
Vaccine Responses in Patients with Liver Cirrhosis: From the Immune System to the Gut Microbiota
 , , and
, , and
Abstract
:1. Introduction
2. Vaccination Efficacy in Patients with Cirrhosis
2.1. Recommendation for Vaccination in Cirrhotic Patients
2.2. Vaccine Response in Liver Transplant Recipients
3. The Immune System: An Overview of the Response to Vaccination
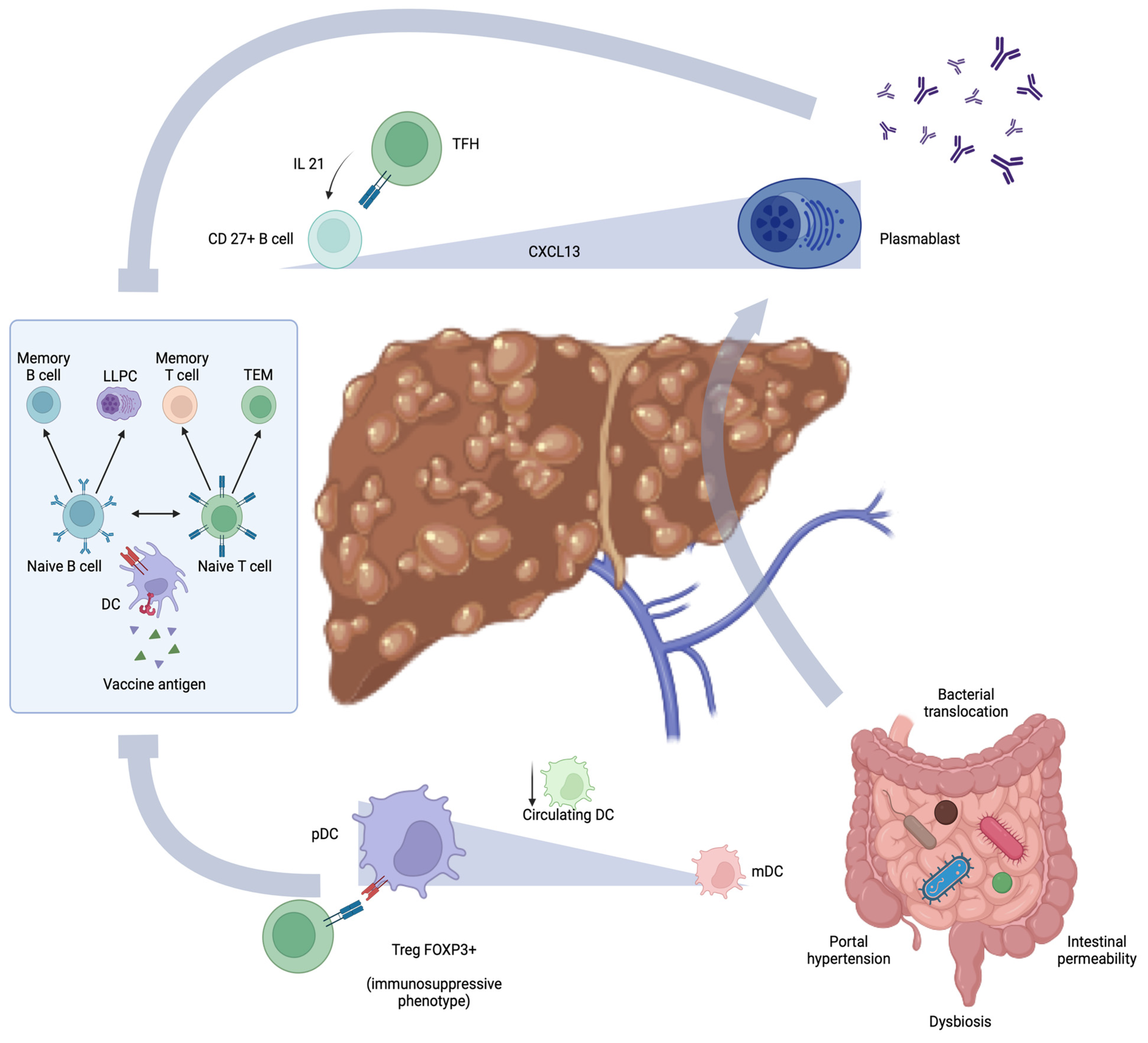
4. Cirrhosis Immune Dysfunction and Defective Immunization
5. Vaccination Response and Gut-Liver Axis in Cirrhosis
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef]
- Wiesen, E.; Diorditsa, S.; Li, X. Progress towards hepatitis B prevention through vaccination in the Western Pacific, 1990–2014. Vaccine 2016, 34, 2855–2862. [Google Scholar] [CrossRef]
- Watson, O.J.; Barnsley, G.; Toor, J.; Hogan, A.B.; Winskill, P.; Ghani, A.C. Global impact of the first year of COVID-19 vaccination: A mathematical modelling study. Lancet Infect. Dis. 2022, 22, 1293–1302. [Google Scholar] [CrossRef]
- Nielsen, B.U.; Drabe, C.H.; Barnkob, M.B.; Johansen, I.S.; Hansen, A.K.K.; Nilsson, A.C.; Rasmussen, L.D. Antibody response following the third and fourth SARS-CoV-2 vaccine dose in individuals with common variable immunodeficiency. Front. Immunol. 2022, 13, 934476. [Google Scholar] [CrossRef]
- Crum-Cianflone, N.F.; Wallace, M.R. Vaccination in HIV-infected adults. AIDS Patient Care STDS 2014, 28, 397–410. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Liu, W.; Zhu, M.; Sheng, J. Microbial Infections as a Trigger for Acute-on-Chronic Liver Failure: A Review. Med. Sci. Monit. 2019, 25, 4773–4783. [Google Scholar] [CrossRef]
- Luo, J.; Li, J.; Li, P.; Liang, X.; Hassan, H.M.; Moreau, R.; Li, J. Acute-on-chronic liver failure: Far to go—A review. Crit. Care 2023, 27, 259. [Google Scholar] [CrossRef]
- Sagnelli, E.; Coppola, N.; Messina, V.; Di Caprio, D.; Marrocco, C.; Marotta, A.; Onofrio, M.; Scolastico, C.; Filippini, P. HBV superinfection in hepatitis C virus chronic carriers, viral interaction, and clinical course. Hepatology 2002, 36, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Benvegnù, L.; Fattovich, G.; Noventa, F.; Tremolada, F.; Chemello, L.; Cecchetto, A.; Alberti, A. Concurrent hepatitis B and C virus infection and risk of hepatocellular carcinoma in cirrhosis. A prospective study. Cancer 1994, 74, 2442–2448. [Google Scholar] [CrossRef]
- Vento, S.; Garofano, T.; Renzini, C.; Cainelli, F.; Casali, F.; Ghironzi, G.; Ferraro, T.; Concia, E. Fulminant hepatitis associated with hepatitis A virus superinfection in patients with chronic hepatitis C. N. Engl. J. Med. 1998, 338, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Pramoolsinsap, C.; Poovorawan, Y.; Hirsch, P.; Busagorn, N.; Attamasirikul, K. Acute, hepatitis-A super-infection in HBV carriers, or chronic liver disease related to HBV or HCV. Ann. Trop. Med. Parasitol. 1999, 93, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Imai, K.; Petigara, T.; Kohn, M.A.; Nakashima, K.; Aoshima, M.; Shito, A.; Kanazu, S. Risk of pneumococcal diseases in adults with underlying medical conditions: A retrospective, cohort study using two Japanese healthcare databases. BMJ Open 2018, 8, e018553. [Google Scholar] [CrossRef] [PubMed]
- Schütte, A.; Ciesek, S.; Wedemeyer, H.; Lange, C.M. Influenza virus infection as precipitating event of acute-on-chronic liver failure. J. Hepatol. 2019, 70, 797–799. [Google Scholar] [CrossRef] [PubMed]
- Duchini, A.; Viernes, M.E.; Nyberg, L.M.; Hendry, R.M.; Pockros, P.J. Hepatic decompensation in patients with cirrhosis during infection with influenza A. Arch. Intern. Med. 2000, 160, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, G.N.; Liang, P.S.; Locke, E.; Green, P.; Berry, K.; O’Hare, A.M.; Shah, J.A.; Crothers, K.; Eastment, M.C.; Fan, V.S.; et al. Cirrhosis and Severe Acute Respiratory Syndrome Coronavirus 2 Infection in US Veterans: Risk of Infection, Hospitalization, Ventilation, and Mortality. Hepatology 2021, 74, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Sarin, S.K.; Choudhury, A.; Lau, G.K.; Zheng, M.H.; Ji, D.; Abd-Elsalam, S.; Hwang, J.; Qi, X.; Cua, I.H.; Suh, J.I.; et al. Pre-existing liver disease is associated with poor outcome in patients with SARS CoV2 infection; The APCOLIS Study (APASL COVID-19 Liver Injury Spectrum Study). Hepatol. Int. 2020, 14, 690–700. [Google Scholar] [CrossRef]
- Iavarone, M.; D’Ambrosio, R.; Soria, A.; Triolo, M.; Pugliese, N.; Del Poggio, P.; Perricone, G.; Massironi, S.; Spinetti, A.; Buscarini, E.; et al. High rates of 30-day mortality in patients with cirrhosis and COVID-19. J. Hepatol. 2020, 73, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, N.S.; Dhampalwar, S.; Saraf, N.; Soin, A.S. Outcomes of COVID-19 in Patients with Cirrhosis or Liver Transplantation. J. Clin. Exp. Hepatol. 2021, 11, 713–719. [Google Scholar] [CrossRef]
- Ballester, M.P.; Jalan, R.; Mehta, G. Vaccination in liver diseases and liver Transplantation: Recommendations, implications and opportunities in the post-covid era. JHEP Rep. 2023, 5, 100776. [Google Scholar] [CrossRef]
- Champion, C.R. Heplisav-B: A Hepatitis B Vaccine With a Novel Adjuvant. Ann. Pharmacother. 2021, 55, 783–791. [Google Scholar] [CrossRef]
- Di Lello, F.A.; Martínez, A.P.; Flichman, D.M. Insights into induction of the immune response by the hepatitis B vaccine. World J. Gastroenterol. 2022, 28, 4249–4262. [Google Scholar] [CrossRef]
- Aggeletopoulou, I.; Davoulou, P.; Konstantakis, C.; Thomopoulos, K.; Triantos, C. Response to hepatitis B vaccination in patients with liver cirrhosis. Rev. Med. Virol. 2017, 27, e1942. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Loucks, J.; Shah, M. Efficacy of Hepatitis B Vaccine in Adults with Chronic Liver Disease. J. Pharm. Pract. 2023, 36, 839–844. [Google Scholar] [CrossRef]
