A Comparative Study of Pathology and Host Immune Response Induced by Very Virulent Infectious Bursal Disease Virus in Experimentally Infected Chickens of Aseel and White Leghorn Breeds

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Virus
2.2. Experimental Birds
2.3. Experimental Design
2.4. Histopathology
2.5. Immunohistochemistry
2.6. RNA Preparation
2.7. Quantitative Real-Time RT-PCR
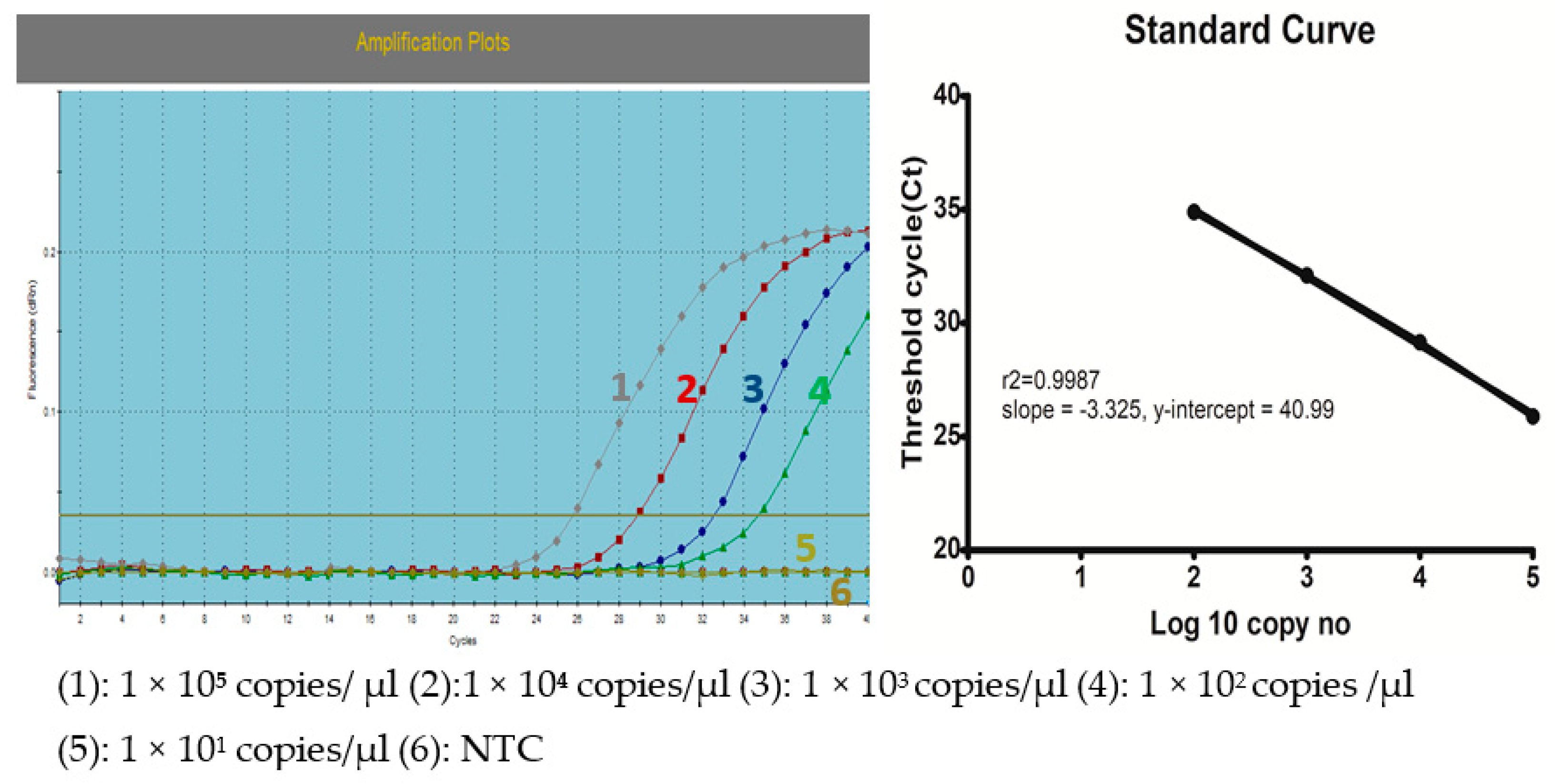
2.8. Determination of Viral Load and Preparation of a Standard Curve
2.9. Statistical Analyses
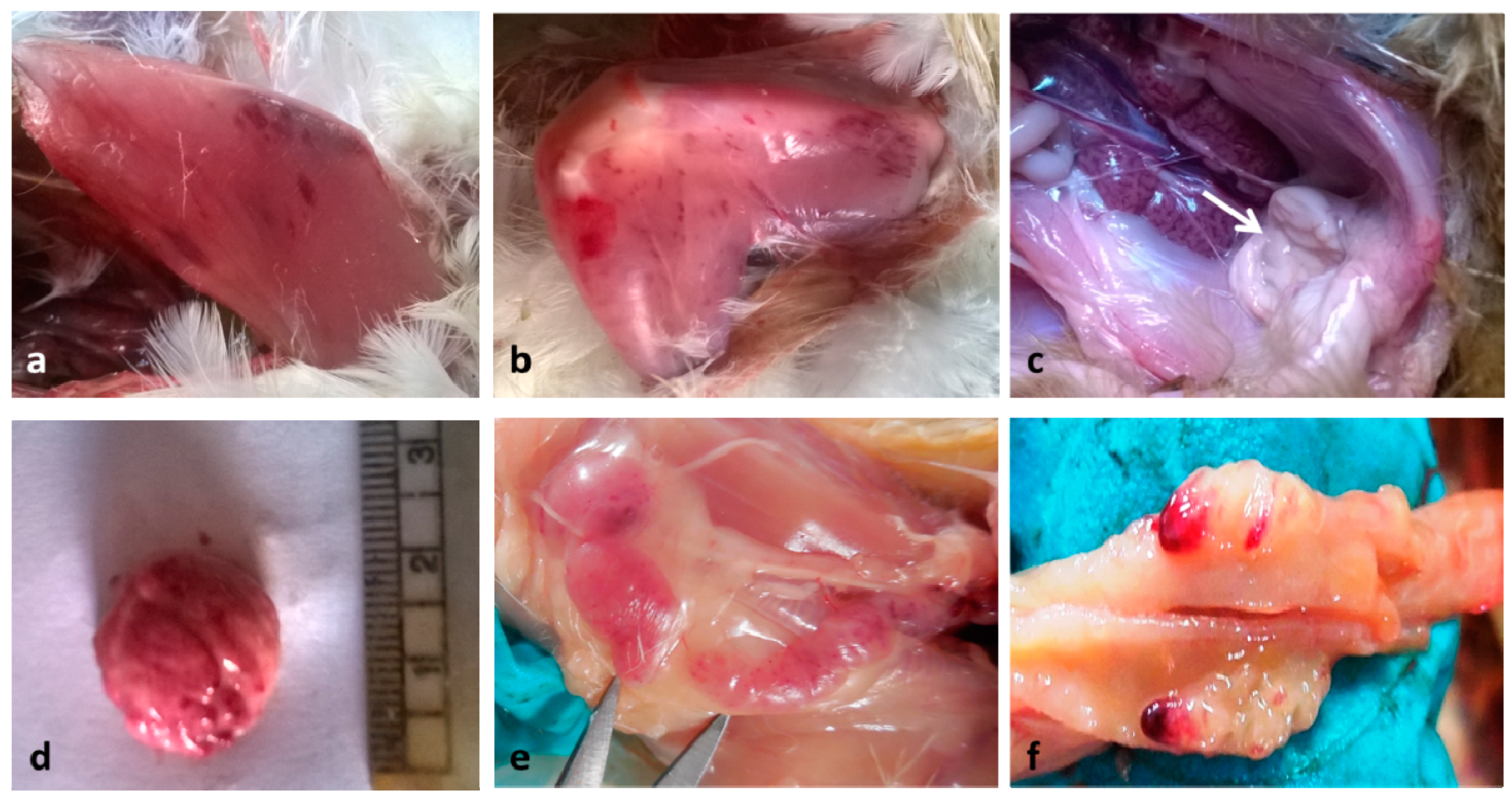
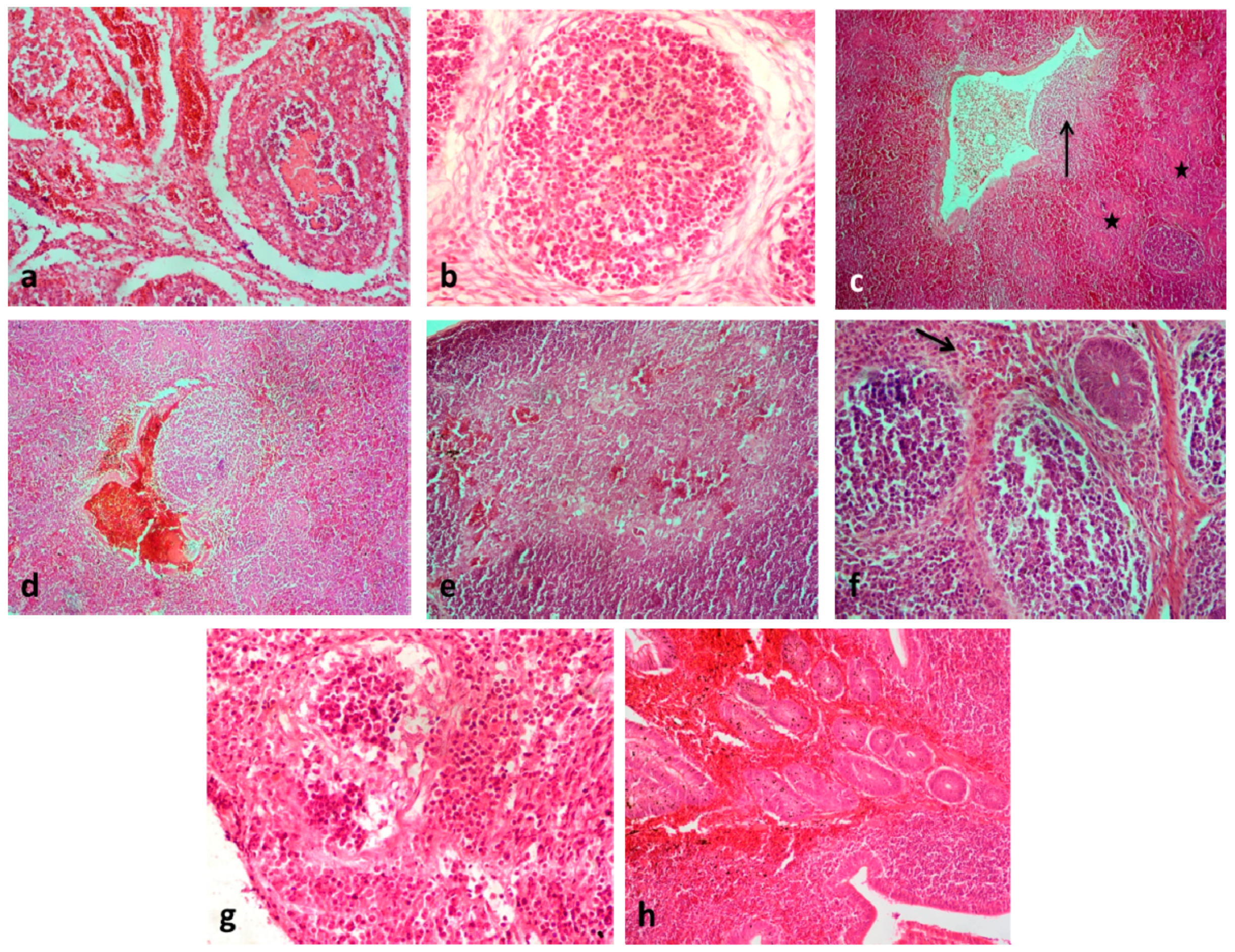
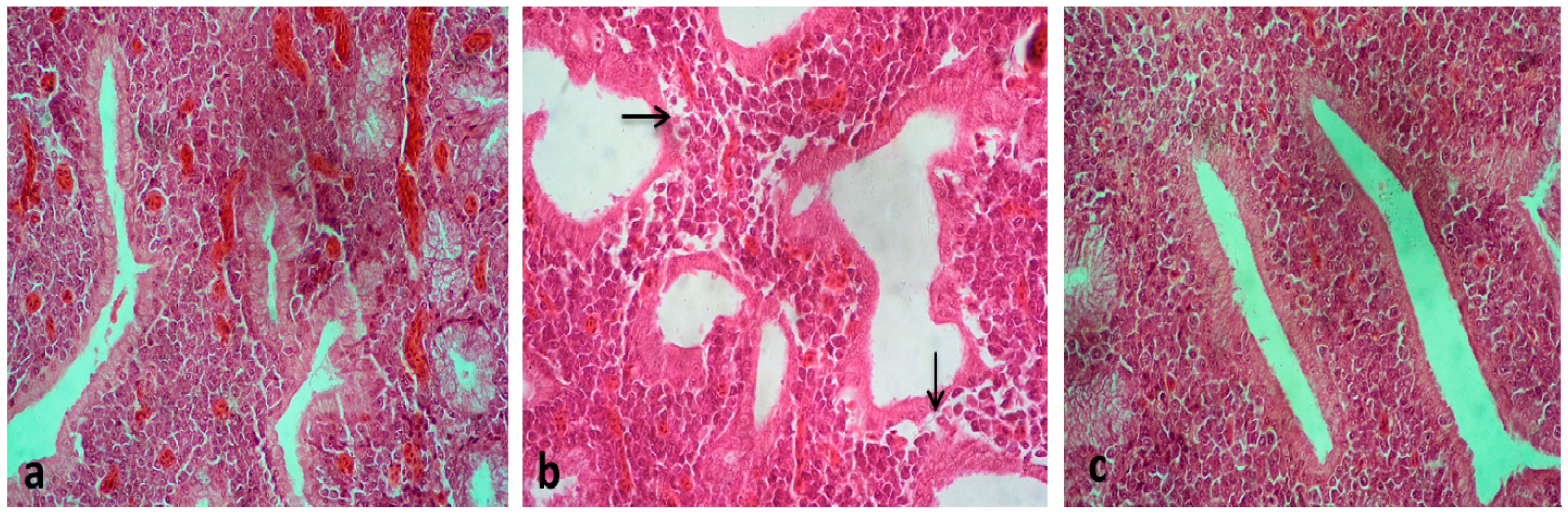
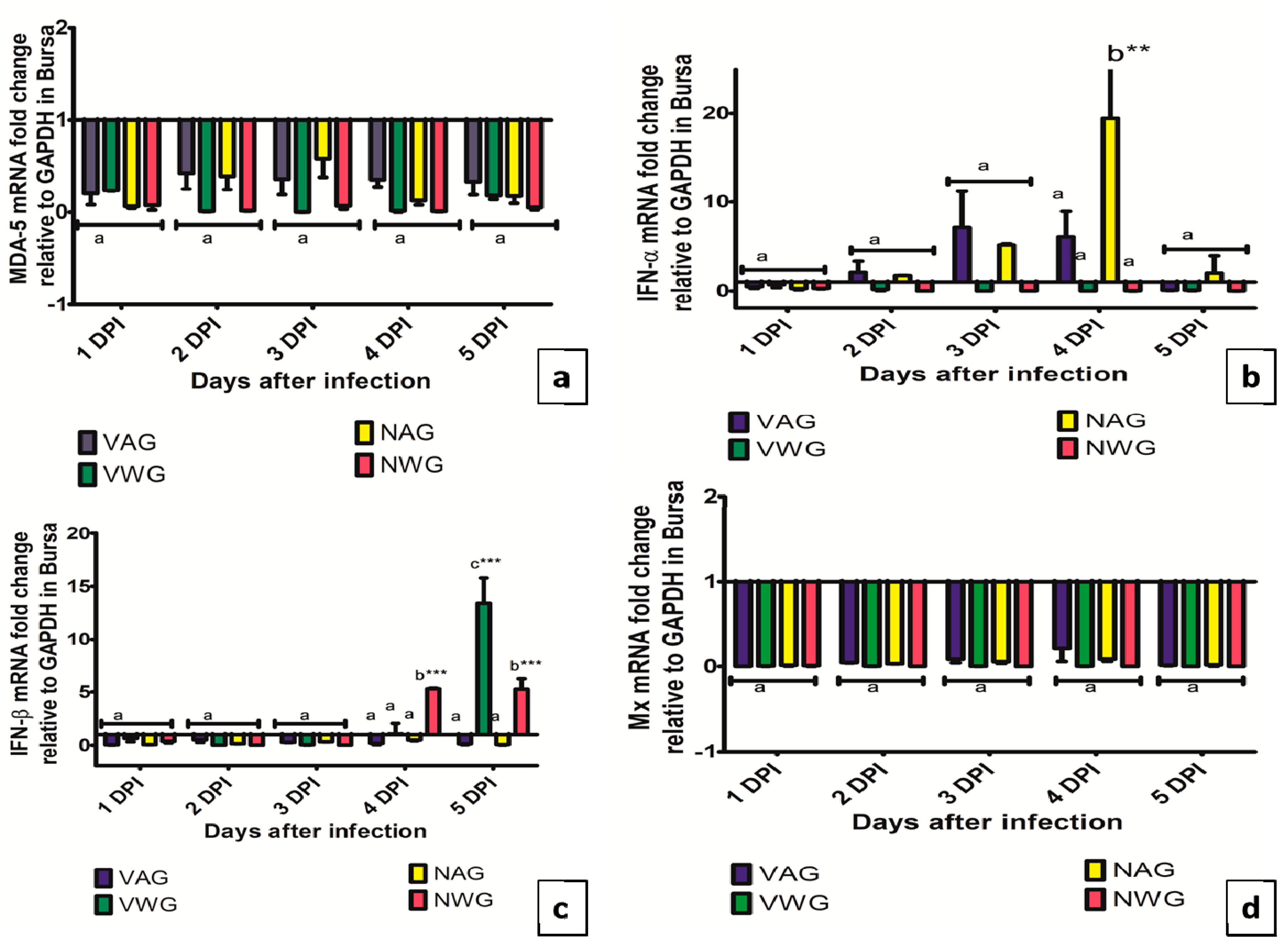
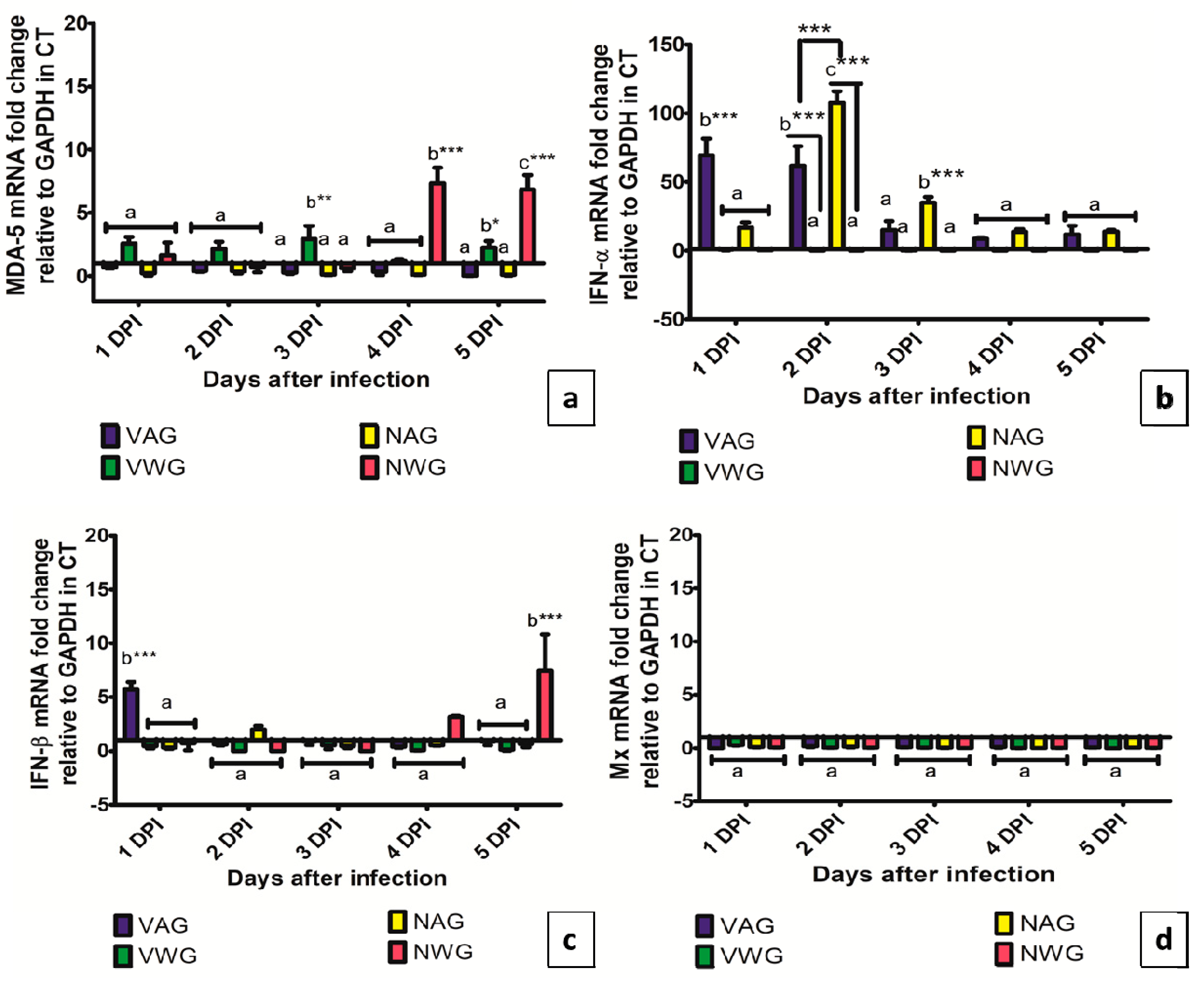
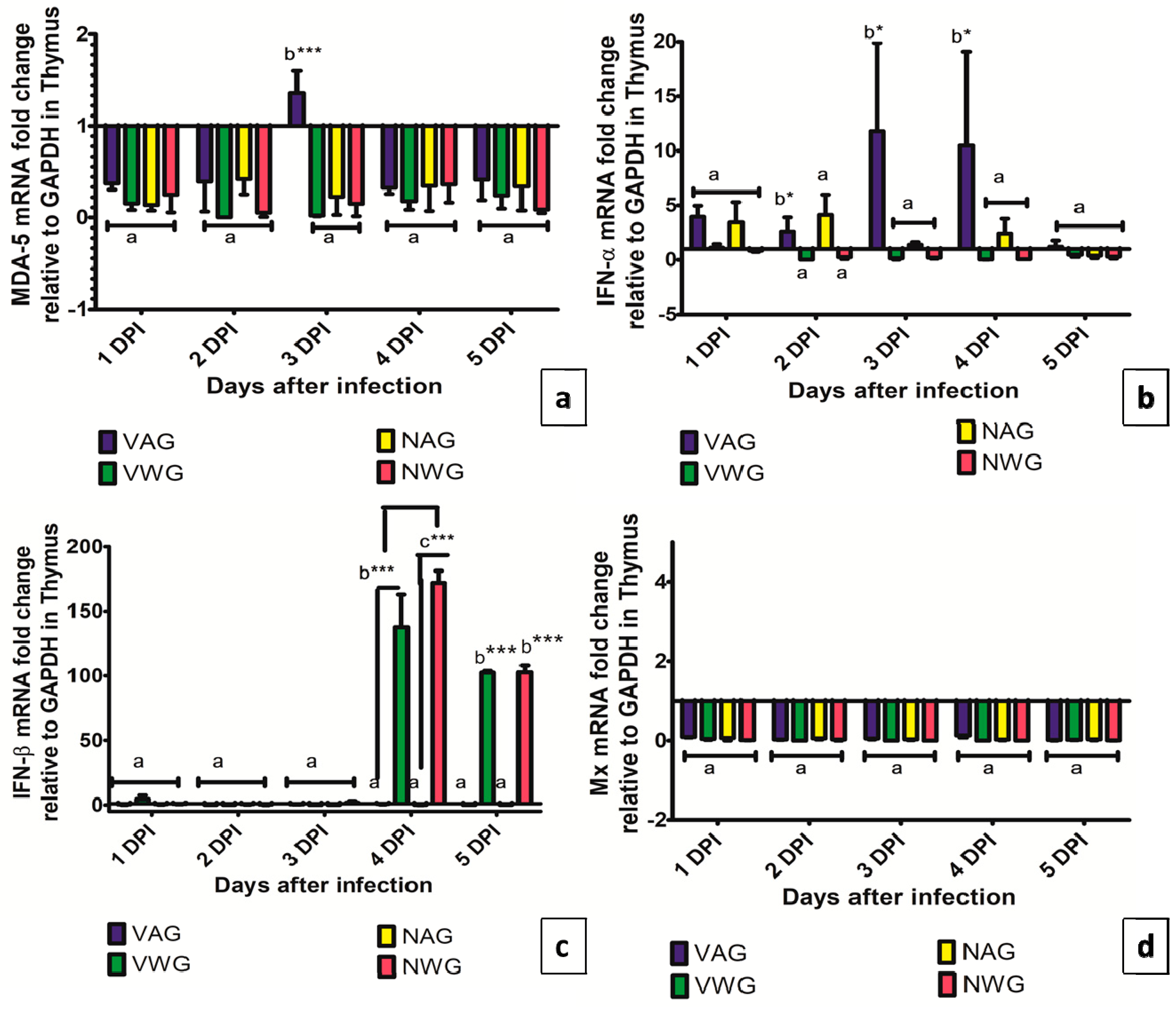
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Kibenge, F.S.B.; Dhillon, A.S.; Russel, R.G. Biochemistry and immunology of Infectious Bursal Disease virus. J. Gen. Virol. 1988, 69, 1757–1775. [Google Scholar] [CrossRef] [PubMed]
- Kegne, T.; Chanie, M. Review on the Incidence and pathology of Infectious Bursal Disease. Br. J. Poult. Sci. 2014, 3, 68–77. [Google Scholar]
- Prandini, F.; Simon, B.; Jung, A.; Poppel, M.; Lemiere, S.; Rautenschlein, S. Comparison of infectious bursal disease live vaccines and a hvt-ibd vector vaccine and their effects on the immune system of commercial layer pullets. Avian Pathol. 2016, 45, 114–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chrysostome, C.A.A.M.; Bell, J.G.; Demey, F.; Verhulst, A. Sero prevalencies to three diseases in village chickens in Benin. Prev. Vet. Med. 1995, 22, 257–261. [Google Scholar] [CrossRef]
- Bumstead, N.; Reece, R.L.; Cook, J.K. Genetic differences in susceptibility of chicken lines to infection with infectious bursal disease virus. Poult. Sci. 1993, 72, 403–410. [Google Scholar] [CrossRef]
- Nielsen, O.L.; Sorensen, P.; Hedemand, J.E.; Laursen, S.B.; Jorgensen, P.H. Inflammatory response of different chicken lines and B haplotypes to infection with infectious bursal disease virus. Avian Pathol. 1998, 27, 181–189. [Google Scholar] [CrossRef]
- Tippenhauer, M.; Heller, D.E.; Weigend, S.; Rautenschlein, S. The host genotype influences infectious bursal disease virus pathogenesis in chickens by modulation of T cells responses and cytokine gene expression. Dev. Comp. Immunol. 2013, 40, 1–10. [Google Scholar] [CrossRef]
- Igwe, A.O. Comparative study of genetic influence on the susceptibility of exotic cockerels, pullets and broilers to infectious bursal disease virus. Niger. Vet. J. 2017, 38, 235–249. [Google Scholar]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Beutler, B.; Eidenschenk, C.; Crozat, K.; Imler, J.L.; Takeuchi, O.; Hoffmann, J.A.; Akira, S. Genetic analysis of resistance to viral infection. Nat. Rev. Immunol. 2007, 7, 753–766. [Google Scholar] [CrossRef]
