
Neutralizing Activity against BQ.1.1, BN.1, and XBB.1 in Bivalent COVID-19 Vaccine Recipients: Comparison by the Types of Prior Infection and Vaccine Formulations
 , , ,
, , ,
Abstract
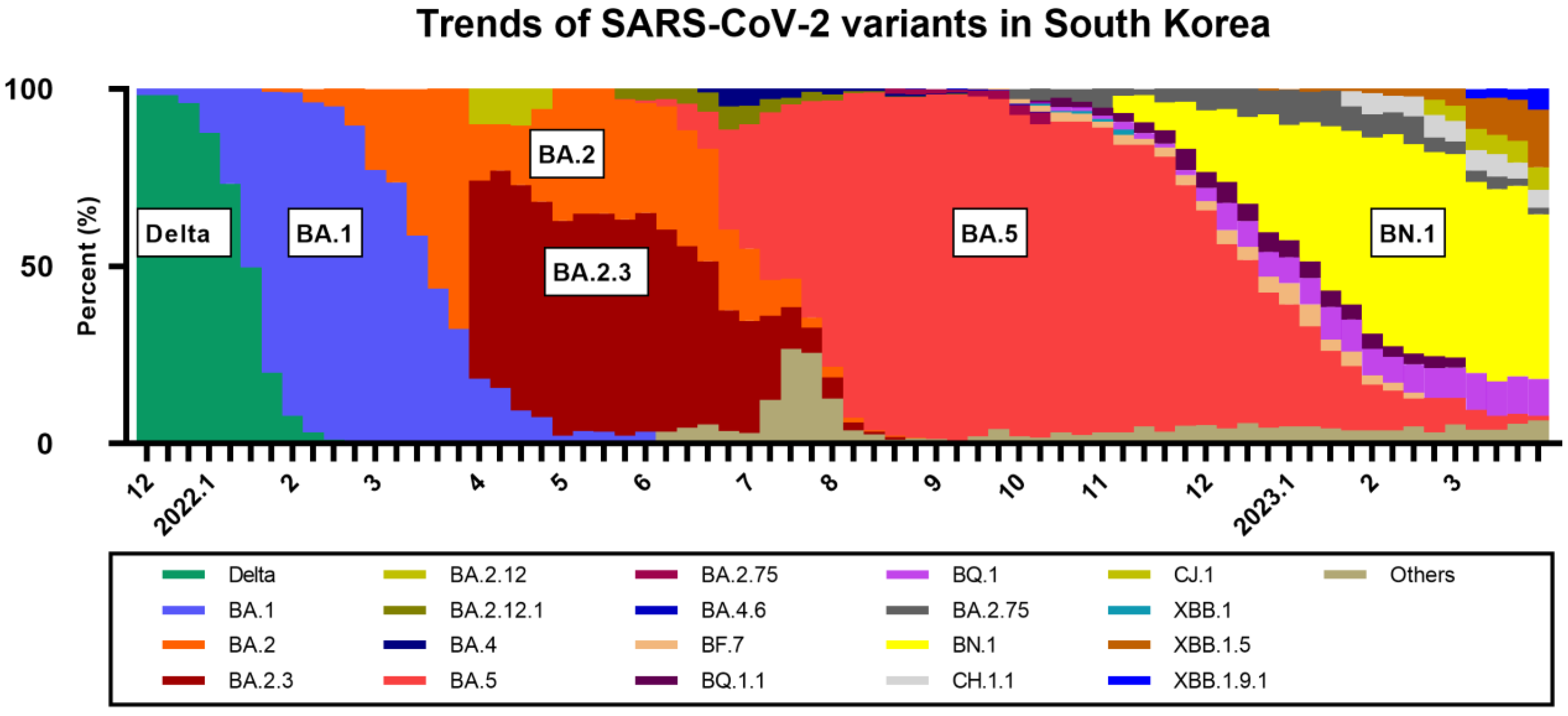
:1. Introduction
2. Materials and Methods
2.1. Study Design and Procedure
2.2. Immunological Analysis
2.3. Statistical Analysis
2.4. Ethics Statement
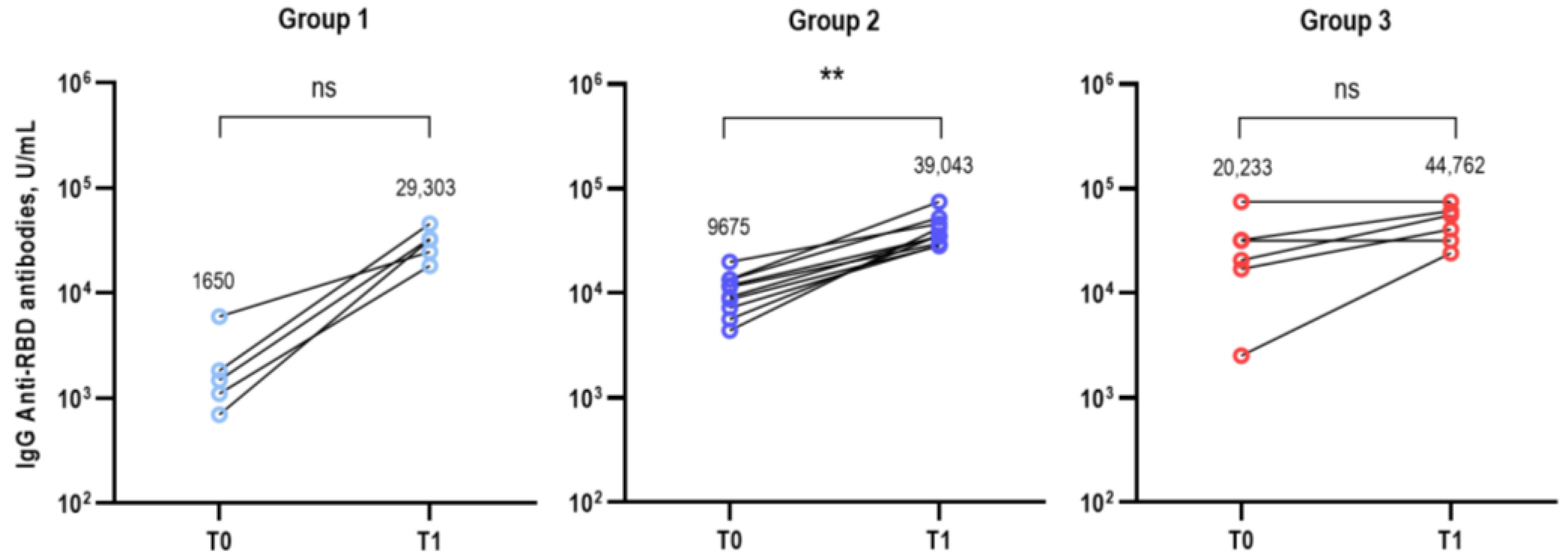
3. Results
3.1. Study Participants
3.2. Immunological Analysis after Bivalent COVID-19 Vaccination
3.3. Reactogenicity of the Bivalent COVID-19 Vaccine
4. Discussions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Classification of Omicron (B.1.1.529): SARS-CoV-2 Variant of Concern. 2021. Available online: https://www.who.int/news/item/26-11-2021-classification-of-omicron-(b.1.1.529)-sars-cov-2-variant-of-concern (accessed on 30 June 2023).
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/region/wpro/country/kr (accessed on 30 June 2023).
