Infect. Dis. 2001, 32, 76–102. [Google Scholar] [CrossRef]
- Tseng, C.-T.; Sbrana, E.; Iwata-Yoshikawa, N.; Newman, P.C.; Garron, T.; Atmar, R.L.; Peters, C.J.; Couch, R.B. Immunization with SARS coronavirus vaccines leads to pulmonary immunopathology on challenge with the SARS virus. PLoS ONE 2012, 7, e35421. [Google Scholar] [CrossRef]
- Bolles, M.; Deming, D.; Long, K.; Agnihothram, S.; Whitmore, A.; Ferris, M.; Funkhouser, W.; Gralinski, L.; Totura, A.; Heise, M.; et al. A double-inactivated severe acute respiratory syndrome coronavirus vaccine provides incomplete protection in mice and induces increased eosinophilic proinflammatory pulmonary response upon challenge. J. Virol. 2011, 85, 12201–12215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDermott, D.S.; Weiss, K.A.; Knudson, C.J.; Varga, S.M. Central role of dendritic cells in shaping the adaptive immune response during respiratory syncytial virus infection. Future Virol. 2011, 6, 963–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, J.J.; Rudd, B.D.; Lukacs, N.W. Plasmacytoid dendritic cells inhibit pulmonary immunopathology and promote clearance of respiratory syncytial virus. J. Exp. Med. 2006, 203, 1153–1159. [Google Scholar] [CrossRef]
- Jang, S.; Smit, J.; Kallal, L.E.; Lukacs, N.W. Respiratory syncytial virus infection modifies and accelerates pulmonary disease via DC activation and migration. J. Leukoc. Biol. 2013, 94, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stetson, D.B.; Medzhitov, R. Type I interferons in host defense. Immunity 2006, 25, 373–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, P.L.; Fearns, R.; Graham, B.S. Respiratory Syncytial Virus: Virology, Reverse Genetics, and Pathogenesis of Disease. Curr. Top. Microbiol. Immunol. 2013, 372, 3–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, M.S.; Brazas, R.M.; Holtzman, M.J. Respiratory Syncytial Virus Nonstructural Proteins NS1 and NS2 Mediate Inhibition of Stat2 Expression and Alpha/Beta Interferon Responsiveness. J. Virol. 2005, 79, 9315–9319. [Google Scholar] [CrossRef] [Green Version]
- Bukreyev, A.; Yang, L.; Fricke, J.; Cheng, L.; Ward, J.M.; Murphy, B.R.; Collins, P.L. The Secreted Form of Respiratory Syncytial Virus G Glycoprotein Helps the Virus Evade Antibody-Mediated Restriction of Replication by Acting as an Antigen Decoy and through Effects on Fc Receptor-Bearing Leukocytes. J. Virol. 2008, 82, 12191–12204. [Google Scholar] [CrossRef] [Green Version]
- Gabriele, L.; Fragale, A.; Romagnoli, G.; Parlato, S.; Lapenta, C.; Santini, S.M.; Ozato, K.; Capone, I. Type I IFN-dependent antibody response at the basis of sex dimorphism in the outcome of COVID-19. Cytokine Growth Factor Rev. 2020. epublication, ahead of press. [Google Scholar] [CrossRef]
- Hadjadj, J.; Yatim, N.; Barnabei, L.; Corneau, A.; Boussier, J.; Smith, N.; Péré, H.; Charbit, B.; Bondet, V.; Chenevier-Gobeaux, C.; et al. Impaired type I interferon activity and inflammatory responses in severe COVID-19 patients. Science 2020, 369, 718–724. [Google Scholar] [CrossRef]
- Zhang, Q.; Bastard, P.; Liu, Z.; Pen, J.L.; Moncada-Velez, M.; Chen, J.; Ogishi, M.; Sabli, I.K.D.; Hodeib, S.; Korol, C.; et al. Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science 2020, 370, eabd4570. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Joosten, L.A.B.; Latz, E.; Mills, K.H.G.; Natoli, G.; Stunnenberg, H.G.; O’Neill, L.A.J.; Xavier, R.J. Trained immunity: A program of innate immune memory in health and disease. Science 2016, 352, aaf1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.M.; Colonna, M. Immune Training Unlocks Innate Potential. Cell 2018, 172, 3–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajishengallis, G.; Li, X.; Mitroulis, I.; Chavakis, T. Trained Innate Immunity and Its Implications for Mucosal Immunity and Inflammation. Adv. Exp. Med. Biol. 2019, 1197, 11–26. [Google Scholar] [CrossRef]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Joosten, L.A.B.; Ifrim, D.C.; Saeed, S.; Jacobs, C.; van Loenhout, J.; de Jong, D.; Stunnenberg, H.G.; et al. Bacille Calmette-Guerin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes. Proc. Natl. Acad. Sci. USA 2012, 109, 17537–17542. [Google Scholar] [CrossRef] [Green Version]