- Engler, S.H.; Sauer, P.W.; Golling, M.; Klar, E.A.; Benz, C.; Stremmel, W.; Kallinowski, B. Immunogenicity of two accelerated hepatitis B vaccination protocols in liver transplant candidates. Eur. J. Gastroenterol. Hepatol. 2001, 13, 363–367. [Google Scholar] [CrossRef]
- Arslan, M.; Wiesner, R.H.; Sievers, C.; Egan, K.; Zein, N.N. Double-dose accelerated hepatitis B vaccine in patients with end-stage liver disease. Liver Transpl. 2001, 7, 314–320. [Google Scholar] [CrossRef]
- Dhillon, S.; Moore, C.; Li, S.D.; Aziz, A.; Kakar, A.; Dosanjh, A.; Beesla, A.; Murphy, L.; Van Thiel, D.H. Efficacy of high-dose intra-dermal hepatitis B virus vaccine in previous vaccination non-responders with chronic liver disease. Dig. Dis. Sci. 2012, 57, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Arbizu, E.A.; Marugán, R.B.; Grijalba, J.Y.; Serrano, P.L.; Grande, L.G.; Del Campo Terrón, S. Intramuscular versus intradermal administration of anti-hepatitis B vaccine in non-cirrhotic hepatitis C patients. Vaccine 2003, 21, 2747–2750. [Google Scholar] [CrossRef] [PubMed]
- Amjad, W.; Alukal, J.; Zhang, T.; Maheshwari, A.; Thuluvath, P.J. Two-Dose Hepatitis B Vaccine (Heplisav-B) Results in Better Seroconversion Than Three-Dose Vaccine (Engerix-B) in Chronic Liver Disease. Dig. Dis. Sci. 2021, 66, 2101–2106. [Google Scholar] [CrossRef]
- Gutierrez Domingo, I.; Pascasio Acevedo, J.M.; Alcalde Vargas, A.; Ramos Cuadra, A.; Ferrer Ríos, M.T.; Sousa Martin, J.M.; Sayago Mota, M.; Giráldez Gallego, A.; Suárez Artacho, G. Response to vaccination against hepatitis B virus with a schedule of four 40-μg doses in cirrhotic patients evaluated for liver transplantation: Factors associated with a response. Transplant. Proc. 2012, 44, 1499–1501. [Google Scholar] [CrossRef]
- Roni, D.A.; Pathapati, R.M.; Kumar, A.S.; Nihal, L.; Sridhar, K.; Tumkur Rajashekar, S. Safety and efficacy of hepatitis B vaccination in cirrhosis of liver. Adv. Virol. 2013, 2013, 196704. [Google Scholar] [CrossRef]
- Giráldez-Gallego, Á.; Rodríguez-Seguel, E.D.P.; Valencia-Martín, R.; Morillo-García, Á.; Salamanca-Rivera, C.; Ruiz-Pérez, R.; Cuaresma-Duque, M.; Rosso-Fernández, C.; Ferrer-Ríos, M.T.; Sousa-Martín, J.M.; et al. Three double-dose reinforced hepatitis B revaccination scheme for patients with cirrhosis unresponsive to the standard regimen: An open-label randomised clinical trial. Gut 2023, 73, 166–174. [Google Scholar] [CrossRef]
- Keeffe, E.B.; Iwarson, S.; McMahon, B.J.; Lindsay, K.L.; Koff, R.S.; Manns, M.; Baumgarten, R.; Wiese, M.; Fourneau, M.; Safary, A.; et al. Safety and immunogenicity of hepatitis A vaccine in patients with chronic liver disease. Hepatology 1998, 27, 881–886. [Google Scholar] [CrossRef]
- Arguedas, M.R.; Johnson, A.; Eloubeidi, M.A.; Fallon, M.B. Immunogenicity of hepatitis A vaccination in decompensated cirrhotic patients. Hepatology 2001, 34, 28–31. [Google Scholar] [CrossRef]
- Pirovino, M.; Lydick, E.; Grob, P.J.; Arrenbrecht, S.; Altorfer, J.; Schmid, M. Pneumococcal vaccination: The response of patients with alcoholic liver cirrhosis. Hepatology 1984, 4, 946–949. [Google Scholar] [CrossRef] [PubMed]
- Preheim, L.C.; Mellencamp, M.A.; Snitily, M.U.; Gentry, M.J. Effect of cirrhosis on the production and efficacy of pneumococcal capsular antibody in a rat model. Am. Rev. Respir. Dis. 1992, 146, 1054–1058. [Google Scholar] [CrossRef]
- McCashland, T.M.; Preheim, L.C.; Gentry, M.J. Pneumococcal vaccine response in cirrhosis and liver transplantation. J. Infect. Dis. 2000, 181, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Härmälä, S.; Parisinos, C.A.; Shallcross, L.; O’Brien, A.; Hayward, A. Effectiveness of influenza vaccines in adults with chronic liver disease: A systematic review and meta-analysis. BMJ Open 2019, 9, e031070. [Google Scholar] [CrossRef] [PubMed]
- Gaeta, G.B.; Stornaiuolo, G.; Precone, D.F.; Amendola, A.; Zanetti, A.R. Immunogenicity and safety of an adjuvanted influenza vaccine in patients with decompensated cirrhosis. Vaccine 2002, 20 (Suppl. 5), B33–B35. [Google Scholar] [CrossRef] [PubMed]
- Thuluvath, P.J.; Robarts, P.; Chauhan, M. Analysis of antibody responses after COVID-19 vaccination in liver transplant recipients and those with chronic liver diseases. J. Hepatol. 2021, 75, 1434–1439. [Google Scholar] [CrossRef] [PubMed]
- Bakasis, A.D.; Bitzogli, K.; Mouziouras, D.; Pouliakis, A.; Roumpoutsou, M.; Goules, A.V.; Androutsakos, T. Antibody Responses after SARS-CoV-2 Vaccination in Patients with Liver Diseases. Viruses 2022, 14, 207. [Google Scholar] [CrossRef]
- Iavarone, M.; Tosetti, G.; Facchetti, F.; Topa, M.; Er, J.M.; Hang, S.K.; Licari, D.; Lombardi, A.; D’Ambrosio, R.; Degasperi, E.; et al. Spike-specific humoral and cellular immune responses after COVID-19 mRNA vaccination in patients with cirrhosis: A prospective single center study. Dig. Liver Dis. 2023, 55, 160–168. [Google Scholar] [CrossRef]
- Beran, A.; Mhanna, A.; Mhanna, M.; Hassouneh, R.; Abuhelwa, Z.; Mohamed, M.F.H.; Sayeh, W.; Musallam, R.; Assaly, R.; Abdeljawad, K. Real-world effectiveness of COVID-19 vaccination in liver cirrhosis: A systematic review with meta-analysis of 51,834 patients. Proceedings (Bayl. Univ. Med. Cent.) 2023, 36, 151–156. [Google Scholar] [CrossRef]
- Loulergue, P.; Pol, S.; Mallet, V.; Sogni, P.; Launay, O.; GEVACCIM Group. Why actively promote vaccination in patients with cirrhosis? J. Clin. Virol. 2009, 46, 206–209. [Google Scholar] [CrossRef]
- Casella, G.; Ingravalle, F.; Ingravalle, A.; Andreotti, S.; Bonetti, F.; Monti, C.; Falbo, R.; Rumi, M.G. Approaches for Selective Vaccinations in Cirrhotic Patients. Vaccines 2023, 11, 460. [Google Scholar] [CrossRef]