- Medzhitov, R. Recognition of microorganisms and activation of the immune response. Nature 2007, 449, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Ingrao, F.; Rauw, F.; Lambrecht, B.; van den Berg, T. Infectious Bursal disease: A complex host–pathogen interaction. Dev. Comp. Immunol. 2013, 41, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Bruns, A.M.; Horvath, C.M. Antiviral RNA recognition and assembly by RLR family innate immune sensors. Cytokine Growth Factor Rev. 2014, 25, 507–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoneyama, M.; Onomoto, K.; Jogi, M.; Akaboshi, T.; Fujita, T. Viral RNA detection by RIG-I-like receptors. Curr. Opin. Immunol. 2015, 32, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Meylan, E.; Curran, J.; Hofmann, K.; Moradpour, D.; Binder, M.; Bartenschlager, R.; Tschopp, J. Cardif is an adaptor protein in the RIG-I antiviral pathway and is targeted by hepatitis C virus. Nature 2005, 437, 1167–1172. [Google Scholar] [CrossRef]
- Seth, R.B.; Sun, L.; Ea, C.K.; Chen, Z.J. Identification and characterization of MAVS, a mitochondrial antiviral signaling protein that activates NF-κB and IRF3. Cell 2005, 122, 669–682. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.G.; Wang, Y.Y.; Han, K.J.; Li, L.Y.; Zhai, Z.; Shu, H.B. VISA is an adapter protein required for virus-triggered IFN-β signaling. Mol. Cell 2005, 19, 727–740. [Google Scholar] [CrossRef]
- Stark, G.R.; Kerr, I.M.; Williams, B.R.; Silverman, R.H.; Schreiber, R.D. How cells respond to interferons. Annu. Rev. Biochem. 1998, 67, 227–264. [Google Scholar] [CrossRef] [Green Version]
- Stetson, D.B.; Medzhitov, R. Type I interferons in host defense. Immunity 2006, 25, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Muller, U.; Steinhoff, U.; Reis, L.F.; Hemmi, S.; Pavlovic, J.; Zinkernagel, R.M.; Aguet, M. Functional role of type I and type I1 interferons in antiviral defense. Science 1994, 264, 8–12. [Google Scholar] [CrossRef]
- Horisberger, M.A. Interferons, Mx genes, and resistance to influenza virus. Am. J. Respir. Crit. Care Med. 1995, 152, S67–S71. [Google Scholar] [CrossRef] [PubMed]
- Kochs, G.; Haller, O. GTP-bound human Mx protein interacts with the nucleocapsids of Thogoto virus. J. Biol. Chem. 1999, 274, 4370–4376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randall, R.E.; Goodbourn, S. Interferons and viruses: Interplay between induction, signalling, antiviral responses and virus countermeasures. J. Gen. Virol. 2008, 89, 1–47. [Google Scholar] [CrossRef]
