
Intradermal Fractional ChAdOx1 nCoV-19 Booster Vaccine Induces Memory T Cells: A Follow-Up Study

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
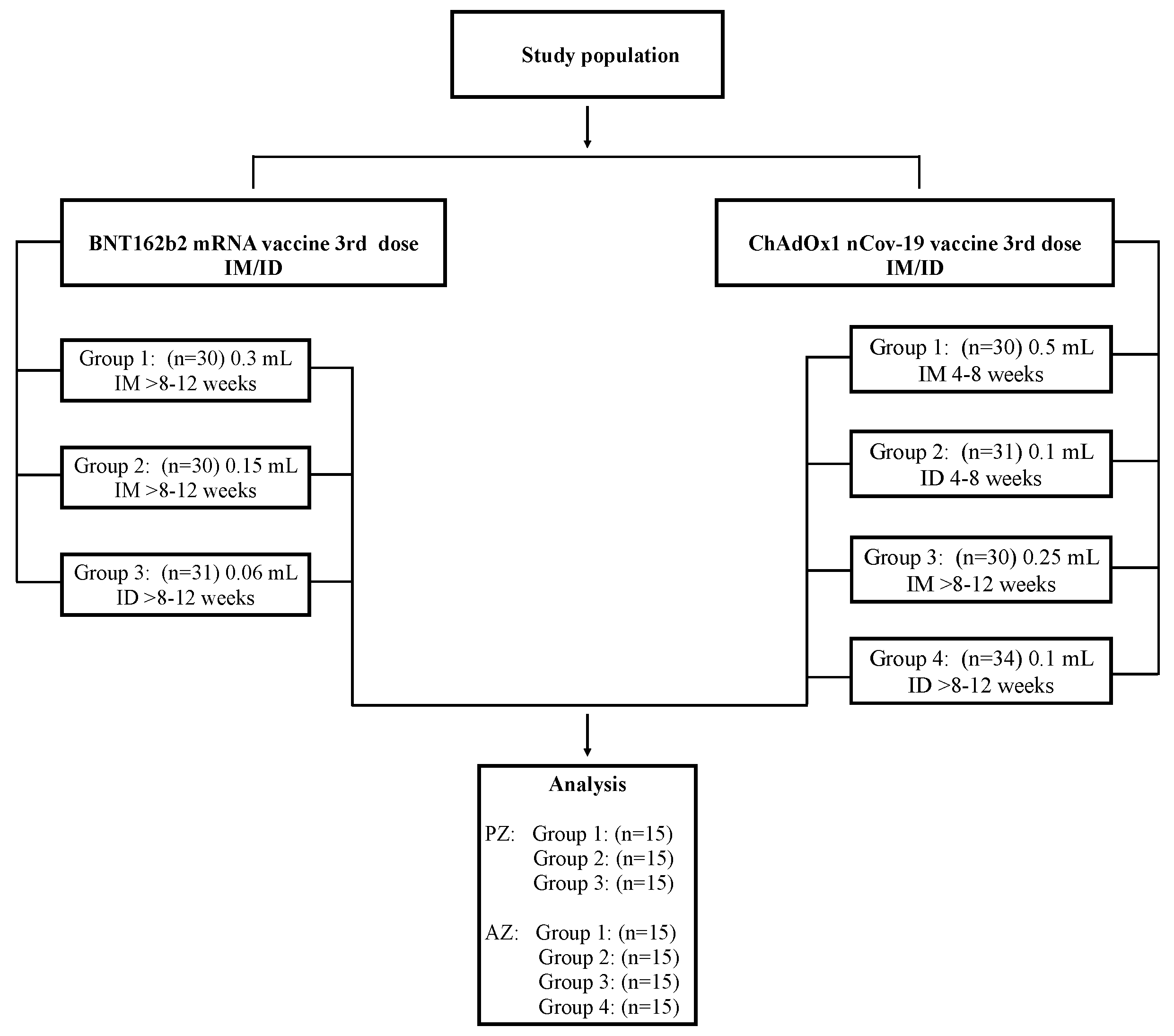
2.1. Study Population
2.2. Separation of Peripheral Blood Mononuclear Cells (PBMCs)
2.3. Flow Cytometry Analysis
2.4. Statistical Analysis
3. Results
3.1. Study Participants
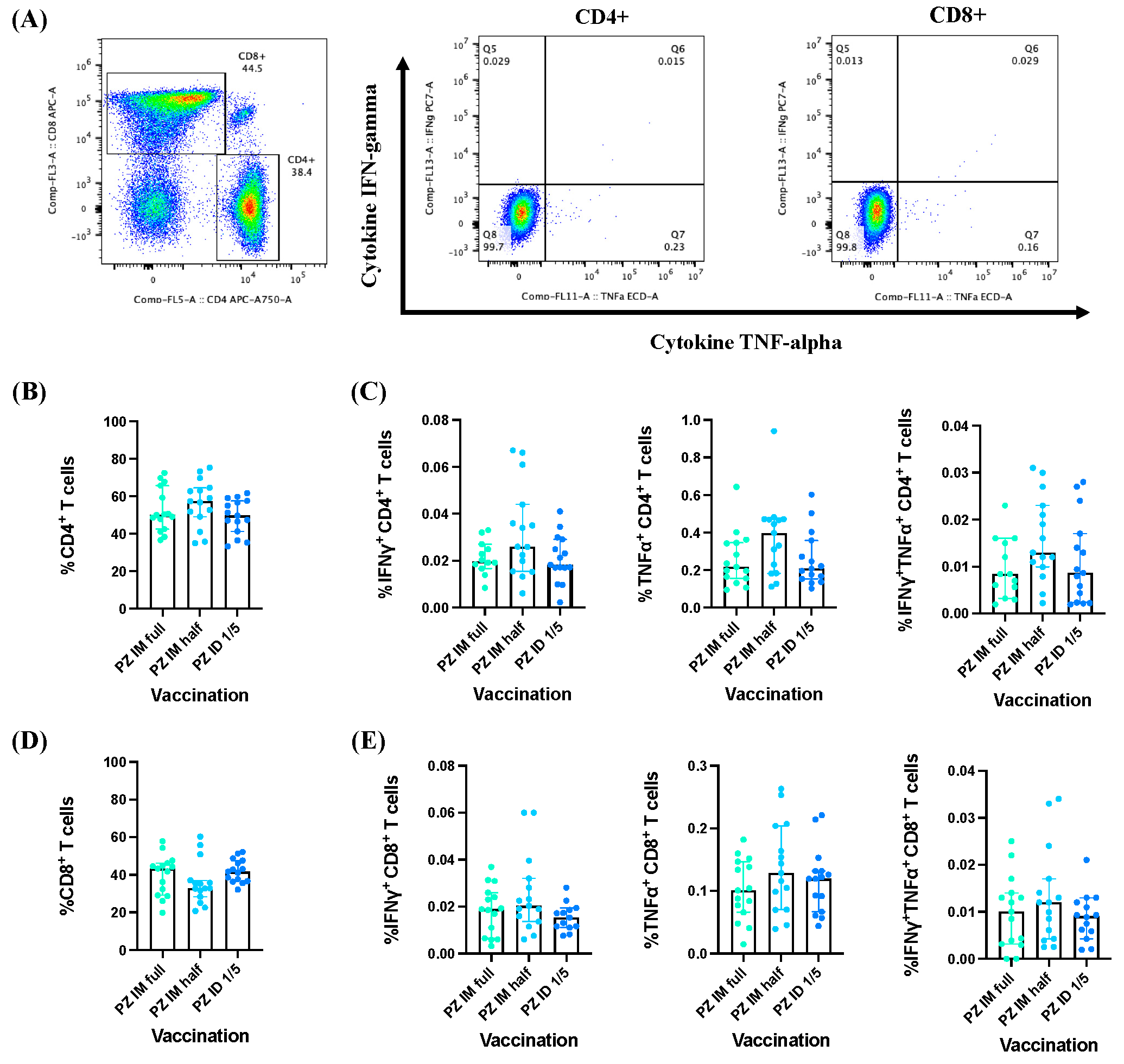
3.2. Effector Cytokine Responses by CD4+/CD8+ T Cells Induced by BNT162b2 mRNA Vaccine Booster Administration following Primary Series of Inactivated SARS-CoV-2 Vaccinations
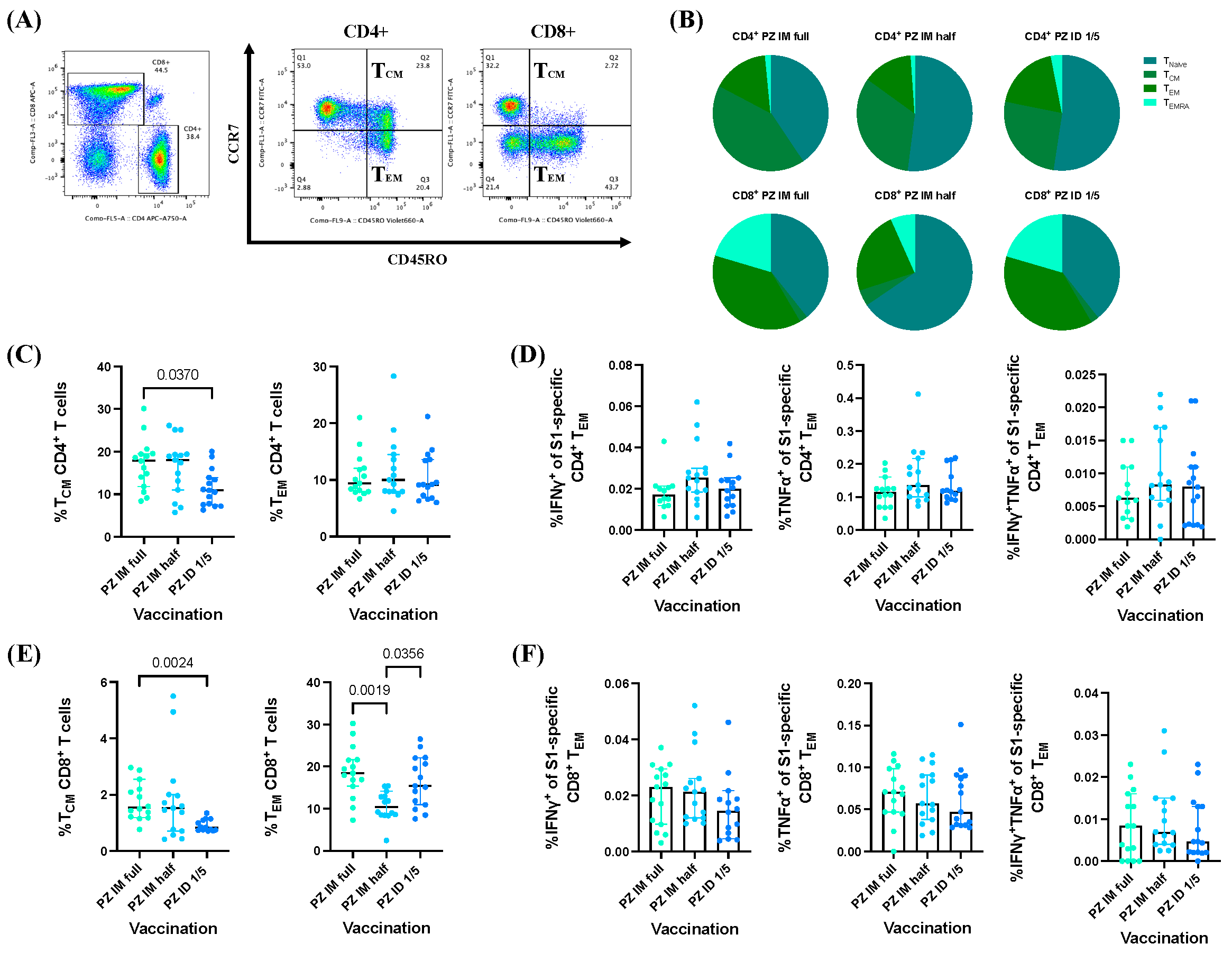
3.3. BNT162b2 mRNA Vaccine Booster Administration following Primary Series of Inactivated SARS-CoV-2 Vaccinations Induced S1-Specific CD4+/CD8+ Effector Memory T Cell (TEM) Responses
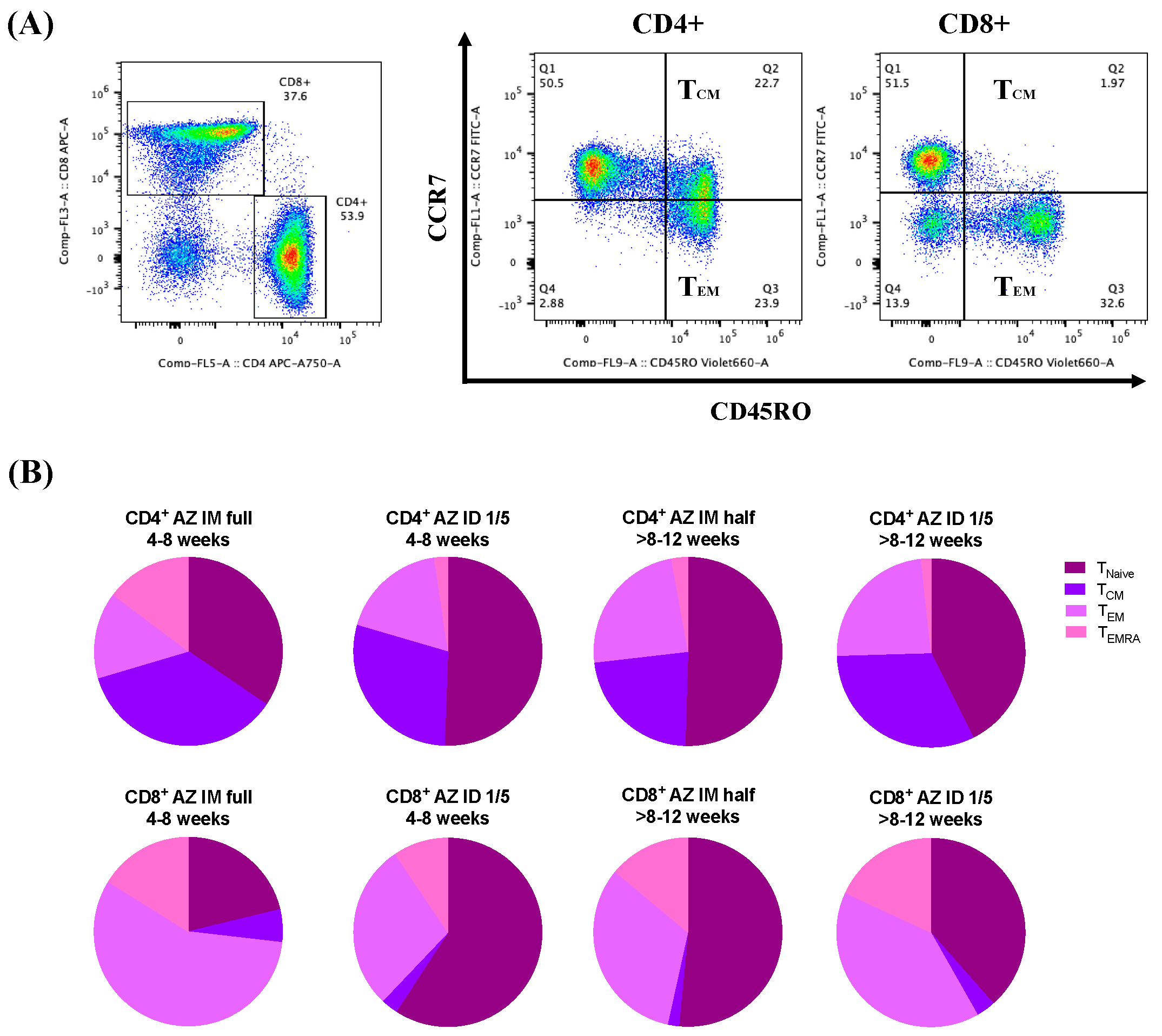
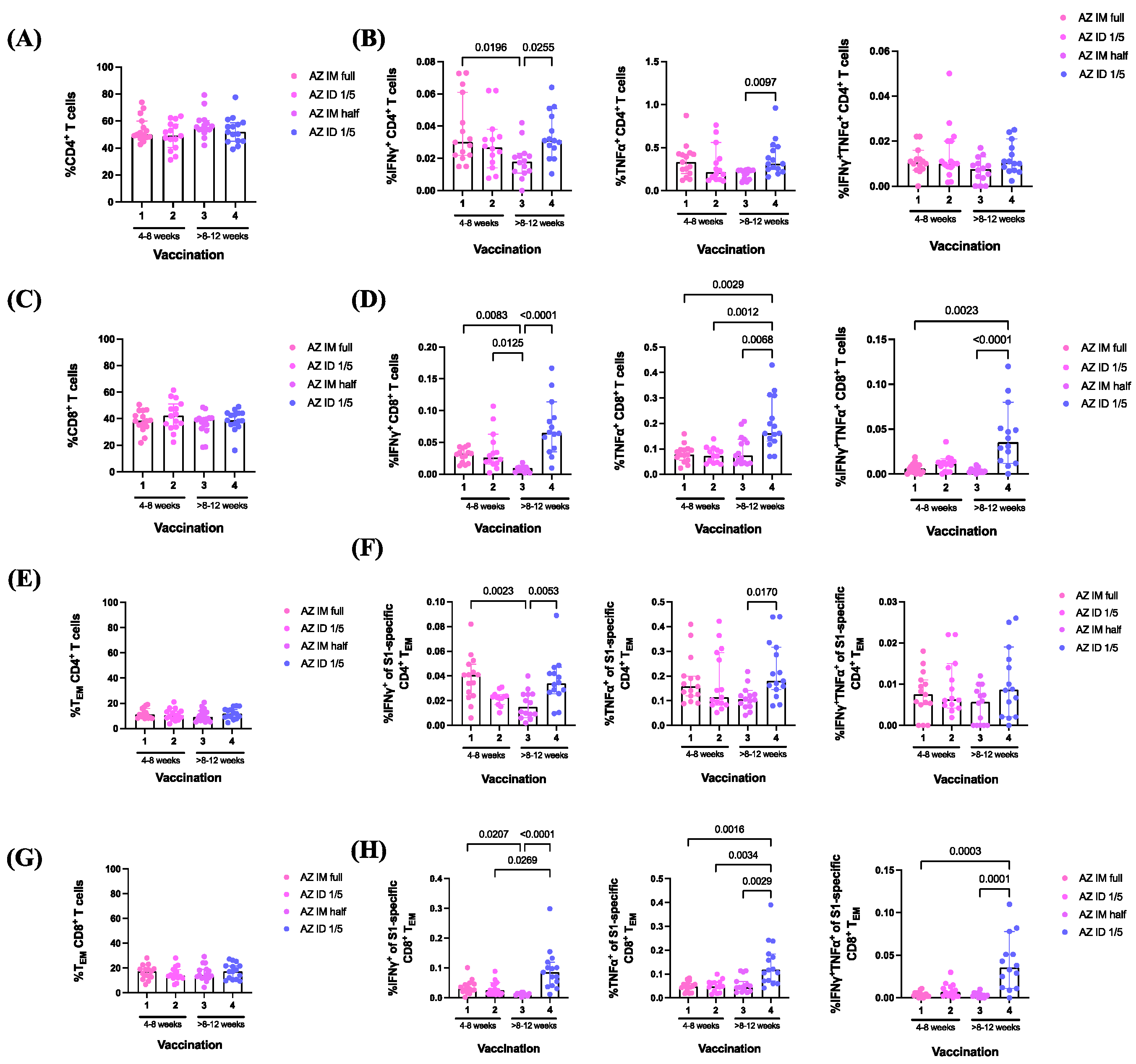
3.4. ChAdOx1 nCoV-19 Vaccine Booster Administration following Primary Series of Inactivated SARS-CoV-2 Vaccinations Induced S1-Specific CD4+/CD8+ Effector Memory T Cell (TEM) Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nohynek, H.; Wilder-Smith, A. Does the world still need new COVID-19 vaccines? N. Engl. J. Med. 2022, 386, 2140–2142. [Google Scholar] [CrossRef]
- Levin, Y.; Balakirski, N.M.; Caraco, Y.; Ben-Ami, E.; Atsmon, J.; Marcus, H. Ethics and execution of developing a 2nd wave COVID vaccine—Our interim phase I/II VSV-SARS-CoV2 vaccine experience. Vaccine 2021, 39, 2821–2823. [Google Scholar] [CrossRef]
- Drozdzik, A.; Drozdzik, M. Oral pathology in COVID-19 and SARS-CoV-2 infection—Molecular aspects. Int. J. Mol. Sci. 2022, 23, 1431. [Google Scholar] [CrossRef]