- Jacobs, R.J.; Meyerhoff, A.S.; Saab, S. Immunization needs of chronic liver disease patients seen in primary care versus specialist settings. Dig. Dis. Sci. 2005, 50, 1525–1531. [Google Scholar] [CrossRef] [PubMed]
- Advisory Committee on Immunization Practices (ACIP); Fiore, A.E.; Wasley, A.; Bell, B.P. Prevention of hepatitis A through active or passive immunization: Recommendations of the Advisory Committee on Immunization Practices (ACIP). MMWR Recomm. Rep. 2006, 55, 1–23. [Google Scholar] [PubMed]
- Murthy, N.; Wodi, A.P.; McNally, V.; Cineas, S.; Ault, K. Advisory Committee on Immunization Practices Recommended Immunization Schedule for Adults Aged 19 Years or Older—United States, 2023. MMWR Morb. Mortal. Wkly. Rep. 2023, 72, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Loulergue, P.; Launay, O.; le groupe GEVACCIM (Groupe d’Etude sur la Vaccination des Sujets Immunodéprimés). Vaccinations chez les patients ayant une cirrhose [Vaccination in patients with cirrhosis]. Presse Med. 2009, 38, 1134–1140. [Google Scholar] [CrossRef]
- Song, J.Y.; Cheong, H.J.; Ha, S.H.; Hwang, I.S.; Kee, S.Y.; Jeong, H.W.; Lee, C.G.; Kim, W.J. Clinical impact of influenza immunization in patients with liver cirrhosis. J. Clin. Virol. 2007, 39, 159–163. [Google Scholar] [CrossRef]
- Mast, E.E.; Margolis, H.S.; Fiore, A.E.; Brink, E.W.; Goldstein, S.T.; Wang, S.A.; Moyer, L.A.; Bell, B.P.; Alter, M.J.; Advisory Committee on Immunization Practices (ACIP). A comprehensive immunization strategy to eliminate transmission of hepatitis B virus infection in the United States: Recommendations of the Advisory Committee on Immunization Practices (ACIP) part 1: Immunization of infants, children, and adolescents. MMWR Recomm. Rep. 2005, 54, 1–31, Erratum in MMWR Morb. Mortal. Wkly. Rep. 2006, 55, 158–159; Erratum in MMWR Morb. Mortal. Wkly. Rep. 2007, 56, 1267. [Google Scholar]
- Granito, A.; Muratori, P.; Muratori, L. Acute-on-chronic liver failure: A complex clinical entity in patients with autoimmune hepatitis. J. Hepatol. 2021, 75, 1503–1505. [Google Scholar] [CrossRef]
- Al-Dury, S.; Kanberg, N. Advancements in Vaccine Strategies for Chronic Liver Disease Patients: Navigating Post-COVID Challenges and Opportunities. Vaccines 2024, 12, 197. [Google Scholar] [CrossRef] [PubMed]
- Leise, M.D.; Talwalkar, J.A. Immunizations in chronic liver disease: What should be done and what is the evidence. Curr. Gastroenterol. Rep. 2013, 15, 300. [Google Scholar] [CrossRef] [PubMed]
- Alukal, J.J.; Naqvi, H.A.; Thuluvath, P.J. Vaccination in Chronic Liver Disease: An Update. J. Clin. Exp. Hepatol. 2022, 12, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, M.; Bárcena, R.; García, M.; López-Sanroman, A.; Nuño, J. Vaccination against hepatitis B virus in cirrhotic patients on liver transplant waiting list. Liver Transpl. 2000, 6, 440–442. [Google Scholar] [CrossRef]
- Stark, K.; Günther, M.; Neuhaus, R.; Reinke, P.; Schröder, K.; Linnig, S.; Bienzle, U. Immunogenicity and safety of hepatitis A vaccine in liver and renal transplant recipients. J. Infect. Dis. 1999, 180, 2014–2017. [Google Scholar] [CrossRef] [PubMed]
- Burbach, G.; Bienzle, U.; Stark, K.; Rayes, N.; Neuhaus, R.; Serke, S.; Engelmann, H.; Künzel, W.; Türk, G.; Neuhaus, P. Influenza vaccination in liver transplant recipients. Transplantation 1999, 67, 753–755. [Google Scholar] [CrossRef] [PubMed]
- Vilchez, R.A.; Fung, J.J.; Kusne, S. Influenza A myocarditis developing in an adult liver transplant recipient despite vaccination: A case report and review of the literature. Transplantation 2000, 70, 543–545. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, B.L.; Pellicane, A.J.; Tyring, S.K. Vaccine immunology. Dermatol. Ther. 2009, 22, 104–109. [Google Scholar] [CrossRef]
- Soto, J.A.; Díaz, F.E.; Retamal-Díaz, A.; Gálvez, N.M.S.; Melo-González, F.; Piña-Iturbe, A.; Ramírez, M.A.; Bohmwald, K.; González, P.A.; Bueno, S.M.; et al. BCG-Based Vaccines Elicit Antigen-Specific Adaptive and Trained Immunity against SARS-CoV-2 and Andes orthohantavirus. Vaccines 2022, 10, 721. [Google Scholar] [CrossRef]
- Sancho, D.; Gómez, M.; Sánchez-Madrid, F. CD69 is an immunoregulatory molecule induced following activation. Trends Immunol. 2005, 26, 136–140. [Google Scholar] [CrossRef]
- Lauzurica, P.; Sancho, D.; Torres, M.; Albella, B.; Marazuela, M.; Merino, T.; Bueren, J.A.; Martínez, A.C.; Sánchez-Madrid, F. Phenotypic and functional characteristics of hematopoietic cell lineages in CD69-deficient mice. Blood 2000, 95, 2312–2320. [Google Scholar] [CrossRef]
- Reif, K.; Ekland, E.H.; Ohl, L.; Nakano, H.; Lipp, M.; Förster, R.; Cyster, J.G. Balanced responsiveness to chemoattractants from adjacent zones determines B-cell position. Nature 2002, 416, 94–99. [Google Scholar] [CrossRef]
- Palm, A.E.; Henry, C. Remembrance of Things Past: Long-Term B Cell Memory After Infection and Vaccination. Front. Immunol. 2019, 10, 1787. [Google Scholar] [CrossRef]
- Gilbert, S.C. T-cell-inducing vaccines—What’s the future. Immunology 2012, 135, 19–26. [Google Scholar] [CrossRef]
- Heinzel, F.P. Infections in patients with humoral immunodeficiency. Hosp. Pract. (Off. Ed.) 1989, 24, 99–103, 106–111. [Google Scholar] [CrossRef]
- Gershon, A.A.; Breuer, J.; Cohen, J.I.; Cohrs, R.J.; Gershon, M.D.; Gilden, D.; Grose, C.; Hambleton, S.; Kennedy, P.G.; Oxman, M.N.; et al. Varicella zoster virus infection. Nat. Rev. Dis. Primers 2015, 1, 15016. [Google Scholar] [CrossRef] [PubMed]
- Wherry, E.J.; Barouch, D.H. T cell immunity to COVID-19 vaccines. Science 2022, 377, 821–822. [Google Scholar] [CrossRef] [PubMed]
