Development of Luciferase Immunoprecipitation Systems (LIPS) Assay to Detect IgG Antibodies against Human Respiratory Syncytial Virus G-Glycoprotein

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Antibodies and Sera
2.3. Generation of Renilla Luciferase-Tagged RSV-A2 and RSV-B1 G/mG Protein Constructs
2.4. Renilla Luciferase (Ruc) RSV-GA and RSV-GB Expression
2.5. LIPS-G Assay to Detect RSV-G Specific IgG Antibodies
2.6. Production and Purification of Recombinant RSV Proteins in E. Coli for Competition Assays and Immunizations
2.7. Competition Assay Using Recombinant RSV-GA or RSV-GB Protein
2.8. Determination of Limit of Detection
2.9. Statistical Analysis
3. Results
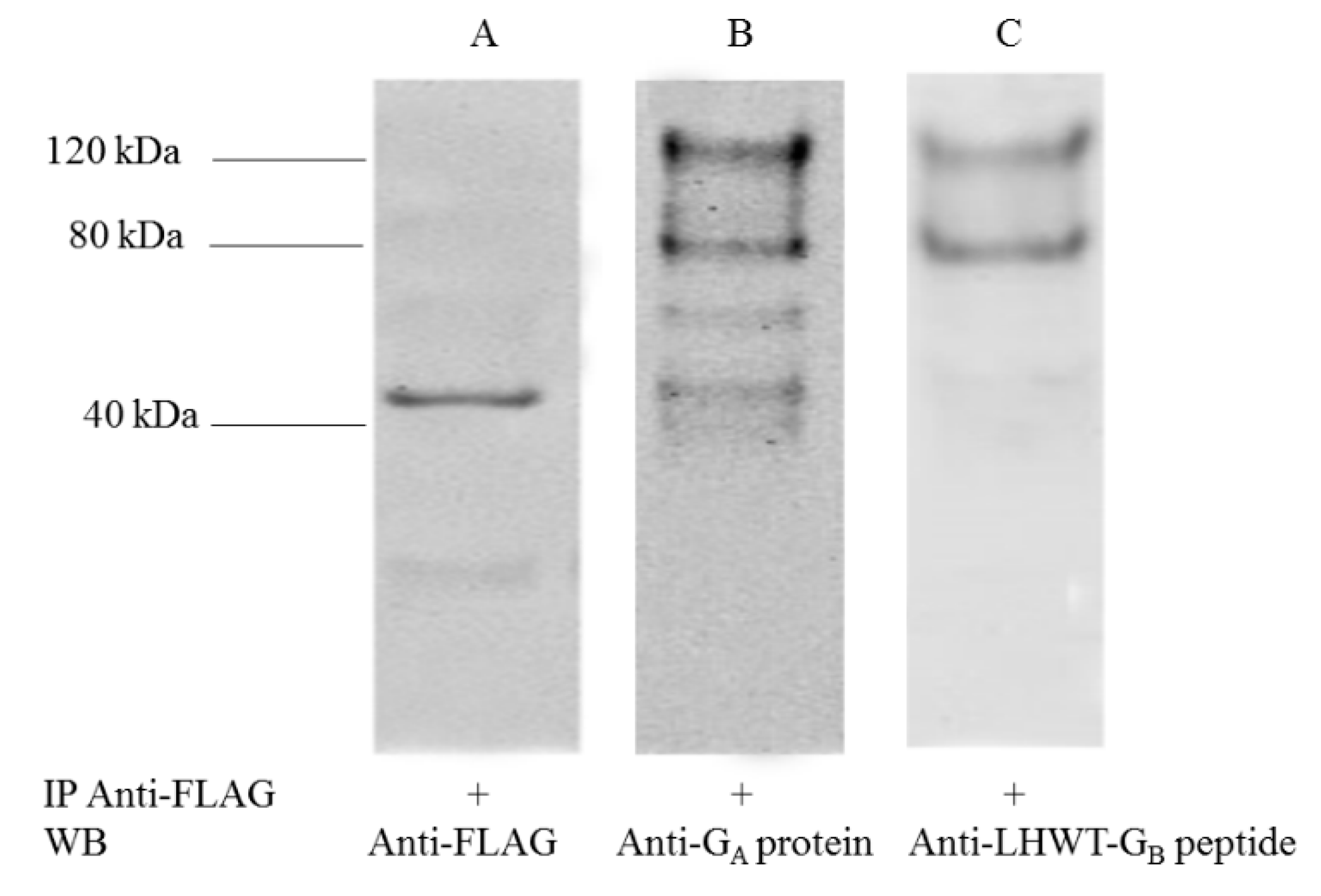
3.1. Expression of Ruc-GA and Ruc-GB Protein in COS-1 Lysates
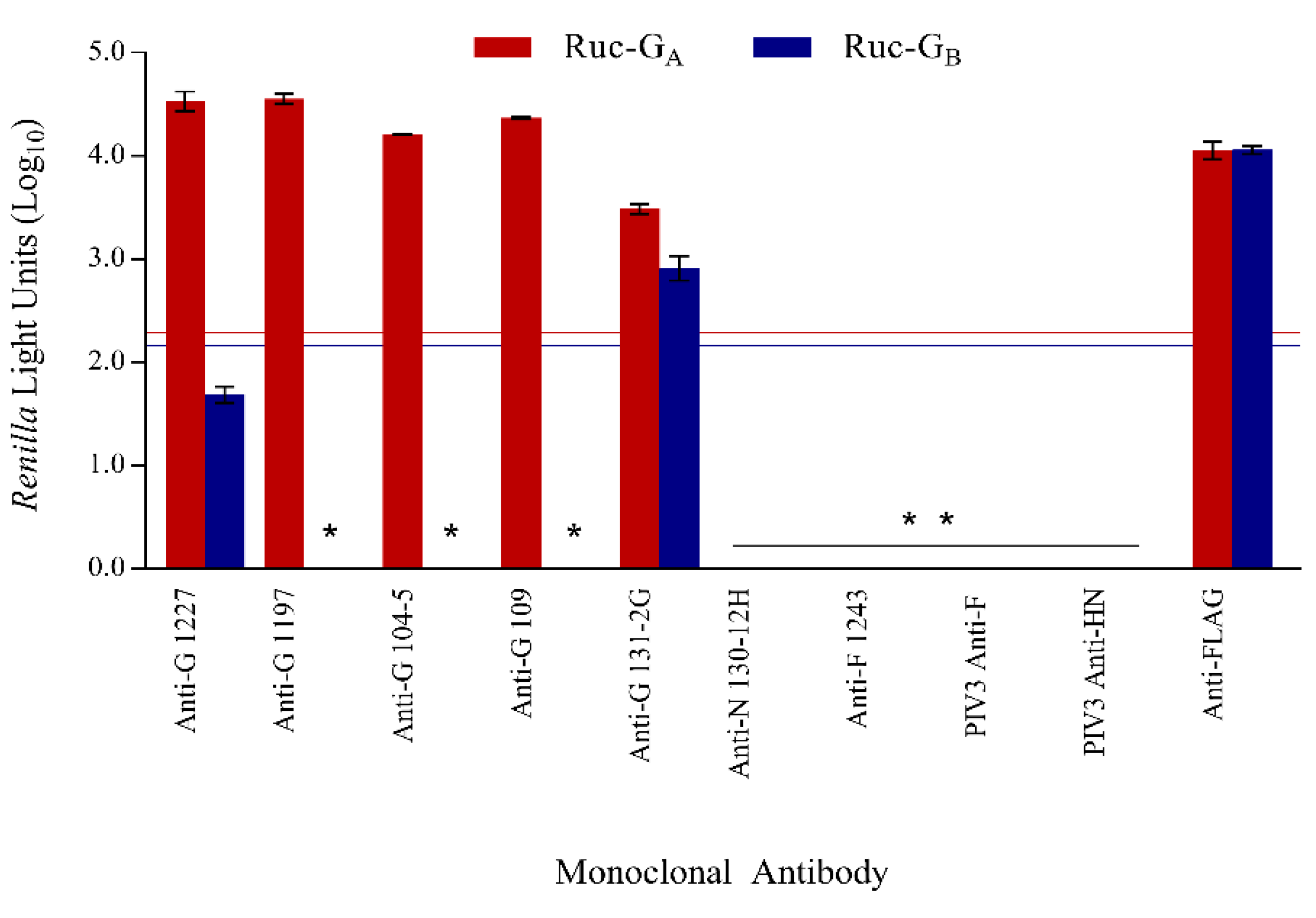
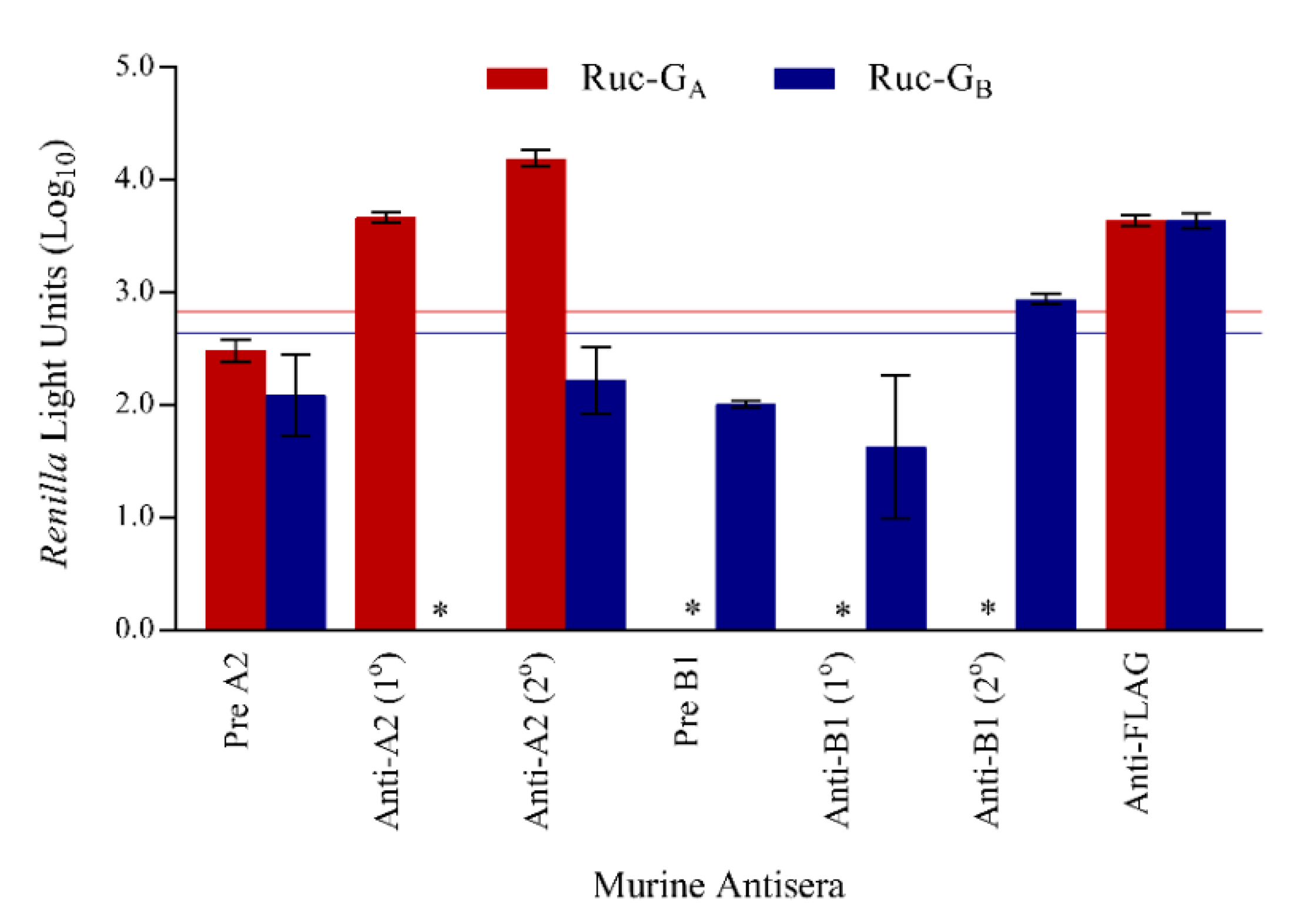