- Covián, C.; Fernández-Fierro, A.; Retamal-Díaz, A.; Díaz, F.E.; Vasquez, A.E.; Lay, M.K.; Riedel, C.A.; González, P.A.; Bueno, S.M.; Kalergis, A.M. BCG-Induced Cross-Protection and Development of Trained Immunity: Implication for Vaccine Design. Front. Immunol. 2019, 10, 2806. [Google Scholar] [CrossRef]
- Stensballe, L.G.; Nante, E.; Jensen, I.P.; Kofoed, P.-E.; Poulsen, A.; Jensen, H.; Newport, M.; Marchant, A.; Aaby, P. Acute lower respiratory tract infections and respiratory syncytial virus in infants in Guinea-Bissau: A beneficial effect of BCG vaccination for girls community based case-control study. Vaccine 2005, 23, 1251–1257. [Google Scholar] [CrossRef]
- Yitbarek, K.; Abraham, G.; Girma, T.; Tilahun, T.; Woldie, M. The effect of Bacillus Calmette–Guérin (BCG) vaccination in preventing severe infectious respiratory diseases other than TB: Implications for the COVID-19 pandemic. Vaccine 2020, 38, 6374–6380. [Google Scholar] [CrossRef]
- Mohapatra, P.R.; Mishra, B.; Behera, B. BCG vaccination induced protection from COVID-19. Indian J. Tuberc. 2020. epublication, ahead of press. [Google Scholar] [CrossRef]
- Rivas, M.N.; Ebinger, J.E.; Wu, M.; Sun, N.; Braun, J.; Sobhani, K.; Eyk, J.E.V.; Cheng, S.; Arditi, M. BCG vaccination history associates with decreased SARS-CoV-2 seroprevalence across a diverse cohort of healthcare workers. J. Clin. Investig. 2020. [Google Scholar] [CrossRef]
- Fidel, P.L.; Noverr, M.C. Could an Unrelated Live Attenuated Vaccine Serve as a Preventive Measure To Dampen Septic Inflammation Associated with COVID-19 Infection? mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Ashford, J.W.; Gold, J.E.; Huenergardt, M.A.; Katz, R.B.A.; Strand, S.E.; Bolanos, J.; Wheeler, C.J.; Perry, G.; Smith, C.J.; Steinman, L.; et al. MMR Vaccination: A Potential Strategy to Reduce Severity and Mortality of COVID-19 Illness. Am. J. Med. 2020. epublication, ahead of press. [Google Scholar] [CrossRef] [PubMed]
- Larenas-Linnemann, D.E.; Rodríguez-Monroy, F. Thirty-six COVID-19 cases preventively vaccinated with mumps-measles-rubella vaccine: All mild course. Allergy 2020. epublication, ahead of press. [Google Scholar] [CrossRef] [PubMed]
- Root-Bernstein, R. Age and Location in Severity of COVID-19 Pathology: Do Lactoferrin and Pneumococcal Vaccination Explain Low Infant Mortality and Regional Differences? BioEssays 2020, 42, 2000076. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Plata, A.; Kolli, D.; Hong, C.; Casola, A.; Garofalo, R.P. Subversion of Pulmonary Dendritic Cell Function by Paramyxovirus Infections. J. Immunol. 2009, 182, 3072–3083. [Google Scholar] [CrossRef] [Green Version]
- Didierlaurent, A.; Goulding, J.; Patel, S.; Snelgrove, R.; Low, L.; Bebien, M.; Lawrence, T.; van Rijt, L.S.; Lambrecht, B.N.; Sirard, J.-C.; et al. Sustained desensitization to bacterial Toll-like receptor ligands after resolutionof respiratory influenza infection. J. Exp. Med. 2008, 205, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Low, J.S.; Farsakoglu, Y.; Amezcua Vesely, M.C.; Sefik, E.; Kelly, J.B.; Harman, C.C.D.; Jackson, R.; Shyer, J.A.; Jiang, X.; Cauley, L.S.; et al. Tissue-resident memory T cell reactivation by diverse antigen-presenting cells imparts distinct functional responses. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- Gilbert, S.C. T-cell-inducing vaccines—What’s the future. Immunology 2012, 135, 19–26. [Google Scholar] [CrossRef]
- Gordon, S.N.; Cecchinato, V.; Andresen, V.; Heraud, J.-M.; Hryniewicz, A.; Parks, R.W.; Venzon, D.; Chung, H.; Karpova, T.; McNally, J.; et al. Smallpox vaccine safety is dependent on T cells and not B cells. J. Infect. Dis. 2011, 203, 1043–1053. [Google Scholar] [CrossRef] [Green Version]
- Johnson, N.; Cunningham, A.F.; Fooks, A.R. The immune response to rabies virus infection and vaccination. Vaccine 2010, 28, 3896–3901. [Google Scholar] [CrossRef]