- Malley, R.; Trzcinski, K.; Srivastava, A.; Thompson, C.M.; Anderson, P.W.; Lipsitch, M. CD4+ T cells mediate antibody-independent acquired immunity to pneumococcal colonization. Proc. Natl. Acad. Sci. USA 2005, 102, 4848–4853. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.A.; Nambiar, J.K.; Wozniak, T.M.; Roediger, B.; Shklovskaya, E.; Britton, W.J.; Fazekas de St Groth, B.; Triccas, J.A. Antigen load governs the differential priming of CD8 T cells in response to the bacille Calmette Guerin vaccine or Mycobacterium tuberculosis infection. J. Immunol. 2009, 182, 7172–7177. [Google Scholar] [CrossRef] [PubMed]
- Goel, R.R.; Painter, M.M.; Apostolidis, S.A.; Mathew, D.; Meng, W.; Rosenfeld, A.M.; Lundgreen, K.A.; Reynaldi, A.; Khoury, D.S.; Pattekar, A.; et al. mRNA vaccines induce durable immune memory to SARS-CoV-2 and variants of concern. Science 2021, 374, abm0829. [Google Scholar] [CrossRef]
- Echaide, M.; Chocarro de Erauso, L.; Bocanegra, A.; Blanco, E.; Kochan, G.; Escors, D. mRNA Vaccines against SARS-CoV-2: Advantages and Caveats. Int. J. Mol. Sci. 2023, 24, 5944. [Google Scholar] [CrossRef] [PubMed]
- Dogan, I.; Bertocci, B.; Vilmont, V.; Delbos, F.; Mégret, J.; Storck, S.; Reynaud, C.A.; Weill, J.C. Multiple layers of B cell memory with different effector functions. Nat. Immunol. 2009, 10, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Mei, H.; Dörner, T.; Hiepe, F.; Radbruch, A.; Fillatreau, S.; Hoyer, B.F. Memory B and memory plasma cells. Immunol. Rev. 2010, 237, 117–139. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, J.; Bos, N.A.; Kroese, F.G.M. Heterogeneity of Memory Marginal Zone B Cells. Crit. Rev. Immunol. 2018, 38, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Kardava, L.; Rachmaninoff, N.; Lau, W.W.; Buckner, C.M.; Trihemasava, K.; Blazkova, J.; Lopes de Assis, F.; Wang, W.; Zhang, X.; Wang, Y.; et al. Early human B cell signatures of the primary antibody response to mRNA vaccination. Proc. Natl. Acad. Sci. USA 2022, 119, e2204607119. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Babor, M.; Lane, J.; Schulten, V.; Patil, V.S.; Seumois, G.; Rosales, S.L.; Fu, Z.; Picarda, G.; Burel, J.; et al. Unique phenotypes and clonal expansions of human CD4 effector memory T cells re-expressing CD45RA. Nat. Commun. 2017, 8, 1473. [Google Scholar] [CrossRef] [PubMed]
- Wilk, M.M.; Mills, K.H.G. CD4 TRM Cells Following Infection and Immunization: Implications for More Effective Vaccine Design. Front. Immunol. 2018, 9, 1860. [Google Scholar] [CrossRef] [PubMed]
- Takamura, S. Niches for the Long-Term Maintenance of Tissue-Resident Memory T Cells. Front. Immunol. 2018, 9, 1214. [Google Scholar] [CrossRef]
- Hassert, M.; Harty, J.T. Tissue resident memory T cells—A new benchmark for the induction of vaccine-induced mucosal immunity. Front. Immunol. 2022, 13, 1039194. [Google Scholar] [CrossRef]
- Rotrosen, E.; Kupper, T.S. Assessing the generation of tissue resident memory T cells by vaccines. Nat. Rev. Immunol. 2023, 23, 655–665. [Google Scholar] [CrossRef]
- Lumsden, J.M.; Schwenk, R.J.; Rein, L.E.; Moris, P.; Janssens, M.; Ofori-Anyinam, O.; Cohen, J.; Kester, K.E.; Heppner, D.G.; Krzych, U. Protective immunity induced with the RTS,S/AS vaccine is associated with IL-2 and TNF-α producing effector and central memory CD4 T cells. PLoS ONE 2011, 6, e20775. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.I.; Westerhof, L.M.; MacLeod, M.K.L. The roles of resident, central and effector memory CD4 T-cells in protective immunity following infection or vaccination. Immunology 2018, 154, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Vinuesa, C.G.; Linterman, M.A.; Yu, D.; MacLennan, I.C. Follicular Helper T Cells. Annu. Rev. Immunol. 2016, 34, 335–368. [Google Scholar] [CrossRef] [PubMed]
- Crotty, S. A brief history of T cell help to B cells. Nat. Rev. Immunol. 2015, 15, 185–189. [Google Scholar] [CrossRef]
- Patente, T.A.; Pinho, M.P.; Oliveira, A.A.; Evangelista, G.C.M.; Bergami-Santos, P.C.; Barbuto, J.A.M. Human Dendritic Cells: Their Heterogeneity and Clinical Application Potential in Cancer Immunotherapy. Front. Immunol. 2019, 9, 3176. [Google Scholar] [CrossRef]
- Heesters, B.A.; Myers, R.C.; Carroll, M.C. Follicular dendritic cells: Dynamic antigen libraries. Nat. Rev. Immunol. 2014, 14, 495–504. [Google Scholar] [CrossRef]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef]
- Segura, E. Cross-Presentation Assay for Human Dendritic Cells. Methods Mol. Biol. 2016, 1423, 189–198. [Google Scholar] [CrossRef]
- Xu, R.H.; Remakus, S.; Ma, X.; Roscoe, F.; Sigal, L.J. Direct presentation is sufficient for an efficient anti-viral CD8+ T cell response. PLoS Pathog. 2010, 6, e1000768. [Google Scholar] [CrossRef]
- Badovinac, V.P.; Messingham, K.A.; Jabbari, A.; Haring, J.S.; Harty, J.T. Accelerated CD8+ T-cell memory and prime-boost response after dendritic-cell vaccination. Nat. Med. 2005, 11, 748–756. [Google Scholar] [CrossRef]
- Querec, T.; Bennouna, S.; Alkan, S.; Laouar, Y.; Gorden, K.; Flavell, R.; Akira, S.; Ahmed, R.; Pulendran, B. Yellow fever vaccine YF-17D activates multiple dendritic cell subsets via TLR2, 7, 8, and 9 to stimulate polyvalent immunity. J. Exp. Med. 2006, 203, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B.S.; Arunachalam, P.; O’Hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef] [PubMed]
- Brandtzaeg, P. Function of mucosa-associated lymphoid tissue in antibody formation. Immunol. Investig. 2010, 39, 303–355. [Google Scholar] [CrossRef] [PubMed]
- Lycke, N. Recent progress in mucosal vaccine development: Potential and limitations. Nat. Rev. Immunol. 2012, 12, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.H.; Kim, S.H.; Jeon, J.H.; Kim, E.B.; Lee, N.K.; Beck, S.; Choi, Y.J.; Kang, S.K. Cytoplasmic expression of a model antigen with M Cell-Targeting moiety in lactic acid bacteria and implication of the mechanism as a mucosal vaccine via oral route. Vaccine 2021, 39, 4072–4081. [Google Scholar] [CrossRef] [PubMed]