3.2. Assay Specificity
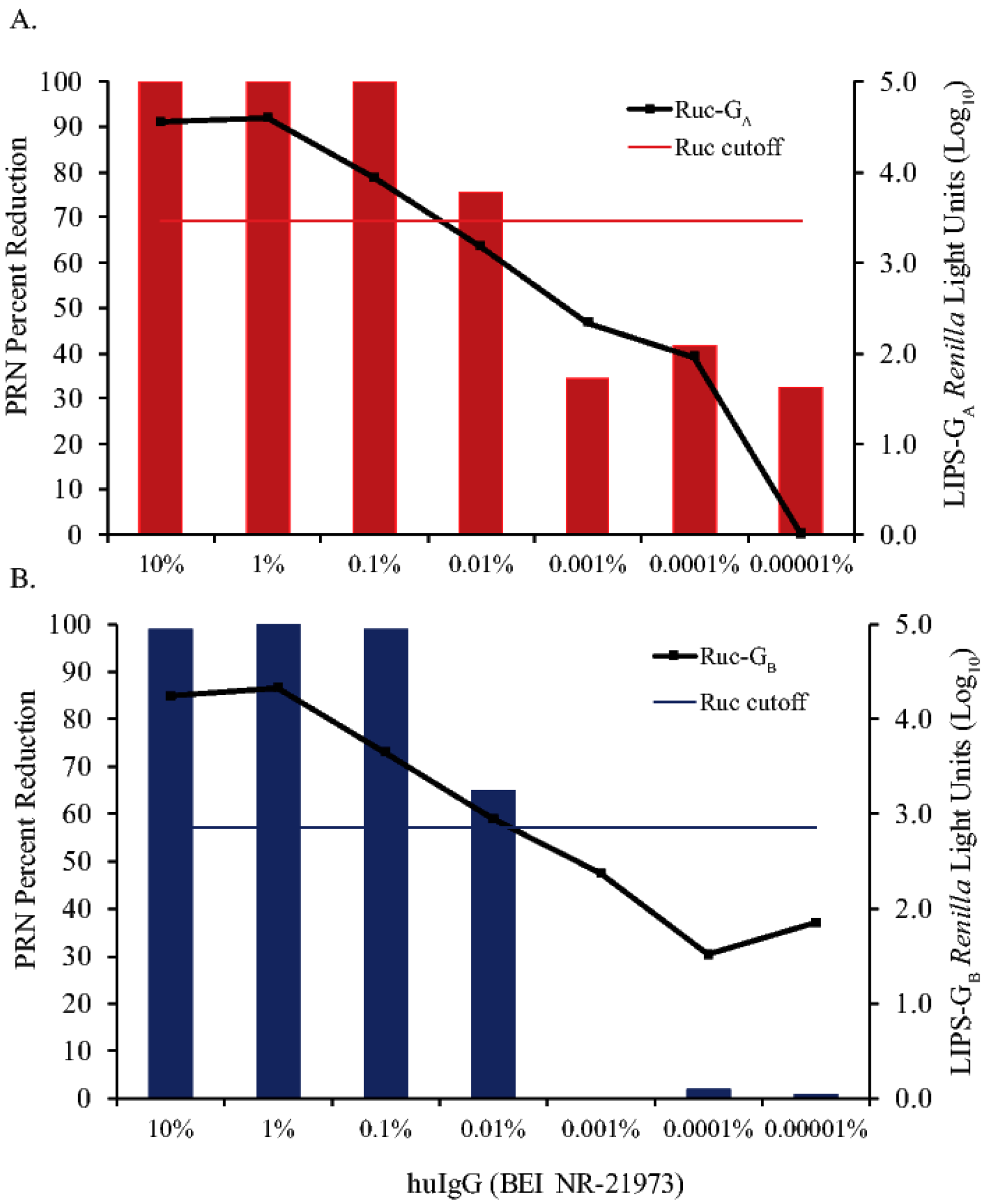
3.3. Sensitivity of LIPS-G Assays in Detecting Anti-RSV-G IgG Antibodies Compared to PRN
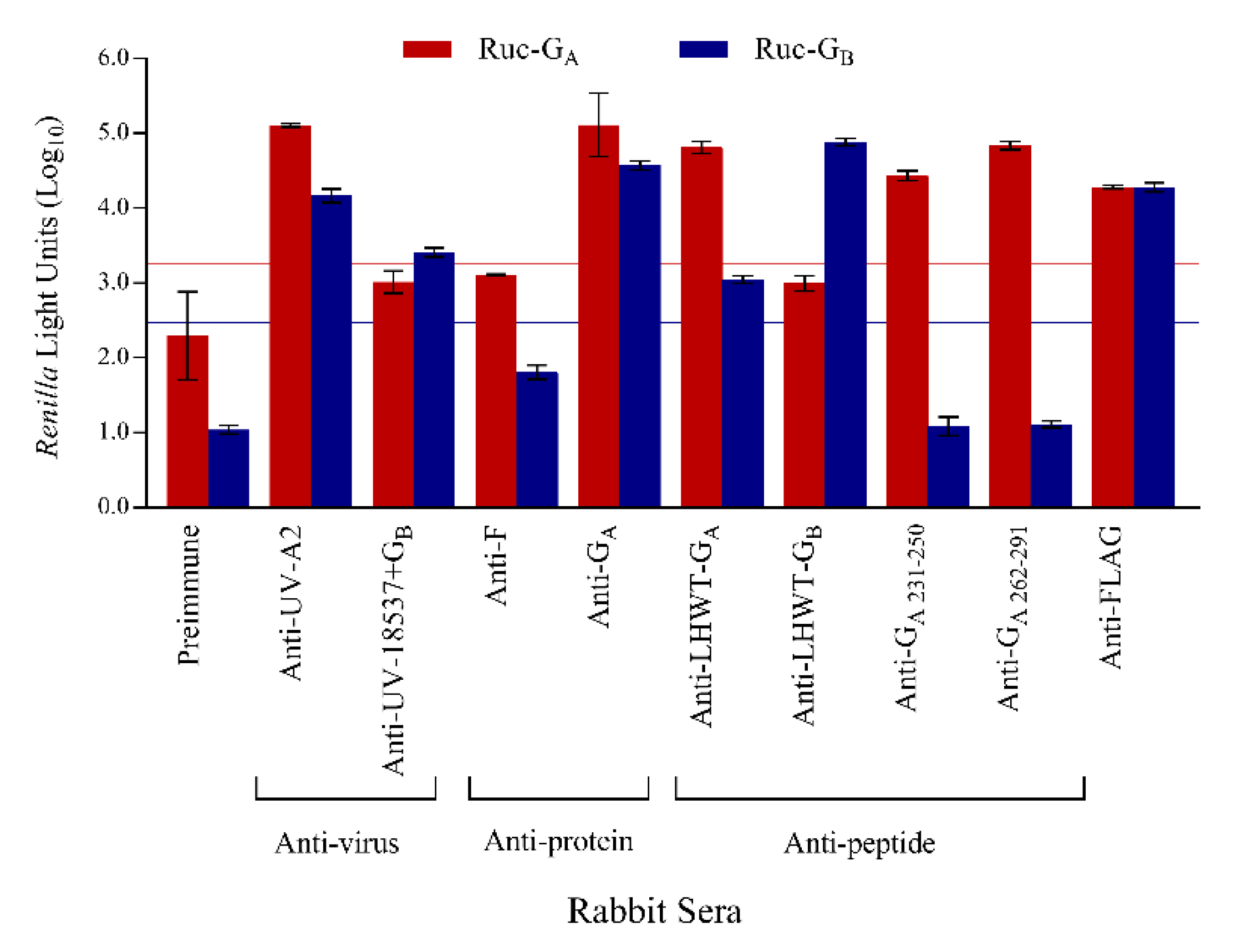
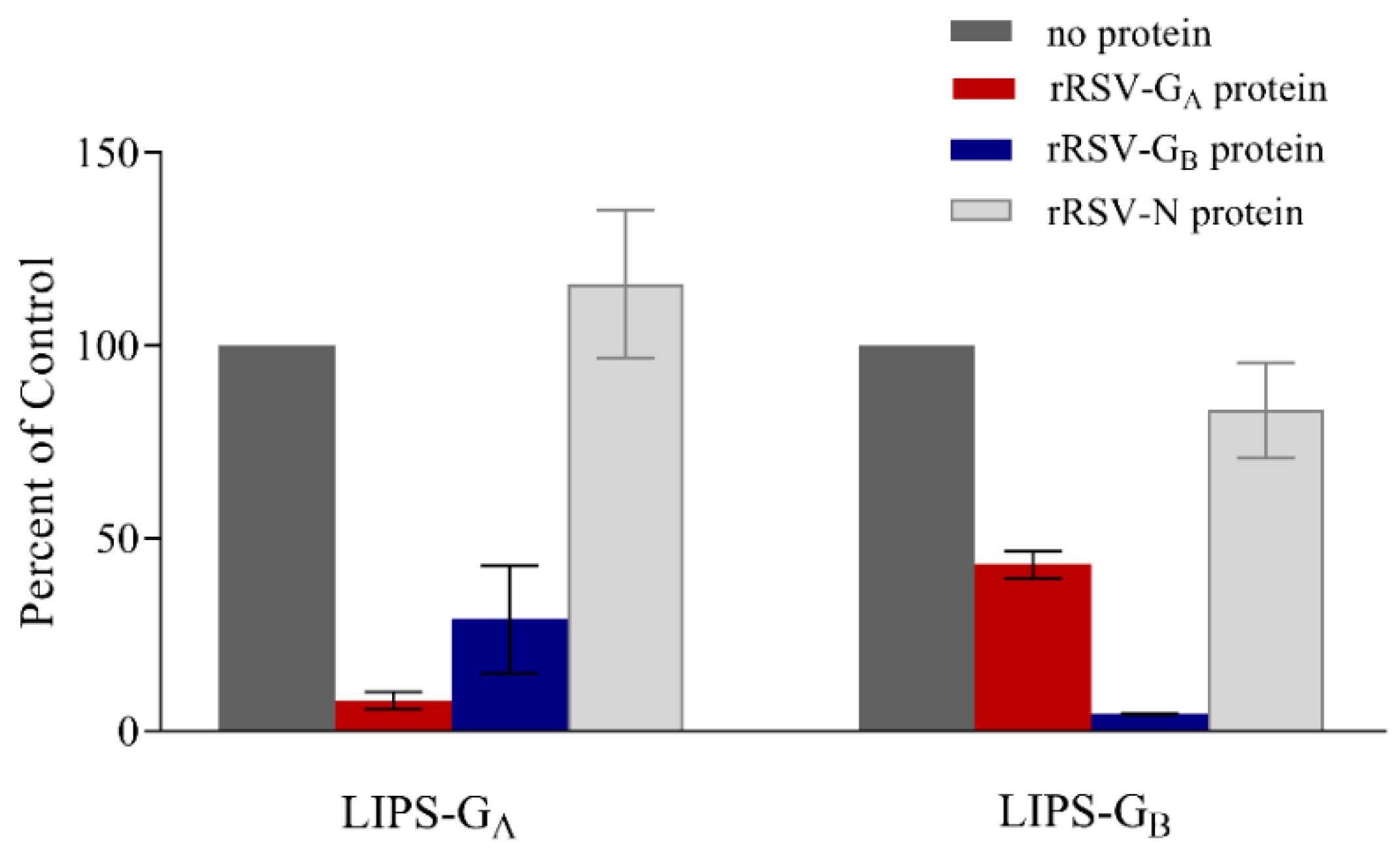
3.4. Confirmation of Specificity
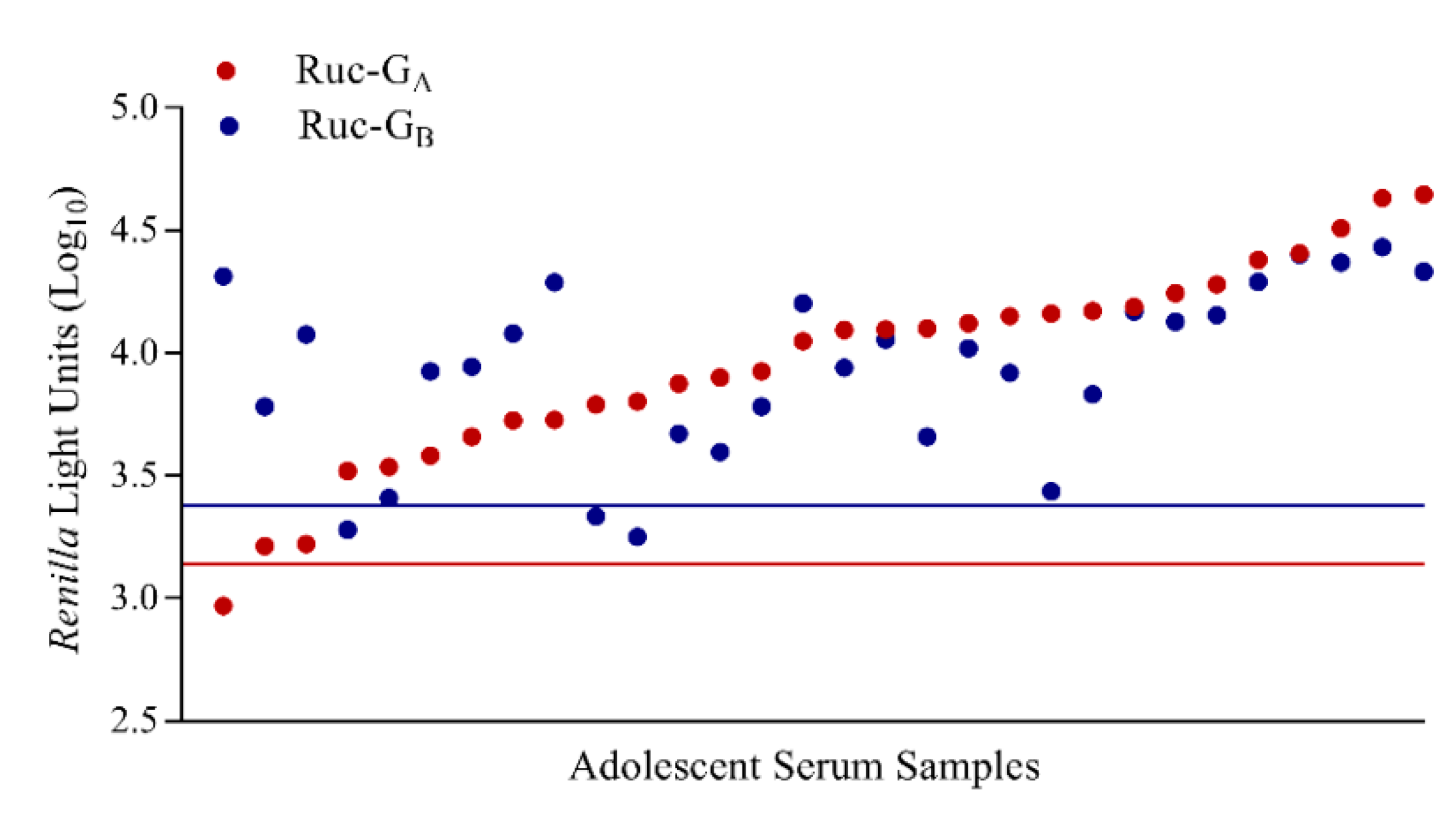
3.5. LIPS-G Assays Detect Antibodies Elicited by Natural Infection
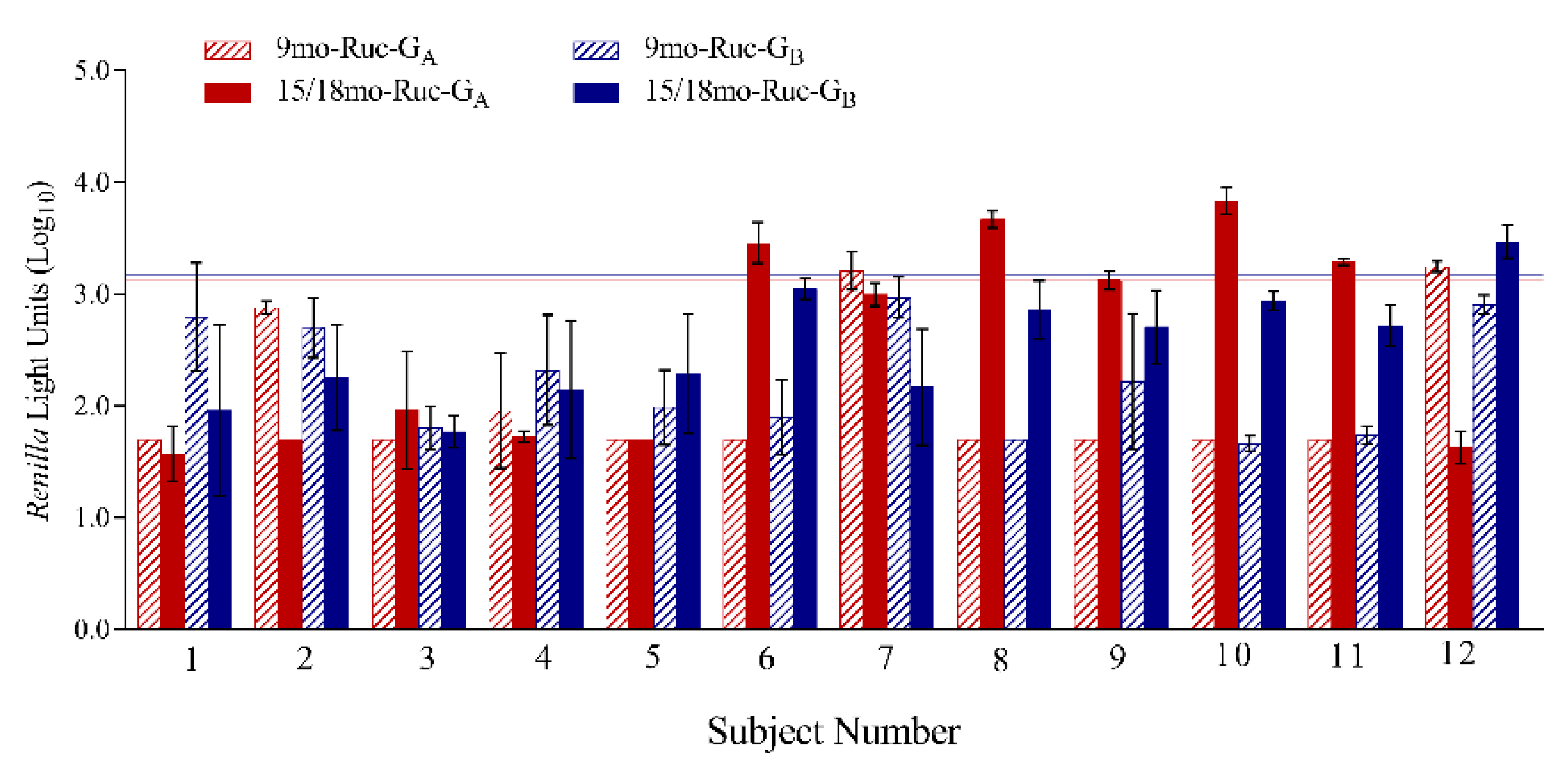
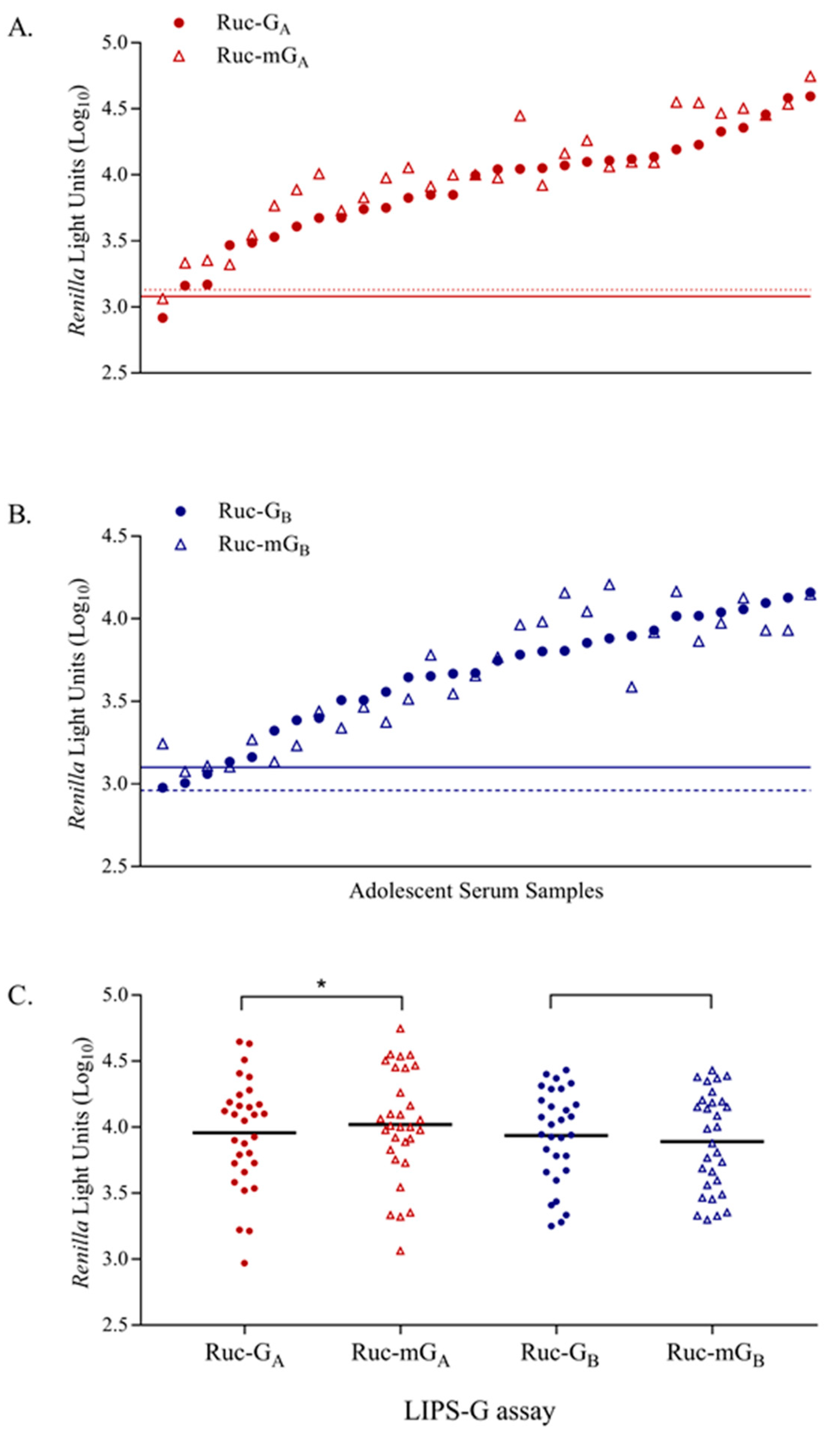
3.6. LIPS-G Assays Detect Anti-RSV-G IgG Specific Responses in Paired Sera from Young Children
3.7. Reactivity of anti-RSV IgG Antibodies with Mutated Ruc-RSV-mG Antigens
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yamin, D.; Jones, F.; de Vincenzo, J.; Gertler, S.; Kobiler, O.; Townsend, J.P.; Galvani, A.P. Vaccination strategies against respiratory syncytial virus. In Proceedings of the National Academy of Sciences, Washington, DC, USA, 27 December 2016; Volume 113, pp. 13239–13244. [Google Scholar]