- Elsayed, H.; Nabi, G.; McKinstry, W.J.; Khoo, K.K.; Mak, J.; Salazar, A.M.; Tenbusch, M.; Temchura, V.; Überla, K. Intrastructural Help: Harnessing T-Helper Cells Induced by Licensed Vaccines for Improvement of HIV Env Antibody Responses to Virus-Like Particle Vaccines. J. Virol. 2018, 92, e00141-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMichael, A.J.; Koff, W.C. Vaccines that stimulate T cell immunity to HIV-1: The next step. Nat. Immunol. 2014, 15, 319–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moris, P.; Jongert, E.; van der Most, R.G. Characterization of T-cell immune responses in clinical trials of the candidate RTS, S malaria vaccine. Hum. Vaccines Immunother. 2018, 14, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nlinwe, O.N.; Kusi, K.A.; Adu, B.; Sedegah, M. T-cell responses against Malaria: Effect of parasite antigen diversity and relevance for vaccine development. Vaccine 2018, 36, 2237–2242. [Google Scholar] [CrossRef]
- Marinaik, C.B.; Kingstad-Bakke, B.; Lee, W.; Hatta, M.; Sonsalla, M.; Larsen, A.; Neldner, B.; Gasper, D.J.; Kedl, R.M.; Kawaoka, Y.; et al. Programming Multifaceted Pulmonary T Cell Immunity by Combination Adjuvants. Cell Rep. Med. 2020, 1, 100095. [Google Scholar] [CrossRef]
- Berthoud, T.K.; Hamill, M.; Lillie, P.J.; Hwenda, L.; Collins, K.A.; Ewer, K.J.; Milicic, A.; Poyntz, H.C.; Lambe, T.; Fletcher, H.A.; et al. Potent CD8+ T-cell immunogenicity in humans of a novel heterosubtypic influenza A vaccine, MVA-NP+M1. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2011, 52, 1–7. [Google Scholar] [CrossRef]
- Stoloff, G.A.; Caparros-Wanderley, W. Synthetic multi-epitope peptides identified in silico induce protective immunity against multiple influenza serotypes. Eur. J. Immunol. 2007, 37, 2441–2449. [Google Scholar] [CrossRef]
- Francis, J.N.; Bunce, C.J.; Horlock, C.; Watson, J.M.; Warrington, S.J.; Georges, B.; Brown, C.B. A novel peptide-based pan-influenza A vaccine: A double blind, randomised clinical trial of immunogenicity and safety. Vaccine 2015, 33, 396–402. [Google Scholar] [CrossRef] [Green Version]
- Atsmon, J.; Kate-Ilovitz, E.; Shaikevich, D.; Singer, Y.; Volokhov, I.; Haim, K.Y.; Ben-Yedidia, T. Safety and immunogenicity of multimeric-001--a novel universal influenza vaccine. J. Clin. Immunol. 2012, 32, 595–603. [Google Scholar] [CrossRef]
- Mateus, J.; Grifoni, A.; Tarke, A.; Sidney, J.; Ramirez, S.I.; Dan, J.M.; Burger, Z.C.; Rawlings, S.A.; Smith, D.M.; Phillips, E.; et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 2020, 370, 89–94. [Google Scholar] [CrossRef]
- Ni, L.; Ye, F.; Cheng, M.-L.; Feng, Y.; Deng, Y.-Q.; Zhao, H.; Wei, P.; Ge, J.; Gou, M.; Li, X.; et al. Detection of SARS-CoV-2-Specific Humoral and Cellular Immunity in COVID-19 Convalescent Individuals. Immunity 2020, 52, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Liu, Y.; Yuan, J.; Wen, Y.; Xu, G.; Zhao, J.; Cheng, L.; Li, J.; Wang, X.; Wang, F.; et al. Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19. Nat. Med. 2020, 26, 842–844. [Google Scholar] [CrossRef]
- Kosanovich, J.L.; Eichinger, K.M.; Lipp, M.A.; Yondola, M.A.; Perkins, T.N.; Empey, K.M. Formulation of the prefusion RSV F protein with a Th1/Th2-balanced adjuvant provides complete protection without Th2-skewed immunity in RSV-experienced young mice. Vaccine 2020, 38, 6357–6362. [Google Scholar] [CrossRef] [PubMed]
- Malinczak, C.-A.; Rasky, A.J.; Fonseca, W.; Schaller, M.A.; Allen, R.M.; Ptaschinski, C.; Morris, S.; Lukacs, N.W. Upregulation of H3K27 Demethylase KDM6 During Respiratory Syncytial Virus Infection Enhances Proinflammatory Responses and Immunopathology. J. Immunol. 2019, 204, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Ptaschinski, C.; Mukherjee, S.; Moore, M.L.; Albert, M.; Helin, K.; Kunkel, S.L.; Lukacs, N.W. RSV-Induced H3K4 Demethylase KDM5B Leads to Regulation of Dendritic Cell-Derived Innate Cytokines and Exacerbates Pathogenesis In Vivo. PLoS Pathog. 2015, 11, e1004978. [Google Scholar] [CrossRef] [PubMed]
- Duthie, M.S.; Windish, H.P.; Fox, C.B.; Reed, S.G. Use of defined TLR ligands as adjuvants within human vaccines. Immunol. Rev. 2011, 239, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Gupta, T.; Gupta, S.K. Potential adjuvants for the development of a SARS-CoV-2 vaccine based on experimental results from similar coronaviruses. Int. Immunopharmacol. 2020, 86, 106717. [Google Scholar] [CrossRef]