- Lund, F.E.; Randall, T.D. Effector and regulatory B cells: Modulators of CD4+ T cell immunity. Nat. Rev. Immunol. 2010, 10, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, M.; Du, G.; Chen, X.; Sun, X. Advanced oral vaccine delivery strategies for improving the immunity. Adv. Drug Deliv. Rev. 2021, 177, 113928. [Google Scholar] [CrossRef] [PubMed]
- Kunisawa, J.; Kurashima, Y.; Kiyono, H. Gut-associated lymphoid tissues for the development of oral vaccines. Adv. Drug Deliv. Rev. 2012, 64, 523–530. [Google Scholar] [CrossRef]
- Robbins, J.B.; Schneerson, R.; Szu, S.C. Perspective: Hypothesis: Serum IgG antibody is sufficient to confer protection against infectious diseases by inactivating the inoculum. J. Infect. Dis. 1995, 171, 1387–1398. [Google Scholar] [CrossRef]
- Kwong, K.W.; Xin, Y.; Lai, N.C.; Sung, J.C.; Wu, K.C.; Hamied, Y.K.; Sze, E.T.; Lam, D.M. Oral Vaccines: A Better Future of Immunization. Vaccines 2023, 11, 1232. [Google Scholar] [CrossRef]
- Albillos, A.; Lario, M.; Álvarez-Mon, M. Cirrhosis-associated immune dysfunction: Distinctive features and clinical relevance. J. Hepatol. 2014, 61, 1385–1396. [Google Scholar] [CrossRef]
- Albillos, A.; Martin-Mateos, R.; Van der Merwe, S.; Wiest, R.; Jalan, R.; Álvarez-Mon, M. Cirrhosis-associated immune dysfunction. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 112–134. [Google Scholar] [CrossRef]
- Simão, A.L.; Palma, C.S.; Izquierdo-Sanchez, L.; Putignano, A.; Carvalho-Gomes, A.; Posch, A.; Zanaga, P.; Girleanu, I.; Henrique, M.M.; Araújo, C.; et al. Cirrhosis is associated with lower serological responses to COVID-19 vaccines in patients with chronic liver disease. JHEP Rep. 2023, 5, 100697. [Google Scholar] [CrossRef]
- Regev-Yochay, G.; Lustig, Y.; Joseph, G.; Gilboa, M.; Barda, N.; Gens, I.; Indenbaum, V.; Halpern, O.; Katz-Likvornik, S.; Levin, T.; et al. Correlates of protection against COVID-19 infection and intensity of symptomatic disease in vaccinated individuals exposed to SARS-CoV-2 in households in Israel (ICoFS): A prospective cohort study. Lancet Microbe 2023, 4, e309–e318. [Google Scholar] [CrossRef]
- Plotkin, S.A. Correlates of protection induced by vaccination. Clin. Vaccine Immunol. 2010, 17, 1055–1065. [Google Scholar] [CrossRef] [PubMed]
- Salzer, U.; Sack, U.; Fuchs, I. Flow Cytometry in the Diagnosis and Follow Up of Human Primary Immunodeficiencies. EJIFCC 2019, 30, 407–422. [Google Scholar] [PubMed]
- De Milito, A.; Mörch, C.; Sönnerborg, A.; Chiodi, F. Loss of memory (CD27) B lymphocytes in HIV-1 infection. AIDS 2001, 15, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Doi, H.; Iyer, T.K.; Carpenter, E.; Li, H.; Chang, K.M.; Vonderheide, R.H.; Kaplan, D.E. Dysfunctional B-cell activation in cirrhosis resulting from hepatitis C infection associated with disappearance of CD27-positive B-cell population. Hepatology 2012, 55, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Jhun, J.Y.; Kim, H.Y.; Byun, J.K.; Chung, B.H.; Bae, S.H.; Yoon, S.K.; Kim, D.G.; Yang, C.W.; Cho, M.L.; Choi, J.Y. B-cell-associated immune profiles in patients with decompensated cirrhosis. Scand. J. Gastroenterol. 2015, 50, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Doi, H.; Hayashi, E.; Arai, J.; Tojo, M.; Morikawa, K.; Eguchi, J.; Ito, T.; Kanto, T.; Kaplan, D.E.; Yoshida, H. Enhanced B-cell differentiation driven by advanced cirrhosis resulting in hyperglobulinemia. J. Gastroenterol. Hepatol. 2018, 33, 1667–1676. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.T.; Jing, Y.Y.; Han, Z.P.; Li, X.N.; Liu, Y.; Lai, F.B.; Li, R.; Zhao, Q.D.; Wu, M.C.; Wei, L.X. The injured liver induces hyperimmunoglobulinemia by failing to dispose of antigens and endotoxins in the portal system. PLoS ONE 2015, 10, e0122739. [Google Scholar] [CrossRef]
- Abdelwahab, F.A.; Hassanein, K.M.; Hetta, H.F.; Abdelmalek, M.O.; Zahran, A.M.; El-Badawy, O. Impact of deranged B cell subsets distribution in the development of HCV-related cirrhosis and HCC in type two diabetes mellitus. Sci. Rep. 2020, 10, 20383. [Google Scholar] [CrossRef]
- Xiong, Y.; Wu, H.; Li, Y.; Huang, R.; Liu, Y.; Chen, Y.; Zhao, X.; Chang, H.; Chen, J.; Wu, C. Characteristics of peripheral and intrahepatic regulatory B cells in HBV-related liver cirrhosis. Int. J. Clin. Exp. Pathol. 2018, 11, 4545–4551. [Google Scholar]
- Huang, M.; Liu, X.; Ye, H.; Zhao, X.; Zhao, J.; Liu, Y.; He, X.; Qu, M.; Pan, J.; Hou, B.; et al. Metabolic defects in splenic B cell compartments from patients with liver cirrhosis. Cell Death Dis. 2020, 11, 915. [Google Scholar] [CrossRef]
- Seifert, M.; Przekopowitz, M.; Taudien, S.; Lollies, A.; Ronge, V.; Drees, B.; Lindemann, M.; Hillen, U.; Engler, H.; Singer, B.B.; et al. Functional capacities of human IgM memory B cells in early inflammatory responses and secondary germinal center reactions. Proc. Natl. Acad. Sci. USA 2015, 112, E546–E555. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Shi, J.; Qu, M.; Zhao, X.; Wang, H.; Huang, M.; Liu, Z.; Li, Z.; He, Q.; Zhang, S.; et al. Hyperactive Follicular Helper T Cells Contribute to Dysregulated Humoral Immunity in Patients With Liver Cirrhosis. Front. Immunol. 2019, 10, 1915. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.Y.; Feng, B.; Zhang, H.H.; Rao, H.Y.; Wang, J.H.; Cong, X.; Wei, L. CD4+CXCR5+ T cells activate CD27+IgG+ B cells via IL-21 in patients with hepatitis C virus infection. Hepatobiliary Pancreat. Dis. Int. 2016, 15, 55–64. [Google Scholar] [CrossRef]
- Noor, M.T.; Manoria, P. Immune Dysfunction in Cirrhosis. J. Clin. Transl. Hepatol. 2017, 5, 50–58. [Google Scholar] [CrossRef]
- Hackstein, C.P.; Spitzer, J.; Symeonidis, K.; Horvatic, H.; Bedke, T.; Steglich, B.; Klein, S.; Assmus, L.M.; Odainic, A.; Szlapa, J.; et al. Interferon-induced IL-10 drives systemic T-cell dysfunction during chronic liver injury. J. Hepatol. 2023, 79, 150–166. [Google Scholar] [CrossRef]