- Xing, Z.; Harper, R.; Anunciacion, J.; Yang, Z.; Gao, W.; Qu, B.; Cardona, C.J. Host immune and apoptotic responses to avian influenza virus H9N2 in human tracheobronchial epithelial cells. Am. J. Respir. Cell Mol. Biol. 2011, 44, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Rue, C.A.; Susta, L.; Cornax, I.; Brown, C.C.; Kapczynski, D.R.; Suarez, D.L.; Afonso, C.L. Virulent Newcastle disease virus elicits a strong innate immune response in chickens. J. Gen. Virol. 2011, 92, 931–939. [Google Scholar] [CrossRef]
- Lee, C.C.; Wu, C.C.; Lin, T.L. Chicken melanoma differentiation-associated gene 5 (MDA5) recognizes infectious bursal disease virus infection and triggers MDA5-related innate immunity. Arch. Virol. 2014, 159, 1671–1686. [Google Scholar] [CrossRef]
- Rodriguez-Chavez, I.R.; Rosenberger, J.K.; Cloud, S.S. Characterization of the antigenic, immunogenic, and pathogenic variation of infectious bursal disease virus due to propagation in different host systems (bursa, embryo, and cell culture). II. Antigenicity at the epitope level. Avian Pathol. 2002, 31, 473–483. [Google Scholar] [CrossRef] [Green Version]
- Villegas, P. Titration of biological suspensions. In A Laboratory Manual for the Isolation and Identification of Avian Pathogens, 4th ed.; Swayne, D.E., Glisson, J.R., Jackwood, M.W., Pearson, J.E., Reed, W.M., Eds.; American Association of Avian Pathogens: Kennett Square, PA, USA, 1998; pp. 248–253. [Google Scholar]
- Tanimura, N.; Tsukamoto, K.; Nakamura, K.; Narita, M.; Maeda, M. Association between pathogenicity of infectious bursal disease virus and viral antigen distribution detected by immunohistochemistry. Avian Dis. 1995, 39, 9–20. [Google Scholar] [CrossRef]
- Williams, A.E.; Davison, T.F. Enhanced immunopathology induced by very virulent infectious bursal disease virus. Avian Pathol. 2005, 34, 4–14. [Google Scholar] [CrossRef]
- Pantin-Jackwood, M.J.; Brown, T.P. Infectious bursal disease virus and proventriculitis in broiler chickens. Avian Dis. 2003, 47, 681–690. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, A.I.; Kulkarni, R.R.; Sharif, S. Synthetic double-stranded RNA oligonucleotides are immunostimulatory for chicken spleen cells. Dev. Comp. Immunol. 2011, 35, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C.; Wu, C.C.; Lin, T.L. Characterization of chicken melanoma differentiation-associated gene 5 (MDA5) from alternative translation initiation. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Brahmakshatriya, V.; Lupiani, B.; Reddy, S.; Okimoto, R.; Li, X.; Chiang, H.; Zhou, H. Associations of chicken Mx1 polymorphism with antiviral responses in avian influenza virus infected embryos and broilers. Poult. Sci. 2012, 91, 3019–3024. [Google Scholar] [CrossRef]
- Li, Y.P.; Handberg, K.J.; Kabell, S.; Kusk, M.; Zhang, M.F.; Jorgensen, P.H. Relative quantification and detection of different types of infectious bursal disease virus in bursa of Fabricius and cloacal swabs using real time RT-PCR SYBR green technology. Res. Vet. Sci. 2007, 82, 126–133. [Google Scholar] [CrossRef]