- Kamphuis, T.; Shafique, M.; Meijerhof, T.; Stegmann, T.; Wilschut, J.; de Haan, A. Efficacy and safety of an intranasal virosomal respiratory syncytial virus vaccine adjuvanted with monophosphoryl lipid A in mice and cotton rats. Vaccine 2013, 31, 2169–2176. [Google Scholar] [CrossRef]
- Shafique, M.; Meijerhof, T.; Wilschut, J.; de Haan, A. Evaluation of an intranasal virosomal vaccine against respiratory syncytial virus in mice: Effect of TLR2 and NOD2 ligands on induction of systemic and mucosal immune responses. PLoS ONE 2013, 8, e61287. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.R.; Rao, S.; Seder, R.A.; Chen, M.; Graham, B.S. TLR9 Agonist, but Not TLR7/8, Functions As An Adjuvant To Diminish FI-RSV Vaccine-Enhanced Disease, While Either Agonist Used as Therapy During Primary RSV Infection Increases Disease Severity. Vaccine 2009, 27, 3045–3052. [Google Scholar] [CrossRef] [Green Version]
- Dowling, D.J. Recent Advances in the Discovery and Delivery of TLR7/8 Agonists as Vaccine Adjuvants. ImmunoHorizons 2018, 2, 185–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwata-Yoshikawa, N.; Uda, A.; Suzuki, T.; Tsunetsugu-Yokota, Y.; Sato, Y.; Morikawa, S.; Tashiro, M.; Sata, T.; Hasegawa, H.; Nagata, N. Effects of Toll-Like Receptor Stimulation on Eosinophilic Infiltration in Lungs of BALB/c Mice Immunized with UV-Inactivated Severe Acute Respiratory Syndrome-Related Coronavirus Vaccine. J. Virol. 2014, 88, 8597–8614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Wohlford-Lenane, C.; Zhao, J.; Fleming, E.; Lane, T.E.; McCray, P.B.; Perlman, S. Intranasal Treatment with Poly(I·C) Protects Aged Mice from Lethal Respiratory Virus Infections. J. Virol. 2012, 86, 11416–11424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gai, W.; Zou, W.; Lei, L.; Luo, J.; Tu, H.; Zhang, Y.; Wang, K.; Tien, P.; Yan, H. Effects of different immunization protocols and adjuvant on antibody responses to inactivated SARS-CoV vaccine. Viral Immunol. 2008, 21, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, K.; Wang, H.; Wu, C. The immune responses of HLA-A*0201 restricted SARS-CoV S peptide-specific CD8+ T cells are augmented in varying degrees by CpG ODN, PolyI:C and R848. Vaccine 2011, 29, 6670–6678. [Google Scholar] [CrossRef] [PubMed]
- Ausiello, C.M.; Cassone, A. Acellular Pertussis Vaccines and Pertussis Resurgence: Revise or Replace? mBio 2014, 5, e01339-14. [Google Scholar] [CrossRef] [Green Version]
- Al-Halifa, S.; Gauthier, L.; Arpin, D.; Bourgault, S.; Archambault, D. Nanoparticle-Based Vaccines Against Respiratory Viruses. Front. Immunol. 2019, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- Pachioni-Vasconcelos, J.d.A.; Lopes, A.M.; Apolinário, A.C.; Valenzuela-Oses, J.K.; Costa, J.S.R.; Nascimento, L.d.O.; Pessoa, A.; Barbosa, L.R.S.; Rangel-Yagui, C.D.O. Nanostructures for protein drug delivery. Biomater. Sci. 2016, 4, 205–218. [Google Scholar] [CrossRef]
- Lindell, D.M.; Morris, S.B.; White, M.P.; Kallal, L.E.; Lundy, P.K.; Hamouda, T.; Baker, J.R., Jr.; Lukacs, N.W. A Novel Inactivated Intranasal Respiratory Syncytial Virus Vaccine Promotes Viral Clearance without Th2 Associated Vaccine-Enhanced Disease. PLoS ONE 2011, 6, e21823. [Google Scholar] [CrossRef]
- Bielinska, A.U.; Makidon, P.E.; Janczak, K.W.; Blanco, L.P.; Swanson, B.; Smith, D.M.; Pham, T.; Szabo, Z.; Kukowska-Latallo, J.F.; Baker, J.R. Distinct Pathways of Humoral and Cellular Immunity Induced with the Mucosal Administration of a Nanoemulsion Adjuvant. J. Immunol. 2014, 192, 2722–2733. [Google Scholar] [CrossRef] [Green Version]