- Falsey, A.R.; Hennessey, P.A.; Formica, M.; Cox, C.; Walsh, E.E. Respiratory syncytial virus infections in elderly and high-risk adults. N. Engl. J. Med. 2005, 352, 1749–1759. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, S.; Auais, A.; Piedimonte, G. Palivizumab in the prophylaxis of respiratory syncytial virus infection. Expert Rev. Antiinfect. Ther. 2005, 3, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.R.; Graham, B.S.; Prince, G.A.; Walsh, E.E.; Chanock, R.M.; Karzon, D.T.; Wright, P.F. Serum and nasal-wash immunoglobulin G and A antibody response of infants and children to respiratory syncytial virus F and G glycoproteins following primary infection. J. Clin. Microbiol. 1986, 23, 1009–1014. [Google Scholar] [PubMed]
- Ngwuta, J.O.; Chen, M.; Modjarrad, K.; Joyce, M.G.; Kanekiyo, M.; Kumar, A.; Yassine, H.M.; Moin, S.M.; Killikelly, A.M.; Chuang, G.Y.; et al. Prefusion F-specific antibodies determine the magnitude of RSV neutralizing activity in human sera. Sci. Transl. Med. 2015, 7, 309ra162. [Google Scholar] [CrossRef] [PubMed]
- Capella, C.; Chaiwatpongsakorn, S.; Gorrell, E.; Risch, Z.A.; Ye, F.; Mertz, S.E.; Johnson, S.M.; Moore-Clingenpeel, M.; Ramilo, O.; Mejias, A.; et al. Prefusion F, postfusion F, G antibodies, and disease severity in infants and young children with acute respiratory syncytial virus infection. J. Infect. Dis. 2017, 216, 1398–1406. [Google Scholar] [CrossRef] [PubMed]
- Tripp, R.A.; Power, U.F.; Openshaw, P.J.M.; Kauvar, L.M. Respiratory syncytial virus: Targeting the G protein provides a new approach for an old problem. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.M.; McNally, B.A.; Ioannidis, I.; Flano, E.; Teng, M.N.; Oomens, A.G.; Walsh, E.E.; Peeples, M.E. Respiratory syncytial virus uses CX3CR1 as a receptor on primary human airway epithelial cultures. PLoS Pathog. 2015, 11, e1005318. [Google Scholar] [CrossRef] [PubMed]
- Chirkova, T.; Lin, S.; Oomens, A.G.; Gaston, K.A.; Boyoglu-Barnum, S.; Meng, J.; Stobart, C.C.; Cotton, C.U.; Hartert, T.V.; Moore, M.L.; et al. CX3CR1 is an important surface molecule for respiratory syncytial virus infection in human airway epithelial cells. J. Gen. Virol. 2015, 96, 2543–2556. [Google Scholar] [CrossRef] [Green Version]
- Haynes, L.M.; Caidi, H.; Radu, G.U.; Miao, C.; Harcourt, J.L.; Tripp, R.A.; Anderson, L.J. Therapeutic monoclonal antibody treatment targeting respiratory syncytial virus (RSV) G protein mediates viral clearance and reduces the pathogenesis of RSV infection in BALB/c mice. J. Infect. Dis. 2009, 200, 439–447. [Google Scholar] [CrossRef]