- Guerrini, G.; Magrì, D.; Gioria, S.; Medaglini, D.; Calzolai, L. Characterization of nanoparticles-based vaccines for COVID-19. Nat. Nanotechnol. 2022, 17, 570–576. [Google Scholar] [CrossRef]
- Agrati, C.; Castilletti, C.; Goletti, D.; Sacchi, A.; Bordoni, V.; Mariotti, D. Persistent spike-specific T cell immunity despite antibody reduction after 3 months from SARS-CoV-2 BNT162b2-mRNA vaccine. Sci. Rep. 2022, 12, 6687. [Google Scholar] [CrossRef]
- Burki, T.K. Omicron variant and booster COVID-19 vaccines. Lancet Respir. Med. 2022, 10, e17. [Google Scholar] [CrossRef]
- Wherry, E.J.; Barouch, D.H. T cell immunity to COVID-19 vaccines. Science 2022, 377, 821–822. [Google Scholar] [CrossRef]
- Kelly, N.E.W.; De Barra, C.; Shaamile, F.; Holland, A.; Shaw, L.; Mallon, P.W.; O’Shea, D. Antigen specific T cells in people with obesity at five months following ChAdOx1 COVID-19 vaccination. Int. J. Obes. 2023, 47, 83–86. [Google Scholar] [CrossRef]
- Mattoo, S.U.S.; Myoung, J. T cell responses to SARS-CoV-2 in humans and animals. J. Microbiol. 2022, 60, 276–289. [Google Scholar] [CrossRef]
- Barouch, D.H. COVID-19 vaccines—Immunity, variants, boosters. N. Engl. J. Med. 2022, 387, 1011–1020. [Google Scholar] [CrossRef]
- Watson, O.J.; Barnsley, G.; Toor, J.; Hogan, A.B.; Winskill, P.; Ghani, A.C. Global impact of the first year of COVID-19 vaccination: A mathematical modelling study. Lancet Infect. Dis. 2022, 22, 1293–1302. [Google Scholar] [CrossRef]
- Geers, D.; Shamier, M.C.; Bogers, S.; den Hartog, G.; Gommers, L.; Nieuwkoop, N.N.; GeurtsvanKessel, C.H. SARS-CoV-2 variants of concern partially escape humoral but not T cell responses in COVID-19 convalescent donors and vaccine recipients. Sci. Immunol. 2021, 6, eabj1750. [Google Scholar] [CrossRef]