- O’Konek, J.J.; Makidon, P.E.; Landers, J.J.; Cao, Z.; Malinczak, C.-A.; Pannu, J.; Sun, J.; Bitko, V.; Ciotti, S.; Hamouda, T.; et al. Intranasal nanoemulsion-based inactivated respiratory syncytial virus vaccines protect against viral challenge in cotton rats. Hum. Vaccines Immunother. 2015, 11, 2904–2912. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, M.J.; Lyke, K.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Raabe, V.; Bailey, R.; Swanson, K.A.; et al. Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature 2020, 586, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and T H 1 T cell responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.J.; Rouphael, N.G.; Widge, A.T.; Jackson, L.A.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Safety and Immunogenicity of SARS-CoV-2 mRNA-1273 Vaccine in Older Adults. N. Engl. J. Med. 2020. epublication, ahead of press. [Google Scholar] [CrossRef]
- Walls, A.C.; Fiala, B.; Schäfer, A.; Wrenn, S.; Pham, M.N.; Murphy, M.; Tse, L.V.; Shehata, L.; O’Connor, M.A.; Chen, C.; et al. Elicitation of Potent Neutralizing Antibody Responses by Designed Protein Nanoparticle Vaccines for SARS-CoV-2. Cell 2020, 183, 1367–1382.e17. [Google Scholar] [CrossRef]
- Corbett, K.S.; Flynn, B.; Foulds, K.E.; Francica, J.R.; Boyoglu-Barnum, S.; Werner, A.P.; Flach, B.; O’Connell, S.; Bock, K.W.; Minai, M.; et al. Evaluation of the mRNA-1273 Vaccine against SARS-CoV-2 in Nonhuman Primates. N. Engl. J. Med. 2020, 383, 1544–1555. [Google Scholar] [CrossRef]
- Callaway, E. What Pfizer’s landmark COVID vaccine results mean for the pandemic. Nature 2020. Available online: https://www.nature.com/articles/d41586-020-03166-8 (accessed on 15 December 2020). [CrossRef]
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An mRNA Vaccine against SARS-CoV-2—Preliminary Report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef]
- Roldão, A.; Mellado, M.C.M.; Castilho, L.R.; Carrondo, M.J.T.; Alves, P.M. Virus-like particles in vaccine development. Expert Rev. Vaccines 2010, 9, 1149–1176. [Google Scholar] [CrossRef]
- Kim, A.-R.; Lee, D.-H.; Lee, S.-H.; Rubino, I.; Choi, H.-J.; Quan, F.-S. Protection induced by virus-like particle vaccine containing tandem repeat gene of respiratory syncytial virus G protein. PLoS ONE 2018, 13, e0191277. [Google Scholar] [CrossRef] [Green Version]
- Quan, F.-S.; Kim, Y.; Lee, S.; Yi, H.; Kang, S.-M.; Bozja, J.; Moore, M.L.; Compans, R.W. Viruslike Particle Vaccine Induces Protection Against Respiratory Syncytial Virus Infection in Mice. J. Infect. Dis. 2011, 204, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-H.; Lee, Y.-T.; Hwang, H.S.; Kwon, Y.-M.; Kim, M.-C.; Ko, E.-J.; Lee, J.S.; Lee, Y.; Kang, S.-M. Virus-Like Particle Vaccine Containing the F Protein of Respiratory Syncytial Virus Confers Protection without Pulmonary Disease by Modulating Specific Subsets of Dendritic Cells and Effector T Cells. J. Virol. 2015, 89, 11692–11705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Lu, G.; Tan, S.; Xia, J.; Xiong, H.; Yu, X.; Qi, Q.; Yu, X.; Li, L.; Yu, H.; et al. A COVID-19 mRNA vaccine encoding SARS-CoV-2 virus-like particles induces a strong antiviral-like immune response in mice. Cell Res. 2020, 30, 936–939. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Shi, M.; Li, J.; Song, P.; Li, N. Construction of SARS-CoV-2 Virus-Like Particles by Mammalian Expression System. Front. Bioeng. Biotechnol. 2020, 8, 862. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Huang, Y.; Huang, L.; Li, B.; Zheng, Z.; Chen, Z.; Chen, J.; Hu, Q.; Wang, H. Effect of mucosal and systemic immunization with virus-like particles of severe acute respiratory syndrome coronavirus in mice. Immunology 2010, 130, 254–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Zheng, X.; Gai, W.; Wong, G.; Wang, H.; Jin, H.; Feng, N.; Zhao, Y.; Zhang, W.; Li, N.; et al. Novel chimeric virus-like particles vaccine displaying MERS-CoV receptor-binding domain induce specific humoral and cellular immune response in mice. Antiviral Res. 2017, 140, 55–61. [Google Scholar] [CrossRef]
- Ewer, K.; Sebastian, S.; Spencer, A.J.; Gilbert, S.; Hill, A.V.S.; Lambe, T. Chimpanzee adenoviral vectors as vaccines for outbreak pathogens. Hum. Vaccines Immunother. 2017, 13, 3020–3032. [Google Scholar] [CrossRef]
- Zhang, C.; Zhou, D. Adenoviral vector-based strategies against infectious disease and cancer. Hum. Vaccines Immunother. 2016, 12, 2064–2074. [Google Scholar] [CrossRef] [Green Version]