- Lebossé, F.; Gudd, C.; Tunc, E.; Singanayagam, A.; Nathwani, R.; Triantafyllou, E.; Pop, O.; Kumar, N.; Mukherjee, S.; Hou, T.Z.; et al. CD8+ T cells from patients with cirrhosis display a phenotype that may contribute to cirrhosis-associated immune dysfunction. EBioMedicine 2019, 49, 258–268. [Google Scholar] [CrossRef]
- Shive, C.L.; Kowal, C.M.; Desotelle, A.F.; Nguyen, Y.; Carbone, S.; Kostadinova, L.; Davitkov, P.; O’Mara, M.; Reihs, A.; Siddiqui, H.; et al. Endotoxemia Associated with Liver Disease Correlates with Systemic Inflammation and T Cell Exhaustion in Hepatitis C Virus Infection. Cells 2023, 12, 2034. [Google Scholar] [CrossRef]
- Lario, M.; Muñoz, L.; Ubeda, M.; Borrero, M.J.; Martínez, J.; Monserrat, J.; Díaz, D.; Alvarez-Mon, M.; Albillos, A. Defective thymopoiesis and poor peripheral homeostatic replenishment of T-helper cells cause T-cell lymphopenia in cirrhosis. J. Hepatol. 2013, 59, 723–730. [Google Scholar] [CrossRef]
- John, B.V.; Deng, Y.; Schwartz, K.B.; Taddei, T.H.; Kaplan, D.E.; Martin, P.; Chao, H.H.; Dahman, B. Postvaccination COVID-19 infection is associated with reduced mortality in patients with cirrhosis. Hepatology 2022, 76, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Boulouis, C.; Kammann, T.; Cuapio, A.; Parrot, T.; Gao, Y.; Mouchtaridi, E.; Wullimann, D.; Lange, J.; Chen, P.; Akber, M.; et al. MAIT cell compartment characteristics are associated with the immune response magnitude to the BNT162b2 mRNA anti-SARS-CoV-2 vaccine. Mol. Med. 2022, 28, 54. [Google Scholar] [CrossRef] [PubMed]
- Niehaus, C.E.; Strunz, B.; Cornillet, M.; Falk, C.S.; Schnieders, A.; Maasoumy, B.; Hardtke, S.; Manns, M.P.; Kraft, A.R.M.; Björkström, N.K.; et al. MAIT Cells Are Enriched and Highly Functional in Ascites of Patients With Decompensated Liver Cirrhosis. Hepatology 2020, 72, 1378–1393. [Google Scholar] [CrossRef] [PubMed]
- Hegde, P.; Weiss, E.; Paradis, V.; Wan, J.; Mabire, M.; Sukriti, S.; Rautou, P.E.; Albuquerque, M.; Picq, O.; Gupta, A.C.; et al. Mucosal-associated invariant T cells are a profibrogenic immune cell population in the liver. Nat. Commun. 2018, 9, 2146. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Chouik, Y.; Lebossé, F.; Khamri, W. Dysfunctions of Circulating Adaptive Immune Cells in End-Stage Liver Disease. Livers 2023, 3, 369–382. [Google Scholar] [CrossRef]
- Cardoso, C.C.; Matiollo, C.; Pereira, C.H.J.; Fonseca, J.S.; Alves, H.E.L.; da Silva, O.M.; de Souza Menegassi, V.; Dos Santos, C.R.; de Moraes, A.C.R.; de Lucca Schiavon, L.; et al. Patterns of dendritic cell and monocyte subsets are associated with disease severity and mortality in liver cirrhosis patients. Sci. Rep. 2021, 11, 5923. [Google Scholar] [CrossRef] [PubMed]
- Kakazu, E.; Kondo, Y.; Kogure, T.; Ninomiya, M.; Kimura, O.; Ueno, Y.; Shimosegawa, T. Plasma amino acids imbalance in cirrhotic patients disturbs the tricarboxylic acid cycle of dendritic cell. Sci. Rep. 2013, 3, 3459. [Google Scholar] [CrossRef]
- Della Bella, S.; Crosignani, A.; Riva, A.; Presicce, P.; Benetti, A.; Longhi, R.; Podda, M.; Villa, M.L. Decrease and dysfunction of dendritic cells correlate with impaired hepatitis C virus-specific CD4+ T-cell proliferation in patients with hepatitis C virus infection. Immunology 2007, 121, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Crosignani, A.; Riva, A.; Della Bella, S. Analysis of peripheral blood dendritic cells as a non-invasive tool in the follow-up of patients with chronic hepatitis C. World J. Gastroenterol. 2016, 22, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- Ouaguia, L.; Leroy, V.; Dufeu-Duchesne, T.; Durantel, D.; Decaens, T.; Hubert, M.; Valladeau-Guilemond, J.; Bendriss-Vermare, N.; Chaperot, L.; Aspord, C. Circulating and Hepatic BDCA1+, BDCA2+, and BDCA3+ Dendritic Cells Are Differentially Subverted in Patients With Chronic HBV Infection. Front. Immunol. 2019, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Irla, M.; Küpfer, N.; Suter, T.; Lissilaa, R.; Benkhoucha, M.; Skupsky, J.; Lalive, P.H.; Fontana, A.; Reith, W.; Hugues, S. MHC class II-restricted antigen presentation by plasmacytoid dendritic cells inhibits T cell-mediated autoimmunity. J. Exp. Med. 2010, 207, 1891–1905. [Google Scholar] [CrossRef] [PubMed]
- Oh, N.A.; O’Shea, T.; Ndishabandi, D.K.; Yuan, Q.; Hong, S.; Gans, J.; Ge, J.; Gibney, S.; Chase, C.; Yang, C.; et al. Plasmacytoid Dendritic Cell-driven Induction of Treg Is Strain Specific and Correlates With Spontaneous Acceptance of Kidney Allografts. Transplantation 2020, 104, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Mazariegos, G.V.; Zahorchak, A.F.; Reyes, J.; Ostrowski, L.; Flynn, B.; Zeevi, A.; Thomson, A.W. Dendritic cell subset ratio in peripheral blood correlates with successful withdrawal of immunosuppression in liver transplant patients. Am. J. Transplant. 2003, 3, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Rogers, N.M.; Isenberg, J.S.; Thomson, A.W. Plasmacytoid dendritic cells: No longer an enigma and now key to transplant tolerance? Am. J. Transplant. 2013, 13, 1125–1133. [Google Scholar] [CrossRef]
- Brezar, V.; Godot, V.; Cheng, L.; Su, L.; Lévy, Y.; Seddiki, N. T-Regulatory Cells and Vaccination “Pay Attention and Do Not Neglect Them”: Lessons from HIV and Cancer Vaccine Trials. Vaccines 2016, 4, 30. [Google Scholar] [CrossRef]
- Fang, W.N.; Shi, M.; Meng, C.Y.; Li, D.D.; Peng, J.P. The Balance between Conventional DCs and Plasmacytoid DCs Is Pivotal for Immunological Tolerance during Pregnancy in the Mouse. Sci. Rep. 2016, 6, 26984. [Google Scholar] [CrossRef]