- Rabail, R.; Ahmed, W.; Ilyas, M.; Rajoka, M.S.R.; Hassoun, A.; Khalid, A.R.; Aadil, R.M. The side effects and adverse clinical cases reported after COVID-19 immunization. Vaccines 2022, 10, 488. [Google Scholar] [CrossRef]
- Elgendy, M.O.; El-Gendy, A.O.; Mahmoud, S.; Mohammed, T.Y.; Abdelrahim, M.E.; Sayed, A.M. Side effects and efficacy of COVID-19 vaccines among the Egyptian population. Vaccines 2022, 10, 109. [Google Scholar] [CrossRef]
- Lounis, M.; Rais, M.A.; Bencherit, D.; Aouissi, H.A.; Oudjedi, A.; Klugarová, J.; Riad, A. Side effects of COVID-19 inactivated virus vs. adenoviral vector vaccines: Experience of Algerian healthcare workers. Front. Public Health 2022, 10, 896343. [Google Scholar] [CrossRef]
- Omeish, H.; Najadat, A.; Al-Azzam, S.; Tarabin, N.; Abu Hameed, A.; Al-Gallab, N.; Aldeyab, M.A. Reported COVID-19 vaccines side effects among Jordanian population: A cross sectional study. Hum. Vaccines Immunother. 2022, 18, 1981086. [Google Scholar] [CrossRef]
- Roozen, G.V.T.; Anna, H.E.R.; Meta, R. COVID-19 vaccine dose sparing: Strategies to improve vaccine equity and pandemic preparedness. Lancet Glob. Health 2022, 10, e570–e573. [Google Scholar] [CrossRef]
- Migliore, A.; Gigliucci, G.; Di Marzo, R.; Russo, D.; Mammucari, M. Intradermal vaccination: A potential tool in the battle against the COVID-19 pandemic? Risk Manag. Healthc. Policy 2021, 14, 2079–2087. [Google Scholar] [CrossRef]
- Chalermphanchai, N.; Arunothong, W.; Jettavan, N.; Chanta, S.; Boonpisitsakul, T.; Mayoon, M.; Nantawong, T. Safety, tolerability, and antibody response after intradermal vaccination of PFE-BNT in adults who have completed two-doses of Verocell (inactivated vaccine). Vaccine X 2022, 10, 100148. [Google Scholar] [CrossRef]