- Hassan, A.O.; Kafai, N.M.; Dmitriev, I.P.; Fox, J.M.; Smith, B.K.; Harvey, I.B.; Chen, R.E.; Winkler, E.S.; Wessel, A.W.; Case, J.B.; et al. A Single-Dose Intranasal ChAd Vaccine Protects Upper and Lower Respiratory Tracts against SARS-CoV-2. Cell 2020, 183, 169–184.e13. [Google Scholar] [CrossRef]
- Folegatti, P.M.; Ewer, K.J.; Aley, P.K.; Angus, B.; Becker, S.; Belij-Rammerstorfer, S.; Bellamy, D.; Bibi, S.; Bittaye, M.; Clutterbuck, E.A.; et al. Safety and immunogenicity of the ChAdOx1 nCoV-19 vaccine against SARS-CoV-2: A preliminary report of a phase 1/2, single-blind, randomised controlled trial. Lancet Lond. Engl. 2020, 396, 467–478. [Google Scholar] [CrossRef]
- Sadoff, J.; Gars, M.L.; Shukarev, G.; Heerwegh, D.; Truyers, C.; de Groot, A.M.; Stoop, J.; Tete, S.; Damme, W.V.; Leroux-Roels, I.; et al. Safety and immunogenicity of the Ad26.COV2.S COVID-19 vaccine candidate: Interim results of a phase 1/2a, double-blind, randomized, placebo-controlled trial. medRxiv 2020. preprint. [Google Scholar] [CrossRef]
- Zhu, F.-C.; Guan, X.-H.; Li, Y.-H.; Huang, J.-Y.; Jiang, T.; Hou, L.-H.; Li, J.-X.; Yang, B.-F.; Wang, L.; Wang, W.-J.; et al. Immunogenicity and safety of a recombinant adenovirus type-5-vectored COVID-19 vaccine in healthy adults aged 18 years or older: A randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2020, 396, 479–488. [Google Scholar] [CrossRef]
- Logunov, D.Y.; Dolzhikova, I.V.; Zubkova, O.V.; Tukhvatullin, A.I.; Shcheblyakov, D.V.; Dzharullaeva, A.S.; Grousova, D.M.; Erokhova, A.S.; Kovyrshina, A.V.; Botikov, A.G.; et al. Safety and immunogenicity of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine in two formulations: Two open, non-randomised phase 1/2 studies from Russia. Lancet 2020, 396, 887–897. [Google Scholar] [CrossRef]
- Widjojoatmodjo, M.N.; Bogaert, L.; Meek, B.; Zahn, R.; Vellinga, J.; Custers, J.; Serroyen, J.; Radošević, K.; Schuitemaker, H. Recombinant low-seroprevalent adenoviral vectors Ad26 and Ad35 expressing the respiratory syncytial virus (RSV) fusion protein induce protective immunity against RSV infection in cotton rats. Vaccine 2015, 33, 5406–5414. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.R.; Rangel, D.; Graham, B.S.; Brough, D.E.; Gall, J.G. Genetic vaccine for respiratory syncytial virus provides protection without disease potentiation. Mol. Ther. J. Am. Soc. Gene Ther. 2014, 22, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Pierantoni, A.; Esposito, M.L.; Ammendola, V.; Napolitano, F.; Grazioli, F.; Abbate, A.; Del Sorbo, M.; Siani, L.; D’Alise, A.M.; Taglioni, A.; et al. Mucosal delivery of a vectored RSV vaccine is safe and elicits protective immunity in rodents and nonhuman primates. Mol. Ther. Methods Clin. Dev. 2015, 2, 15018. [Google Scholar] [CrossRef]
- Taylor, G.; Thom, M.; Capone, S.; Pierantoni, A.; Guzman, E.; Herbert, R.; Scarselli, E.; Napolitano, F.; Giuliani, A.; Folgori, A.; et al. Efficacy of a virus-vectored vaccine against human and bovine respiratory syncytial virus infections. Sci. Transl. Med. 2015, 7, 300ra127. [Google Scholar] [CrossRef]
- Green, C.A.; Scarselli, E.; Sande, C.J.; Thompson, A.J.; de Lara, C.M.; Taylor, K.S.; Haworth, K.; Del Sorbo, M.; Angus, B.; Siani, L.; et al. Chimpanzee adenovirus- and MVA-vectored respiratory syncytial virus vaccine is safe and immunogenic in adults. Sci. Transl. Med. 2015, 7, 300ra126. [Google Scholar] [CrossRef] [Green Version]
- Cicconi, P.; Jones, C.; Sarkar, E.; Silva-Reyes, L.; Klenerman, P.; de Lara, C.; Hutchings, C.; Moris, P.; Janssens, M.; Fissette, L.A.; et al. First-in-Human Randomized Study to Assess the Safety and Immunogenicity of an Investigational Respiratory Syncytial Virus (RSV) Vaccine Based on Chimpanzee-Adenovirus-155 Viral Vector–Expressing RSV Fusion, Nucleocapsid, and Antitermination Viral Proteins in Healthy Adults. Clin. Infect. Dis. 2020, 70, 2073–2081. [Google Scholar] [CrossRef]
- Sayedahmed, E.E.; Elkashif, A.; Alhashimi, M.; Sambhara, S.; Mittal, S.K. Adenoviral Vector-Based Vaccine Platforms for Developing the Next Generation of Influenza Vaccines. Vaccines 2020, 8, 574. [Google Scholar] [CrossRef]