- Musumeci, A.; Lutz, K.; Winheim, E.; Krug, A.B. What Makes a pDC: Recent Advances in Understanding Plasmacytoid DC Development and Heterogeneity. Front. Immunol. 2019, 10, 1222. [Google Scholar] [CrossRef]
- Lee, G.H.; Lim, S.G. CpG-Adjuvanted Hepatitis B Vaccine (HEPLISAV-B®) Update. Expert. Rev. Vaccines 2021, 20, 487–495. [Google Scholar] [CrossRef]
- Nieto, J.C.; Perea, L.; Soriano, G.; Zamora, C.; Cantó, E.; Medina, A.; Poca, M.; Sanchez, E.; Roman, E.; Julià, G.; et al. Ascitic fluid regulates the local innate immune response of patients with cirrhosis. J. Leukoc. Biol. 2018, 104, 833–841. [Google Scholar] [CrossRef]
- Muñoz, L.; Caparrós, E.; Albillos, A.; Francés, R. The shaping of gut immunity in cirrhosis. Front. Immunol. 2023, 14, 1139554. [Google Scholar] [CrossRef]
- Alter, G.; Sekaly, R.P. Beyond adjuvants: Antagonizing inflammation to enhance vaccine immunity. Vaccine 2015, 33 (Suppl. 2), B55–B59. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, G.; Briantais, M.J.; Buffet, C.; Pillot, J.; Etienne, J.P. Serum and intestinal secretory IgA in alcoholic cirrhosis of the liver. Gut 1982, 23, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, V.; Angeli, P.; Moreau, R.; Jalan, R.; Clària, J.; Trebicka, J.; Fernández, J.; Gustot, T.; Caraceni, P.; Bernardi, M.; et al. The systemic inflammation hypothesis: Towards a new paradigm of acute decompensation and multiorgan failure in cirrhosis. J. Hepatol. 2021, 74, 670–685. [Google Scholar] [CrossRef]
- Wilde, B.; Katsounas, A. Immune Dysfunction and Albumin-Related Immunity in Liver Cirrhosis. Mediat. Inflamm. 2019, 2019, 7537649. [Google Scholar] [CrossRef]
- Chen, D.; Le, T.H.; Shahidipour, H.; Read, S.A.; Ahlenstiel, G. The Role of Gut-Derived Microbial Antigens on Liver Fibrosis Initiation and Progression. Cells 2019, 8, 1324. [Google Scholar] [CrossRef] [PubMed]
- Schulthess, J.; Pandey, S.; Capitani, M.; Rue-Albrecht, K.C.; Arnold, I.; Franchini, F.; Chomka, A.; Ilott, N.E.; Johnston, D.G.W.; Pires, E.; et al. The Short Chain Fatty Acid Butyrate Imprints an Antimicrobial Program in Macrophages. Immunity 2019, 50, 432–445. [Google Scholar] [CrossRef]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef]
- Burger-van Paassen, N.; Vincent, A.; Puiman, P.J.; van der Sluis, M.; Bouma, J.; Boehm, G.; van Goudoever, J.B.; van Seuningen, I.; Renes, I.B. The regulation of intestinal mucin MUC2 expression by short-chain fatty acids: Implications for epithelial protection. Biochem. J. 2009, 420, 211–219. [Google Scholar] [CrossRef]
- Jin, M.; Kalainy, S.; Baskota, N.; Chiang, D.; Deehan, E.C.; McDougall, C.; Tandon, P.; Martínez, I.; Cervera, C.; Walter, J.; et al. Faecal microbiota from patients with cirrhosis has a low capacity to ferment non-digestible carbohydrates into short-chain fatty acids. Liver Int. 2019, 39, 1437–1447. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, C.; Zuo, S.; Cao, K.; Li, H. Integrative analysis of the gut microbiota and faecal and serum short-chain fatty acids and tryptophan metabolites in patients with cirrhosis and hepatic encephalopathy. J. Transl. Med. 2023, 21, 395. [Google Scholar] [CrossRef]
- Kim, M.; Qie, Y.; Park, J.; Kim, C.H. Gut Microbial Metabolites Fuel Host Antibody Responses. Cell Host Microbe 2016, 20, 202–214. [Google Scholar] [CrossRef] [PubMed]
- Sauerbruch, T.; Hennenberg, M.; Trebicka, J.; Beuers, U. Bile Acids, Liver Cirrhosis, and Extrahepatic Vascular Dysfunction. Front. Physiol. 2021, 12, 718783. [Google Scholar] [CrossRef]
- Begley, M.; Gahan, C.G.; Hill, C. The interaction between bacteria and bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.D.; Chen, W.D.; Yu, D.; Forman, B.M.; Huang, W. The G-protein-coupled bile acid receptor, Gpbar1 (TGR5), negatively regulates hepatic inflammatory response through antagonizing nuclear factor κ light-chain enhancer of activated B cells (NF-κB) in mice. Hepatology 2011, 54, 1421–1432. [Google Scholar] [CrossRef]
- Takeda, M.; Takei, H.; Suzuki, M.; Tsukui, T.; Tsuboi, K.; Watayo, H.; Ochi, T.; Koga, H.; Nittono, H.; Yamataka, A. Bile acid profiles in adult patients with biliary atresia who achieve native liver survival after portoenterostomy. Sci. Rep. 2024, 14, 2492. [Google Scholar] [CrossRef] [PubMed]
- Hagan, T.; Cortese, M.; Rouphael, N.; Boudreau, C.; Linde, C.; Maddur, M.S.; Das, J.; Wang, H.; Guthmiller, J.; Zheng, N.Y.; et al. Antibiotics-Driven Gut Microbiome Perturbation Alters Immunity to Vaccines in Humans. Cell 2019, 178, 1313–1328. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fei, Y.; Zhou, T.; Ji, H.; Wu, J.; Gu, X.; Luo, Y.; Zhu, J.; Feng, M.; Wan, P.; et al. Bile Acids Impair Vaccine Response in Children with Biliary Atresia. Front. Immunol. 2021, 12, 642546. [Google Scholar] [CrossRef]
- Ponziani, F.R.; Coppola, G.; Rio, P.; Caldarelli, M.; Borriello, R.; Gambassi, G.; Gasbarrini, A.; Cianci, R. Factors Influencing Microbiota in Modulating Vaccine Immune Response: A Long Way to Go. Vaccines 2023, 11, 1609. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Authors | Year | Study Design | Type of Anti-HBV Vaccine | Number of Patients | Outcome |
|---|---|---|---|---|---|
| Gutierrez Domingo et al. [29] | 2012 | Retrospective cohort | Three-dose recombinant vaccine (Engerix B, Recombivax HB) | 278 cirrhotic patients | Higher response in Child-Pugh A (54%) than in B and C (30–33%) |
| Roni et al. [30] | 2013 | Prospective cohort | Three-dose recombinant vaccine (Shanvac-B) | 52 cirrhotic patients | Higher response in Child-Pugh A (88%) than B (33%); alcohol-related disease linked to poorer efficacy |
| Aggeletopoulou et al. [22] | 2017 | Review | Single or double dose of three-dose recombinant vaccine | 961 cirrhotic patients included in 11 prospective and retrospective studies | The mean response rate was 38% for the standard dose and 53% for the double dose |