- Intapiboon, P.; Seepathomnarong, P.; Ongarj, J.; Surasombatpattana, S.; Uppanisakorn, S.; Mahasirimongkol, S.; Pinpathomrat, N. Immunogenicity and safety of an intradermal BNT162b2 mRNA vaccine booster after two doses of inactivated SARS-CoV-2 vaccine in healthy population. Vaccines 2021, 9, 1375. [Google Scholar] [CrossRef]
- Pinpathomrat, N.; Intapiboon, P.; Seepathomnarong, P.; Ongarj, J.; Sophonmanee, R.; Hengprakop, J.; Chusri, S. Immunogenicity and safety of an intradermal ChAdOx1 nCoV-19 boost in a healthy population. NPJ Vaccines 2022, 7, 52. [Google Scholar] [CrossRef]
- Chen, Y.; Yin, S.; Tong, X.; Tao, Y.; Ni, J.; Pan, J.; Shen, H. Dynamic SARS-CoV-2-specific B-cell and T-cell responses following immunization with an inactivated COVID-19 vaccine. Clin. Microbiol. Infect. 2022, 28, 410–418. [Google Scholar] [CrossRef]
- Diniz, M.O.; Mitsi, E.; Swadling, L.; Rylance, J.; Johnson, M.; Goldblatt, D.; Maini, M.K. Airway-resident T cells from unexposed individuals cross-recognize SARS-CoV-2. Nat. Immunol. 2022, 23, 1324–1329. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Immunological memory to SARS-CoV-2 infection and COVID-19 vaccines. Immunol. Rev. 2022, 310, 27–46. [Google Scholar] [CrossRef]
- Niyomnaitham, S.; Chatsiricharoenkul, S.; Toh, Z.Q.; Senawong, S.; Pheerapanyawaranun, C.; Phumiamorn, S.; Chokephaibulkit, K. Evaluation of the safety and immunogenicity of fractional intradermal COVID-19 vaccines as a booster: A pilot study. Vaccines 2022, 10, 1497. [Google Scholar] [CrossRef]