- Gao, W.; Soloff, A.C.; Lu, X.; Montecalvo, A.; Nguyen, D.C.; Matsuoka, Y.; Robbins, P.D.; Swayne, D.E.; Donis, R.O.; Katz, J.M.; et al. Protection of mice and poultry from lethal H5N1 avian influenza virus through adenovirus-based immunization. J. Virol. 2006, 80, 1959–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoelscher, M.A.; Garg, S.; Bangari, D.S.; Belser, J.A.; Lu, X.; Stephenson, I.; Bright, R.A.; Katz, J.M.; Mittal, S.K.; Sambhara, S. Development of adenoviral-vector-based pandemic influenza vaccine against antigenically distinct human H5N1 strains in mice. Lancet Lond. Engl. 2006, 367, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Philbin, V.J.; Levy, O. Developmental Biology of the Innate Immune Response: Implications for Neonatal and Infant Vaccine Development. Pediatr. Res. 2009, 65, 98–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, A.K.; Hollander, G.A.; McMichael, A. Evolution of the immune system in humans from infancy to old age. Proc. R. Soc. B Biol. Sci. 2015, 282, 20143085. [Google Scholar] [CrossRef]
- Kollmann, T.R.; Kampmann, B.; Mazmanian, S.K.; Marchant, A.; Levy, O. Protecting the Newborn and Young Infant from Infectious Diseases: Lessons from Immune Ontogeny. Immunity 2017, 46, 350–363. [Google Scholar] [CrossRef] [Green Version]
- Sweet, C.; Bird, R.A.; Jakeman, K.; Coates, D.M.; Smith, H. Production of passive immunity in neonatal ferrets following maternal vaccination with killed influenza A virus vaccines. Immunology 1987, 60, 83–89. [Google Scholar]
- Blanco, J.C.G.; Pletneva, L.M.; McGinnes-Cullen, L.; Otoa, R.O.; Patel, M.C.; Fernando, L.R.; Boukhvalova, M.S.; Morrison, T.G. Efficacy of a respiratory syncytial virus vaccine candidate in a maternal immunization model. Nat. Commun. 2018, 9, 1904. [Google Scholar] [CrossRef]
- Blanco, J.C.G.; Pletneva, L.M.; Otoa, R.O.; Patel, M.C.; Vogel, S.N.; Boukhvalova, M.S. Preclinical assessment of safety of maternal vaccination against respiratory syncytial virus (RSV) in cotton rats. Vaccine 2017, 35, 3951–3958. [Google Scholar] [CrossRef]
- Madhi, S.A.; Polack, F.P.; Piedra, P.A.; Munoz, F.M.; Trenholme, A.A.; Simões, E.A.F.; Swamy, G.K.; Agrawal, S.; Ahmed, K.; August, A.; et al. Respiratory Syncytial Virus Vaccination during Pregnancy and Effects in Infants. N. Engl. J. Med. 2020, 383, 426–439. [Google Scholar] [CrossRef]
- Poehling, K.A.; Szilagyi, P.G.; Staat, M.A.; Snively, B.M.; Payne, D.C.; Bridges, C.B.; Chu, S.Y.; Light, L.S.; Prill, M.M.; Finelli, L.; et al. Impact of maternal immunization on influenza hospitalizations in infants. Am. J. Obstet. Gynecol. 2011, 204, S141–S148. [Google Scholar] [CrossRef] [Green Version]
- García, C.G.; Bhore, R.; Soriano-Fallas, A.; Trost, M.; Chason, R.; Ramilo, O.; Mejias, A. Risk factors in children hospitalized with RSV bronchiolitis versus non-RSV bronchiolitis. Pediatrics 2010, 126, e1453–e1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommer, C.; Resch, B.; Simões, E.A.F. Risk Factors for Severe Respiratory Syncytial Virus Lower Respiratory Tract Infection. Open Microbiol. J. 2011, 5, 144–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| General Vaccine Strategy | Antigen (Respiratory Virus Target) | Candidate Vaccine Examples | Induced Immune Response | Review Location |
|---|---|---|---|---|
| T cell epitope | MVA vector encoding NP, M1 subunit (Influenza) | MVA-NP+M1 | CD8+ IFN-γ-producing T cells | Section 4.1 |
| Multiple Influenza T cell epitopes (Influenza) | Preclinical | HLA-A*0201 Cross-reactive CD8+ T cell responses | Section 4.1 | |
| Multiple CD4+/CD8+ T cell epitopes (Influenza) | FP-01.1 | Dual CD4+/CD8+ T cell responses and vaccine-specific T cells that cross react with multiple divergent influenza strains | Section 4.1 | |
| HA, NP, M1 protein subunit (Influenza) | Multimeric-001 | Th1/IFN-γ-driven protection against H1N1, H3N2, and influenza B | Section 4.1 | |
| TLR-adjuvant | Virosomes + TLR4 (RSV) | Preclinical | Th1 response without Th2 skewing | Section 4.2 |
| Virosomes + TLR2 (RSV) | Preclinical | Activated APC and Th1 response without Th2 skewing | Section 4.2 | |