- Mackay, I. Development and evaluation of a quantitative PCR for the determination of cytomegalovirus load. In Proceedings of the Abstracts of the XIth International Congress of Virology, Sydney, Australia, 9–13 August 1999; VP65 Volume 23, p. 63. [Google Scholar]
- Raj, G.D.; Rajanathan, T.C.; Kumanan, K.; Elankumaran, S. Changes in the cytokine and Toll-Like receptor gene expression following infection of indigenous and commercial chickens with infectious bursal disease virus. Indian J. Virol. 2011, 22, 146–151. [Google Scholar] [CrossRef] [Green Version]
- Ignjatovic, J. Very Virulent Infectious Bursal Disease Virus; Australia and New Zealand Standard Diagnostic Procedures; SCAHLS: Canberra, Australia, 2004; p. 6.
- Haghighi, S.Z.M.; Tavasoly, A.; Marjanmehr, S.H.; Shoshtary, A.; Bahmaninejad, M.A. An experimental study on early pathogenesis of a very virulent isolate of infectious bursal disease virus, employing immunohistochemistry. Iran. J. Vet. Res. 2009, 10, 125–131. [Google Scholar]
- Singh, J.; Banga, H.S.; Brar, R.S.; Singh, N.D.; Sodhi, S.; Leishangthem, G.D. Histopathological and immunohistochemical diagnosis of infectious bursal disease in poultry birds. Vet. World 2015, 8, 1331–1339. [Google Scholar] [CrossRef] [Green Version]
- Prabhu, S.N.; Singh, A.P.; Varghese, B.P.; Kumar, M.A.; Palanivelu, M.; Dhama, K.; Singh, S.D.; Singh, R. Comparative study of very virulent infectious bursal disease virus induced bursal pathology in experimentally infected Indian native Aseel and exotic White Leghorn chicken. Indian J. Vet. Pathol. 2017, 41, 94–101. [Google Scholar] [CrossRef]
- Scanavini Neto, H.; Ito, N.M.K.; Miyaji, C.I.; Lima, E.D.A.; Okabayashi, S.; Corrêa, A.R.A.; Eleutério, G.C.; Zuanaze, M.A. Infectious bursal disease virus: Case report and experimental studies in vaccinated and unvaccinated SPF chickens and commercial broiler chicks. Rev. Bras. Cienc. Avic. 2004, 6, 41–54. [Google Scholar] [CrossRef]
- Survashe, B.D.; Aitken, I.D. Immunocompetent cells in avian paraocular glands. Res. Vet. Sci. 1978, 24, 182–190. [Google Scholar] [CrossRef]
- Eterradossi, N.; Saif, Y.M. Infectious bursal disease. In Diseases of Poultry, 12th ed.; Hofstad, M.S., Barnes, H.J., Calnek, B.W., Reid, W.M., Yoder, H.W., Eds.; Iowa State University Press: Ames, IA, USA, 2008; pp. 194–195. [Google Scholar]
- Hemalatha, S.; Manohar, B.M.; Balachandran, C. Sequential immunohistochemical detection of infectious bursal disease virus antigen in experimental chicken. Indian J. Vet. Pathol. 2009, 33, 82–84. [Google Scholar]
- Zakeri, A.; Kashefi, P. A Study of Apoptosis in Harderian Gland of Infected Chickens by IBDV (Infectious Bursal Disease Virus) with Using EM (Electronic Microscope). Glob. Vet. 2011, 7, 438–442. [Google Scholar]