- Schweiger, M. Intradermal COVID-19 vaccination could solve supply problems. Brit. Med. J. 2021, 374, n1980. [Google Scholar] [CrossRef]
- Egunsola, O.; Clement, F.; Taplin, J.; Mastikhina, L.; Li, J.W.; Lorenzetti, D.L.; Noseworthy, T. Intradermal versus intramuscular administration of influenza vaccination: Rapid review and meta-analysis. medRxiv 2020. [Google Scholar] [CrossRef]
- Caldera, F.; Mercer, M.; Samson, S.I.; Pitt, J.M.; Hayney, M.S. Influenza vaccination in immunocompromised populations: Strategies to improve immunogenicity. Vaccine 2021, 39, A15–A23. [Google Scholar] [CrossRef]
- Sophonmanee, R.; Ongarj, J.; Seeyankem, B.; Seepathomnarong, P.; Intapiboon, P.; Surasombatpattana, S.; Pinpathomrat, N. T-cell responses induced by an intradermal BNT162b2 mRNA vaccine booster following primary vaccination with inactivated SARS-CoV-2 vaccine. Vaccines 2022, 10, 1494. [Google Scholar] [CrossRef]
- Moss, P. The T cell immune response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef]
- Nelde, A.; Bilich, T.; Heitmann, J.S.; Maringer, Y.; Salih, H.R.; Roerden, M.; Walz, J.S. SARS-CoV-2-derived peptides define heterologous and COVID-19-induced T cell recognition. Nat. Immunol. 2021, 22, 74–85. [Google Scholar] [CrossRef]
- van Zelm, M.C. Immune memory to SARS-CoV-2 Omicron BA.1 breakthrough infections: To change the vaccine or not? Sci. Immunol. 2022, 7, eabq5901. [Google Scholar] [CrossRef]
- Cohen, K.W.; Linderman, S.L.; Moodie, Z.; Czartoski, J.; Lai, L.; Mantus, G.; McElrath, M.J. Longitudinal analysis shows durable and broad immune memory after SARS-CoV-2 infection with persisting antibody responses and memory B and T cells. Cell. Rep. Med. 2021, 2, 00354. [Google Scholar] [CrossRef]