- Boyoglu-Barnum, S.; Todd, S.O.; Chirkova, T.; Barnum, T.R.; Gaston, K.A.; Haynes, L.M.; Tripp, R.A.; Moore, M.L.; Anderson, L.J. An anti-G protein monoclonal antibody treats RSV disease more effectively than an anti-F monoclonal antibody in BALB/c mice. Virology 2015, 483, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Collarini, E.J.; Lee, F.E.; Foord, O.; Park, M.; Sperinde, G.; Wu, H.; Harriman, W.D.; Carroll, S.F.; Ellsworth, S.L.; Anderson, L.J.; et al. Potent high-affinity antibodies for treatment and prophylaxis of respiratory syncytial virus derived from B cells of infected patients. J. Immunol. 2009, 183, 6338–6345. [Google Scholar] [CrossRef] [PubMed]
- Cane, P.A.; Thomas, H.M.; Simpson, A.F.; Evans, J.E.; Hart, C.A.; Pringle, C.R. Analysis of the human serological immune response to a variable region of the attachment (G) protein of respiratory syncytial virus during primary infection. J. Med. Virol. 1996, 48, 253–261. [Google Scholar] [CrossRef]
- Cane, P.A. Analysis of linear epitopes recognised by the primary human antibody response to a variable region of the attachment (G) protein of respiratory syncytial virus. J. Med. Virol. 1997, 51, 297–304. [Google Scholar] [CrossRef]
- Lee, J.; Klenow, L.; Coyle, E.M.; Golding, H.; Khurana, S. Protective antigenic sites in respiratory syncytial virus G attachment protein outside the central conserved and cysteine noose domains. PLoS Pathog. 2018, 14, e1007262. [Google Scholar] [CrossRef] [PubMed]
- PATH. RSV Vaccine and mAb Snapshot. Available online: https://www.path.org/resources/rsv-vaccine-and-mab-snapshot/ (accessed on 10 January 2019).