- Tanimura, N.; Sharma, J.M. Appearance of T cells in the bursa of Fabricius and cecal tonsils during the acute phase of infectious bursal disease virus infection in chickens. Avian Dis. 1997, 41, 638–645. [Google Scholar] [CrossRef]
- Corley, M.M.; Giambrone, J.J.; Dormitorio, T.V. Detection of Infectious Bursal Disease Vaccine Viruses in Lymphoid Tissues after in ovo Vaccination of Specific-Pathogen-Free Embryos. Avian Dis. 2001, 45, 897–905. [Google Scholar] [CrossRef]
- Kim, I.J.; You, S.K.; Kim, H.; Yeh, H.Y.; Sharma, J.M. Characteristics of bursal T lymphocytes induced by infectious bursal disease virus. J. Virol. 2000, 74, 8884–8892. [Google Scholar] [CrossRef] [Green Version]
- Kabell, S.; Handberg, K.J.; Li, Y.; Kusk, M.; Bisgaard, M. Detection of vvIBDV in vaccinated SPF chickens. Acta Vet. Scand. 2005, 46, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Kaufer, I.; Weiss, E.U. Significance of bursa of Fabricius as target organ in infectious bursal disease of chickens. Infect. Immun. 1980, 27, 364–367. [Google Scholar] [CrossRef] [Green Version]
- Mc Ferran, J.B. Infectious Bursal Disease. In Virus Infections of Birds; McFerran, J.B., McNulty, M.S., Eds.; Elsevier Science Publishers B.V.: Amsterdam, The Netherlands, 1993; pp. 213–228. [Google Scholar]
- Tanimura, N.; Sharma, J.M. In-Situ apoptosis in chickens infected with infectious bursal disease virus. J. Comp. Pathol. 1998, 118, 15–27. [Google Scholar] [CrossRef]
- Sick, C.; Schultz, U.; Staeheli, P. A family of genes coding for two serologically distinct chicken interferons. J. Biol. Chem. 1996, 271, 7635–7639. [Google Scholar] [CrossRef] [Green Version]
- Staeheli, P.; Puehler, F.; Schneider, K.; Göbel, T.W.; Kaspers, B. Cytokines of birds: Conserved functions- a largely different look. J. Interferon Cytokine Res. 2001, 21, 993–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.; Sadeyen, J.R.; Butter, C.; Kaiser, P.; Burt, D.W. Analysis of the early immune response to infection by infectious bursal disease virus in chickens differing in their resistance to the disease. J. Virol. 2015, 89, 2469–2482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauf, A. Persistence, Distribution and Immunopathogenesis of Infectious Bursal Disease Virus in Chickens. Ph.D. Thesis, The Ohio State University, Columbus, OH, USA, 2011; p. 75. [Google Scholar]
- Qu, H.; Yang, L.; Meng, S.; Xu, L.; Bi, Y.; Jia, X.; Li, J.; Sun, L.; Liu, W. The differential antiviral activities of chicken interferon α (ChIFN-α) and ChIFN-β are related to distinct interferon-stimulated gene expression. PLoS ONE 2013, 8, 59307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gitlin, L.; Barchet, W.; Gilfillan, S.; Cella, M.; Beutler, B.; Flavell, R.A.; Colonna, M. Essential role of MDA-5 in type I IFN responses to polyriboinosinic: Polyribocytidylic acid and encephalomyocarditis picornavirus. Proc. Natl. Acad. Sci. USA 2006, 103, 8459–8464. [Google Scholar] [CrossRef] [Green Version]