- Zhou, X.; Jiang, X.; Qu, M.; Aninwene, G.E.; Jucaud, V.; Moon, J.J.; Khademhosseini, A. Engineering antiviral vaccines ACS nano. ACS Nano 2020, 1, 12370–12389. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Wong, L.A.; Yap, C.G.; Jahan, N.K.; Pillai, N. COVID-19 vaccine: Review of the mechanism of action of different types of vaccine. Open Access Libr. J. 2022, 9, 1–20. [Google Scholar] [CrossRef]
- Cox, R.J.; Brokstad, K.A. Not just antibodies: B cells and T cells mediate immunity to COVID-19. Nat. Rev. Immunol. 2020, 20, 581–582. [Google Scholar] [CrossRef]
- Mascellino, M.T.; Di Timoteo, F.; De Angelis, M.; Oliva, A. Overview of the main anti-SARS-CoV-2 vaccines: Mechanism of action, efficacy, and safety. Infect. Drug Resist. 2021, 14, 3459–3476. [Google Scholar] [CrossRef]
- DiPiazza, A.T.; Graham, B.S.; Ruckwardt, T.J. T cell immunity to SARS-CoV-2 following natural infection and vaccination. Biochem. Biophys. Res. Commun. 2021, 538, 211–217. [Google Scholar] [CrossRef]
- Jarjour, N.N.; Masopust, D.; Jameson, S.C. T cell memory: Understanding COVID-19. Immunity 2021, 54, 14–18. [Google Scholar] [CrossRef]
- Pober, J.S.; Kluger, M.S.; Schechner, J.S. Human endothelial cell presentation of antigen and the homing of memory/effector T cells to skin. Ann. N. Y. Acad. Sci. 2001, 94, 12–25. [Google Scholar] [CrossRef]
- Mathers, A.R.; Larregina, A.T. Professional antigen-presenting cells of the skin. Immunol. Res. 2006, 36, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Lutz, M.B.; Kurts, C. Induction of peripheral CD4+ T-cell tolerance and CD8+ T-cell cross-tolerance by dendritic cells. Eur. J. Immunol. 2009, 39, 2325–2330. [Google Scholar] [CrossRef] [PubMed]