- Zhang, Y.; Sakthivel, S.K.; Bramley, A.; Jain, S.; Haynes, A.; Chappell, J.D.; Hymas, W.; Lenny, N.; Patel, A.; Qi, C.; et al. Serology enhances molecular diagnosis of respiratory virus infections other than influenza in children and adults hospitalized with community-acquired pneumonia. J. Clin. Microbiol. 2017, 55, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Maifeld, S.V.; Ro, B.; Mok, H.; Chu, M.; Yu, L.; Yamagata, R.; Leonardson, T.; Chio, V.; Parhy, B.; Park, S.; et al. Development of electrochemiluminescent serology assays to measure the humoral response to antigens of respiratory syncytial virus. PLoS ONE 2016, 11, e0153019. [Google Scholar] [CrossRef] [PubMed]
- Muelenaer, P.M.; Henderson, F.W.; Hemming, V.G.; Walsh, E.E.; Anderson, L.J.; Prince, G.A.; Murphy, B.R. Group-specific serum antibody responses in children with primary and recurrent respiratory syncytial virus infections. J. Infect. Dis. 1991, 164, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Beeler, J.A.; van Wyke Coelingh, K. Neutralization epitopes of the F glycoprotein of respiratory syncytial virus: Effect of mutation upon fusion function. J. Virol. 1989, 63, 2941–2950. [Google Scholar] [PubMed]
- Burbelo, P.D.; Goldman, R.; Mattson, T.L. A simplified immunoprecipitation method for quantitatively measuring antibody responses in clinical sera samples by using mammalian-produced renilla luciferase-antigen fusion proteins. BMC Biotechnol. 2005, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Varada, J.C.; Teferedegne, B.; Crim, R.L.; Mdluli, T.; Audet, S.; Peden, K.; Beeler, J.; Murata, H. A neutralization assay for respiratory syncytial virus using a quantitative PCR-based endpoint assessment. Virol. J. 2013, 10, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crim, R.L.; Audet, S.A.; Feldman, S.A.; Mostowski, H.S.; Beeler, J.A. Identification of linear heparin-binding peptides derived from human respiratory syncytial virus fusion glycoprotein that inhibit infectivity. J. Virol. 2007, 81, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Mbiguino, A.; Menezes, J. Purification of human respiratory syncytial virus: Superiority of sucrose gradient over percoll, renografin and metrizamide gradients. J. Virol. Methods 1991, 31, 161–170. [Google Scholar] [CrossRef]
- Kumari, S.; Crim, R.L.; Kulkarni, A.; Audet, S.A.; Mdluli, T.; Murata, H.; Beeler, J.A. Development of a luciferase immunoprecipitation system assay to detect IgG antibodies against human respiratory syncytial virus nucleoprotein. Clin. Vaccine Immunol. 2014, 21, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Choi, Y.; Haynes, L.M.; Harcourt, J.L.; Anderson, L.J.; Jones, L.P.; Tripp, R.A. Vaccination to induce antibodies blocking the CX3C-CX3CR1 interaction of respiratory syncytial virus G protein reduces pulmonary inflammation and virus replication in mice. J. Virol. 2010, 84, 1148–1157. [Google Scholar] [CrossRef] [PubMed]
- Bukreyev, A.; Yang, L.; Fricke, J.; Cheng, L.; Ward, J.M.; Murphy, B.R.; Collins, P.L. The secreted form of respiratory syncytial virus G glycoprotein helps the virus evade antibody-mediated restriction of replication by acting as an antigen decoy and through effects on Fc receptor-bearing leukocytes. J. Virol. 2008, 82, 12191–12204. [Google Scholar] [CrossRef] [PubMed]
- Burbelo, P.D.; Ching, K.H.; Klimavicz, C.M.; Iadarola, M.J. Antibody profiling by luciferase immunoprecipitation systems (LIPS). J. Vis. Exp. 2009, 32. [Google Scholar] [CrossRef]
- Reichelt, P.; Schwarz, C.; Donzeau, M. Single step protocol to purify recombinant proteins with low endotoxin contents. Protein Expr. Purif. 2006, 46, 483–488. [Google Scholar] [CrossRef]
- Cohen, B.J.; Audet, S.; Andrews, N.; Beeler, J.; WHO Working Group on Measles Plaque Reduction Neutralization Test. Plaque reduction neutralization test for measles antibodies: Description of a standardised laboratory method for use in immunogenicity studies of aerosol vaccination. Vaccine 2007, 26, 59–66. [Google Scholar] [CrossRef]
- Boyoglu-Barnum, S.; Todd, S.O.; Meng, J.; Barnum, T.R.; Chirkova, T.; Haynes, L.M.; Jadhao, S.J.; Tripp, R.A.; Oomens, A.G.; Moore, M.L.; et al. Mutating the CX3C motif in the G Protein should make a live respiratory syncytial virus vaccine safer and more effective. J. Virol. 2017, 91, JVI-02059. [Google Scholar] [CrossRef]
- Jones, H.G.; Ritschel, T.; Pascual, G.; Brakenhoff, J.P.J.; Keogh, E.; Furmanova-Hollenstein, P.; Lanckacker, E.; Wadia, J.S.; Gilman, M.S.A.; Williamson, R.A.; et al. Structural basis for recognition of the central conserved region of RSV G by neutralizing human antibodies. PLoS Pathog. 2018, 14, e1006935. [Google Scholar] [CrossRef]
- Fedechkin, S.O.; George, N.L.; Wolff, J.T.; Kauvar, L.M.; DuBois, R.M. Structures of respiratory syncytial virus G antigen bound to broadly neutralizing antibodies. Sci. Immunol. 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Varga, S.M.; Wissinger, E.L.; Braciale, T.J. The attachment (G) glycoprotein of respiratory syncytial virus contains a single immunodominant epitope that elicits both Th1 and Th2 CD4 + T cell responses. J. Immunol. 2000, 165, 6487–6495. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence |
|---|---|
| RSV-A2-G Forward | AAGGAATTCAACATGTCCAAAAACAAGGACCAACGCACC |
| RSV-A2-G Reverse | GGGCTCGAGTTAAAGTAACTACTGGCGTGGTGT |
| RSV-B1-G Forward | AAGGAATTCACCATGTCCAAACACAAGAATCAACGCACT |
| RSV-B1-G Reverse | GGGCTCGAGGAATAACTAAGCATGTGATTGGGT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crim, R.L.; Kumari, S.; Jayanti, P.; Audet, S.; Kulkarni, A.; Beeler, J. Development of Luciferase Immunoprecipitation Systems (LIPS) Assay to Detect IgG Antibodies against Human Respiratory Syncytial Virus G-Glycoprotein. Vaccines 2019, 7, 16. https://doi.org/10.3390/vaccines7010016
Crim RL, Kumari S, Jayanti P, Audet S, Kulkarni A, Beeler J. Development of Luciferase Immunoprecipitation Systems (LIPS) Assay to Detect IgG Antibodies against Human Respiratory Syncytial Virus G-Glycoprotein. Vaccines. 2019; 7(1):16. https://doi.org/10.3390/vaccines7010016
Chicago/Turabian StyleCrim, Roberta Lynne, Sangeeta Kumari, Priyanka Jayanti, Susette Audet, Ashwin Kulkarni, and Judy Beeler. 2019. "Development of Luciferase Immunoprecipitation Systems (LIPS) Assay to Detect IgG Antibodies against Human Respiratory Syncytial Virus G-Glycoprotein" Vaccines 7, no. 1: 16. https://doi.org/10.3390/vaccines7010016