- Kato, H.; Takeuchi, O.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Matsui, K.; Uematsu, S.; Jung, A.; Kawai, T.; Ishii, K.J.; et al. Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature 2006, 441, 101–105. [Google Scholar] [CrossRef]
- Larsen, R.; Røkenes, T.P.; Robertsen, B. Inhibition of infectious pancreatic necrosis virus replication by Atlantic salmon Mx1 protein. J. Virol. 2004, 78, 7938–7944. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.H.; Jin, H.K.; Asano, A.; Takada, A.; Ninomiya, A.; Kida, H.; Hokiyama, H.; Ohara, H.; Tsuzuki, M.; Nishibori, M.; et al. Polymorphism and the differential antiviral activity of the chicken Mx gene. Genome Res. 2002, 12, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Sartika, T.; Sulandari, S.; Zein, M.S.A. Selection of Mx gene genotype as genetic marker for Avian Influenza resistance in Indonesian native chicken. In BMC Proceedings; Biomed Central: London, UK, 2011; Volume 5, p. S37. [Google Scholar]
- Fulton, J.E.; Arango, J.; Ali, R.A.; Bohorquez, E.B.; Lund, A.R.; Ashwell, C.M.; Settar, P.; O’Sullivan, N.P.; Koci, M.D. Genetic variation within the Mx gene of commercially selected chicken lines reveals multiple haplotypes, recombination and a protein under selection pressure. PLoS ONE 2014, 9, 108054. [Google Scholar] [CrossRef] [Green Version]
- Sharma, J.M.; Lee, L.F. Effect of infectious bursal disease on natural killer cell activity and mitogenic response of chicken lymphoid cells: Role of adherent cells in cellular immune suppression. Infect. Immun. 1983, 42, 747–754. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′ → 3′) | Product Size (bp) | Slope * | Amplification Factor | PCR Efficiency |
|---|---|---|---|---|---|
| IFN-α [33] | ATCCTGCTGCTCACGCTCCTTCT GGTGTTGCTGGTGTCCAGGATG | 113 | −3.295 | 2.01 | 101.14 |
| IFN-β [34] | ACCAGGATGCCAACTTCTCTTGGA ATGGCTGCTTGCTTCTTGTCCTTG | 145 | −3.673 | 1.87 | 87.18 |
| MX [35] | GCACACACCCAACTGTCAGCGA CCCATGTCCGAAACTCTCTGCGG | 156 | −3.182 | 2.06 | 106.19 |
| MDA5 [34] | TGCAAGGGAGCTGTTGAGCAGAAT GGCTTTCATGTTGGGTTTCTCGCA | 89 | −3.251 | 2.03 | 103.05 |
| GAPDH [35] | ATCAAGAGGGTAGTGAAGGCTGCT TCAAAGGTGGAGGAATGGCTGTCA | 116 | −3.441 | 1.95 | 95.26 |
| vvIBDVVP2 gene [36] | AGATAACCCAGCCAATCAC CACTCTTTCGTAAGCTACTAGTG | 173 | −3.239 | 2.036 | 103.5 |
| Group | White Leghorn | Aseel | ||||
|---|---|---|---|---|---|---|
| VWG | NWG | CWG | VAG | NAG | CAG | |
| 1 dpi | + | + | - | - | - | - |
| 2 dpi | + | ++ | - | - | ++ | - |
| 3 dpi | - | ++ | - | - | ++ | - |
| 4 dpi | + | +++ | - | - | + | - |