- Cinicola, B.L.; Piano Mortari, E.; Zicari, A.M.; Agrati, C.; Bordoni, V.; Albano, C.; Carsetti, R. The BNT162b2 vaccine induces humoral and cellular immune memory to SARS-CoV-2 Wuhan strain and the Omicron variant in children 5 to 11 years of age. Front. Immunol. 2022, 13, 1094727. [Google Scholar] [CrossRef] [PubMed]
- Gyssens, I.; Netea, M. Heterologous effects of vaccination and trained immunity. Clin. Microbiol. Infect. 2019, 25, 1457–1458. [Google Scholar] [CrossRef]
- Kent, S.J.; Khoury, D.S.; Reynaldi, A.; Juno, J.A.; Wheatley, A.K.; Stadler, E.; Davenport, M.P. Disentangling the relative importance of T cell responses in COVID-19: Leading actors or supporting cast? Nat. Rev. Immunol. 2022, 22, 387–397. [Google Scholar] [CrossRef]
- Bilich, T.; Nelde, A.; Heitmann, J.S.; Maringer, Y.; Roerden, M.; Bauer, J.; Walz, J.S. T cell and antibody kinetics delineate SARS-CoV-2 peptides mediating long-term immune responses in COVID-19 convalescent individuals. Sci. Transl. Med. 2021, 13, eabf7517. [Google Scholar] [CrossRef]
- Tarke, A.; Sidney, J.; Methot, N.; Zhang, Y.; Dan, J.M.; Goodwin, B.; Sette, A. Negligible impact of SARS-CoV-2 variants on CD4+ and CD8+ T cell reactivity in COVID-19 exposed donors and vaccinees. bioRxiv 2021. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Baseline Characteristics | Total n = 105 (%) | G1 PZ IM Full (>8–12 wk.) n = 15 (%) | G2 PZ IM Half (>8–12 wk.) n = 15 (%) | G3 PZ ID 1/5 (>8–12 wk.) n = 15 (%) | G1 AZ IM Full (4–8 wk.) n = 15 (%) | G2 AZ ID 1/5 (4–8 wk.) n = 15 (%) | G3 AZ IM Half (>8–12 wk.) n = 15 (%) | G4 AZ ID 1/5 (>8–12 wk.) n = 15 (%) |
|---|---|---|---|---|---|---|---|---|