| Amjad et al. [28] | 2020 | Retrospective cohort | Two-dose (Heplisav-B) and three-dose (Engerix B) recombinant vaccine | 166 CLD patients (34% with cirrhosis) | Higher response rate with two-dose than three-dose regimen (aOR: 2.74, 95% CI 1.31–5.71). |
| Kim et al. [23] | 2023 | Retrospective cohort | Three-dose recombinant vaccine | 126 CLD patients (82% with cirrhosis) | The cirrhotic patient response rate was 51% vs. noncirrhotic, which was 72% (p = 0.04) |
| Authors | Year | Type of Study | Type of Anti-HAV Vaccine | Number of Patients | Outcome |
|---|---|---|---|---|---|
| Keeffe et al. [32] | 1998 | Prospective cohort | Two-dose regimen of the Havrix vaccine | 220 cirrhotic patients (n = 104 with chronic Hepatitis C; n = 46 with chronic Hepatitis B; n = 70 with other CLD) | Adequate seroconversion rate (94–98%) in patients with HBV and HCV-related nonadvanced chronic liver disease |
| Arguedas et al. [33] | 2001 | Prospective cohort | Two-dose regimen of the Havrix vaccine | 84 cirrhotic patients (49 with compensated liver disease and 35 with decompensated disease) | Higher seroconversion rate (98%) in compensated cirrhosis (Child-Pugh A). As the severity of cirrhosis increased, the response rate gradually decreased. |
| Authors | Year | Type of Study | Type of Anti-Pneumococcal Vaccine | Number of Patients | Outcome |
|---|---|---|---|---|---|
| Pirovino et al. [34] | 1984 | Prospective cohort | 14-valent pneumococcal polysaccharide vaccine (Pneumovax-14) | 15 patients with biopsy-proven alcoholic liver cirrhosis (compared to 10 healthy volunteers and 10 patients with chronic obstructive pulmonary disease). | The response rate in patients with alcohol-related cirrhosis was similar to the other groups. |
| Preheim et al. [35] | 1992 | Preclinical study in vivo | Type 3 pneumococcal capsular polysaccharide (PCP) antigen | Rats with induced cirrhosis | Rats with cirrhosis had a substantially higher pneumococcal infection-related mortality than vaccinated healthy rats despite an adequate serological response. |
| McCashland et al. [36] | 2000 | Prospective cohort | Pneumococcal polysaccharide vaccine (PPSV23) | 45 patients with end-stage liver disease (compared to 13 age-matched control subjects) | Specific anti-pneumococcal polysaccharide capsule IgA, IgM, and IgG significantly increased in both patients and healthy controls at one month without statistically significant differences. The comparative 6-month-to-baseline elevations for both IgM and IgA were significantly lower in the patient group than in the control group. |
| Authors | Year | Type of Study | Type of Anti-Influenza- Vaccine | Number of Patients | Outcome |
|---|---|---|---|---|---|
| Gaeta et al. [38] | 2002 | Prospective cohort | 2000/2001 season virosomes adjuvanted influenza vaccine (Inflexal V) | 20 patients with HBV/HCV-related cirrhosis and eight age-matched controls | Seroconversion rate of 75–85% in cirrhotic patients compared to 100% in the control group. |
| Härmälä et al. [37] | 2019 | Meta-analysis comprising 12 studies (1 randomized controlled trial and 11 cohort studies; 6 with clinical outcomes, 6 with serological outcomes) | Monovalent, split virus; trivalent, split virus; trivalent, subunit. | Studies with clinical outcomes: 232 patients with CLD (148 cirrhotic patients), most with viral liver disease. Studies with serological outcomes: 8189 patients with CLD (3258 cirrhotic patients) | A noteworthy seroconversion rate (80% for the A/H1N1 strain and 87% for the B strain). |
| Authors | Year | Type of Study | Type of Anti-COVID-19 Vaccine | Number of Patients | Outcome |
|---|---|---|---|---|---|
| Thuluvath et al. [39] | 2021 | Prospective cohort | mRNA vaccines or Johnson and Johnson vaccine | 233 patients (62 liver transplant recipients, 79 cirrhosis [10 decompensated], 92 CLD without cirrhosis. | Poor antibody responses in 61% of LT recipients and 24% of those with CLD. Only 40% of patients with cirrhosis showed an adequate serological response. |
| Bakasis et al. [40] | 2021 | Prospective cohort | Two doses of mRNA-based vaccinations | 38 patients with cirrhosis and 49 noncirrhotic chronic liver disease compared to 40 controls. | Appropriate rates of seroconversion: 97.4% (37/38) in cirrhotics, 87.8% (43/49) in noncirrhotic liver disease, and 100% (40/40) in controls. |
| Iavarone et al. [41] | 2023 | Prospective cohort | BNT162b2 and mRNA-1273 | 182 cirrhotic patients (85% SARS-CoV-2-naïve) compared to 38 controls. | Anti-spike IgG serum levels were significantly lower in 182 cirrhotic patients who received two doses of mRNA vaccine than in healthy controls (1751 U/mL vs. 4523 U/mL, p = 0.012). |
| Beran et al. [42] | 2023 | Meta-analysis (including four studies) | BNT162b2, mRNA, mRNA-1273, JNJ-784336725, Ad.26.COV2.S, AstraZeneca, Bharat Biotech, CanSino, and Sinovac. | 51834 cirrhotic patients | COVID-19-related hospitalization rate and related mortality rate were significantly lower in vaccinated cirrhotic patients compared to unvaccinated ones. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Airola, C.; Andaloro, S.; Gasbarrini, A.; Ponziani, F.R. Vaccine Responses in Patients with Liver Cirrhosis: From the Immune System to the Gut Microbiota. Vaccines 2024, 12, 349. https://doi.org/10.3390/vaccines12040349
Airola C, Andaloro S, Gasbarrini A, Ponziani FR. Vaccine Responses in Patients with Liver Cirrhosis: From the Immune System to the Gut Microbiota. Vaccines. 2024; 12(4):349. https://doi.org/10.3390/vaccines12040349
Chicago/Turabian StyleAirola, Carlo, Silvia Andaloro, Antonio Gasbarrini, and Francesca Romana Ponziani. 2024. "Vaccine Responses in Patients with Liver Cirrhosis: From the Immune System to the Gut Microbiota" Vaccines 12, no. 4: 349. https://doi.org/10.3390/vaccines12040349