| 5 dpi | + | +++ | - | - | + | - |
| Organ | Groups | Day 1 | Day 2 | Day 3 | Day 4 | Day 5 |
|---|---|---|---|---|---|---|
| Bursa | VWG | 2.00 ± 0.0 c | 2.333 ± 0.33 bc | 1.667 ± 0.33 b | 2.333 ± 0.33 b | 2.333 ± 0.33 b |
| NWG | 2.00 ± 0.57 c | 2.667 ± 0.33 c | 3.00 ± 0.57 c | 3.333 ± 0.33 c | 4.333 ± 0.0 c | |
| CWG | 0.0 ± 0.0 a | 0.00 ± 0.0 a | 0.00 ± 0.0 a | 0.00 ± 0.0 a | 0.00 ± 0.0 a | |
| VAG | 0.00 ± 0.0 a | 0.3333 ± 0.33 a | 0.3333 ± 0.33 a | 0.6667 ± 0.33 a | 0.3333 ± 0.33 a | |
| NAG | 1.00 ± 0.0 b | 1.667 ± 0.33 b | 2.667 ± 0.33 c | 3.333 ± 0.33 c | 2.00 ± 0.33 b | |
| CAG | 0.00 ± 0.0 a | 0.00 ± 0.0 a | 0.00 ± 0.0 a | 0.00 ± 0.0 a | 0.00 ± 0.0 a | |
| Spleen | VWG | 0.3333 ± 0.33 a | 1.667 ± 0.33 b | 1.00 ± 0.57 b | 0.6667 ± 0.33 a | 1.00 ± 0.33 b |
| NWG | 0.3333 ± 0.33 a | 2.667 ± 0.33 c | 2.667 ± 0.33 c | 2.333 ± 0.33 b | 2.00 ± 0.33 c | |
| CWG | 0.00 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | |
| VAG | 0.00 ± 0.0 a | 0.6667 ± 0.33 a | 0.3333 ± 0.33 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | |
| NAG | 0.6667 ± 0.33 a | 0.6667 ± 0.33 a | 1.333 ± 1.33 b | 0.0 ± 0.0 a | 0.0 ± 0.0 a | |
| CAG | 0.00 ± 0.00 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | |
| Thymus | VWG | 0.6667 ± 0.33 a | 0.6667 ± 0.33 a | 1.333 ± 0.33 b | 1.667 ± 0.33 b | 1.00 ± 0.00 b |
| NWG | 0.6667 ± 0.33 a | 1.00 ± 0.00 b | 1.667 ± 0.33 b | 2.333 ± 0.33 b | 2.667 ± 0.33 c | |
| CWG | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | |
| VAG | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | |
| NAG | 0.6667 ± 0.33 a | 0.3333 ± 0.33 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | |
| CAG | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | |
| Caecal tonsils | VWG | 0.6667 ± 0.33 a | 1.333 ± 0.33 b | 1.00 ± 0.00 b | 1.667 ± 0.33 b | 2.00 ± 0.00 b |
| NWG | 1.00 ± 0.57 b | 2.333 ± 0.33 c | 2.333 ± 0.33 c | 1.667 ± 0.33 b | 1.667 ± 0.33 b | |
| CWG | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | |
| VAG | 0.3333 ± 0.33 a | 0.3333 ± 0.33 a | 0.0 ± 0.0 a | 0.3333 ± 0.33 a | 0.0 ± 0.0 a | |
| NAG | 1.667 ± 0.33 b | 1.667 ± 0.33 bc | 1.667 ± 0.33 bc | 0.3333 ± 0.33 a | 0.0 ± 0.0 a | |
| CAG | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a |
| White Leghorn | Aseel | |||||
|---|---|---|---|---|---|---|
| Group | VWG | NWG | CWG | VAG | NAG | CAG |
| 1 dpi | + | ++ | - | - | ++ | - |
| 2 dpi | +++ | +++ | - | + | +++ | - |
| 3 dpi | +++ | +++ | - | + | +++ | - |
| 4 dpi | +++ | +++ | - | ++ | +++ | - |
| 5 dpi | +++ | +++ | - | ++ | +++ | - |
| Organ | DPI | VAG | NAG | VWG | NWG |
|---|---|---|---|---|---|
| Bursa | 1 DPI | 1289.3 ± 12.7 a | 1469.8 ± 57.3 a | 948.9 ± 53.4 a | 778.8 ± 118.1 a |
| 2 DPI | 0.0 ± 0.0 a | 23412.9 ± 368.5 d | 5525.2 ± 154.1 c | 1459.5 ± 111.7 b | |