| Female | 68 (64.8%) | 11 (73.3) | 6 (40) | 7 (46.7) | 15 (100%) | 11 (73.3) | 8 (53.3) | 10 (66.7) |
| Mean age, years (IQR) | 39.8 (9.05) | 41.9 (7.6) | 41.7 (8.1) | 37.4 (9.2) | 33.7 (7.6) | 36.1 (8.9) | 47.6 (6.4) | 40.6 (9.4) |
| Median duration between primary vaccine series and booster dose, days (IQR) | 69.0 (36.0, 94.0) | 73 (72, 73) | 73 (68.5, 74) | 74 (70.5, 74) | 45 (44, 46) | 51 (45, 52) | 68 (67, 73) | 80 (76.5, 82) |
| Median follow-up duration after boosting, days (IQR) | 91.0 (88.0, 111) | 91 (90.5, 91) | 91 (90, 91) | 91 (90, 91) | 93 (93, 93) | 93 (91, 93) | 91 (91, 91) | 91 (91, 93) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sophonmanee, R.; Preampruchcha, P.; Ongarj, J.; Seeyankem, B.; Intapiboon, P.; Surasombatpattana, S.; Uppanisakorn, S.; Sangsupawanich, P.; Chusri, S.; Pinpathomrat, N. Intradermal Fractional ChAdOx1 nCoV-19 Booster Vaccine Induces Memory T Cells: A Follow-Up Study. Vaccines 2024, 12, 109. https://doi.org/10.3390/vaccines12020109
Sophonmanee R, Preampruchcha P, Ongarj J, Seeyankem B, Intapiboon P, Surasombatpattana S, Uppanisakorn S, Sangsupawanich P, Chusri S, Pinpathomrat N. Intradermal Fractional ChAdOx1 nCoV-19 Booster Vaccine Induces Memory T Cells: A Follow-Up Study. Vaccines. 2024; 12(2):109. https://doi.org/10.3390/vaccines12020109
Chicago/Turabian StyleSophonmanee, Ratchanon, Perawas Preampruchcha, Jomkwan Ongarj, Bunya Seeyankem, Porntip Intapiboon, Smonrapat Surasombatpattana, Supattra Uppanisakorn, Pasuree Sangsupawanich, Sarunyou Chusri, and Nawamin Pinpathomrat. 2024. "Intradermal Fractional ChAdOx1 nCoV-19 Booster Vaccine Induces Memory T Cells: A Follow-Up Study" Vaccines 12, no. 2: 109. https://doi.org/10.3390/vaccines12020109