| Formalin-inactivated RSV + TLR9 (RSV) | Preclinical | Increased Th1 cytokine response with decreased Th2; protection from vaccine enhanced disease | Section 4.2 | |
| UV-inactivated SARS-CoV + TLR3/4 (SARS-CoV) | Preclinical | Reduction of immunopathogenic Th2 responses | Section 4.2 | |
| TLR3 pretreatment (Influenza and SARS-CoV) | Preclinical | Upregulation of IFN-β and IFN-γ production | Section 4.2 | |
| SARS-CoV S peptide subunit + TLR9 (SARS-CoV) | Preclinical | Induction of IFN-γ-producing CD8+ memory T cells | Section 4.2 | |
| Conventional | Live-Attenuated (Multiple) | Multiple | Highly immunogenic but may lead to pathogenic immune responses | Section 4.3 |
| Inactivated Whole Virus (Multiple) | Multiple | Weak immune response without the addition of adjuvant | Section 4.3 | |
| Subunit (Multiple) | Multiple | Limited immunogenicity without proper adjuvancy or packaging (i.e., nanoparticle, virus-like or live viral vectors) | Section 4.3 | |
| Nanoparticle | mRNA/ RBD spike (S) protein subunit (SARS-CoV-2) | BNT162 | CD4+/CD8+ IFN-γ-producing T cells | Section 4.3.1 |
| mRNA/ pre-fusion S-2P (SARS-CoV-2) | mRNA-1273 | Strong CD4+ Th1 cell response, low level CD8+ T cell response | Section 4.3.1 | |
| Virus-like Particles (VLP) | RSV-F subunit (RSV) | Preclinical | Induces CD8+ and CD103+ DC and F-specific IFN-γ/TNF-α CD8+ T cells | Section 4.3.2 |
| mRNA/ S, M (membrane), and E (envelope) subunit (SARS-CoV-2) | Preclinical | CD4+/CD8+ IFN-γ-producing T cells | Section 4.3.2 | |
| Chimeric/ MERS RBD and CPV + TLR3 adjuvant (MERS-CoV) | Preclinical | DC activation and Th1/IFN-γ, Th2/IL-4 production by splenocytes | Section 4.3.2 | |
| Adenoviral Vector | S protein (SARS-CoV-2) | ChAdOx1 nCov-19 | Th1/INF-γ-producing CD4+/CD8+ T cells with no Th2-skewing | Section 4.3.3 |
| Pre-fusion S protein (SARS-CoV-2) | Ad26.COV2.S | Th1/INF-γ-producing CD4+/CD8+ T cells with no Th2-skewing | Section 4.3.3 | |
| RBD subunit (SARS-CoV-2) | Ad5-COVID-19 | Th1/INF-γ-producing CD4+/CD8+ T cells with no Th2-skewing | Section 4.3.3 | |
| RSV F fusion subunit (RSV) | Preclinical Human Ad26/35 | F-specific IFN-γ producing T cells | Section 4.3.3 | |
| RSV F fusion subunit (RSV) | Preclinical Gorilla AdV | RSV-specific CD4+/CD8+ Th1 cells with limited Th2 responses | Section 4.3.3 | |
| RSV F, N, and M2-1 subunit (RSV) | PanAd3-RSV Chimpanzee AdV | CD4+/CD8+ IFN-γ-producing T cells without Th2 priming | Section 4.3.3 | |
| RSV F fusion (RSV) | ChAd-155-RSV F | CD4+/CD8+ IFN-γ-producing T cells without Th2 priming | Section 4.3.3 | |
| Numerous HA, NP, and M2 protein subunits (Influenza) | Preclinical Human AdV5, ChAd, and bovine AdV Clinical: Multiple Human AdV | Broad humoral and cellular immune responses | Section 4.3.3 | |
| Maternal Vaccination | Formalin-inactivated influenza A (Influenza) | Preclinical | Limited humoral response in neonatal offspring | Section 4.3.4 |
| RSV F and RSV G protein subunit VLPs (RSV) | Preclinical | Neonatal protection with reduced vaccine enhanced disease | Section 4.3.4 | |
| Formalin-inactivated RSV (RSV) | Preclinical | Enhanced disease of offspring upon live viral challenge | Section 4.3.4 | |
| RSV-F nanoparticle subunit (RSV) | Phase III clinical trial | Decreased LRTI and hospitalization in infants born to vaccinated mothers | Section 4.3.4 | |
| Influenza vaccination | Population-based study | Infants born to vaccinated mothers 45–48% less likely to become hospitalized | Section 4.3.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lukacs, N.W.; Malinczak, C.-A. Harnessing Cellular Immunity for Vaccination against Respiratory Viruses. Vaccines 2020, 8, 783. https://doi.org/10.3390/vaccines8040783
Lukacs NW, Malinczak C-A. Harnessing Cellular Immunity for Vaccination against Respiratory Viruses. Vaccines. 2020; 8(4):783. https://doi.org/10.3390/vaccines8040783
Chicago/Turabian StyleLukacs, Nicholas W., and Carrie-Anne Malinczak. 2020. "Harnessing Cellular Immunity for Vaccination against Respiratory Viruses" Vaccines 8, no. 4: 783. https://doi.org/10.3390/vaccines8040783