| 3 DPI | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 8615.1 ± 301.8 c | 1492.5 ± 83.4 b | |
| 4 DPI | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 5282.1 ± 231.1 b | 4999.8 ± 236.7 b | |
| 5 DPI | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 1786.8 ± 75.6 b | 8937.8 ± 427 c | |
| CT | 1 DPI | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 12405.3 ± 664.4 b |
| 2 DPI | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 625.7 ± 24.2 a | 1116.0 ± 118.3 a | |
| 3 DPI | 0.0 ± 0.0 a | 868.4 ± 41.5 a | 1114.7 ± 93.9 a | 1170.3 ± 213.9 a | |
| 4 DPI | 0.0 ± 0.0 a | 1030.3 ± 64.1 a | 1064.0 ± 62.5 a | 5092.7 ± 1280.5 b | |
| 5 DPI | 0.0 ± 0.0 a | 1463.9 ± 58.1 a | 1168.7 ± 52.4 a | 4665.0 ± 1062 b | |
| Spleen | 1 DPI | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a |
| 2 DPI | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 5000.7 ± 265 b | 4500.7 ± 331.7 b | |
| 3 DPI | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 1065.3 ± 133.5 b | 7836.0 ± 318.9 c | |
| 4 DPI | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 1282.7 ± 97.1 b | 7984.0 ± 544.7 c | |
| 5 DPI | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 1834.3 ± 482.9 b | 7633.0 ± 380 c | |
| Thymus | 1 DPI | 1428.6 ± 65.7 a | 1459.1 ± 46.2 a | 0.0 ± 0.0 b | 1406.3 ± 85.8 a |
| 2 DPI | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 28369.0 ± 863.8 c | 1301.7 ± 179.9 b | |
| 3 DPI | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 2477.3 ± 207.3 b | 1573.7 ± 65.8 b | |
| 4 DPI | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 1160.3 ± 60.9 b | 1684.7 ± 235.3 b | |
| 5 DPI | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 6070.7 ± 237.7 b | 14470.0 ± 559.5 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prabhu, S.N.; Singh, A.P.; Varghese, B.P.; Dhama, K.; Singh, S.D.; Singh, R. A Comparative Study of Pathology and Host Immune Response Induced by Very Virulent Infectious Bursal Disease Virus in Experimentally Infected Chickens of Aseel and White Leghorn Breeds. Vaccines 2020, 8, 627. https://doi.org/10.3390/vaccines8040627
Prabhu SN, Singh AP, Varghese BP, Dhama K, Singh SD, Singh R. A Comparative Study of Pathology and Host Immune Response Induced by Very Virulent Infectious Bursal Disease Virus in Experimentally Infected Chickens of Aseel and White Leghorn Breeds. Vaccines. 2020; 8(4):627. https://doi.org/10.3390/vaccines8040627
Chicago/Turabian StylePrabhu, Shyama N., Ajay Pratap Singh, Berin P. Varghese, Kuldeep Dhama, Shambhu Dayal Singh, and Rajendra Singh. 2020. "A Comparative Study of Pathology and Host Immune Response Induced by Very Virulent Infectious Bursal Disease Virus in Experimentally Infected Chickens of Aseel and White Leghorn Breeds" Vaccines 8, no. 4: 627. https://doi.org/10.3390/vaccines8040627