- Wu, N.; Joyal-Desmarais, K.; Ribeiro, P.A.B.; Vieira, A.M.; Stojanovic, J.; Sanuade, C.; Yip, D.; Bacon, S.L. Long-term effectiveness of COVID-19 vaccines against infections, hospitalisations, and mortality in adults: Findings from a rapid living systematic evidence synthesis and meta-analysis up to December, 2022. Lancet Respir. Med. 2023, 11, 439–452. [Google Scholar] [CrossRef]
- Kim, Y.Y.; Choe, Y.J.; Kim, J.; Kim, R.K.; Jang, E.J.; Lee, H.; Yi, S.; Lee, S.; Park, Y.J. Vaccine Effectiveness Against Severe Disease and Death for Patients With COVID-19 During the Delta-Dominant and Omicron-Emerging Periods: A K-COVE Study. J. Korean Med. Sci. 2023, 38, e87. [Google Scholar] [CrossRef]
- Bobrovitz, N.; Ware, H.; Ma, X.; Li, Z.; Hosseini, R.; Cao, C.; Selemon, A.; Whelan, M.; Premji, Z.; Issa, H.; et al. Protective effectiveness of previous SARS-CoV-2 infection and hybrid immunity against the omicron variant and severe disease: A systematic review and meta-regression. Lancet Infect. Dis. 2023, 23, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Iketani, S.; Li, Z.; Liu, L.; Guo, Y.; Huang, Y.; Bowen, A.D.; Liu, M.; Wang, M.; Yu, J.; et al. Alarming antibody evasion properties of rising SARS-CoV-2 BQ and XBB subvariants. Cell 2023, 186, 279–286.e278. [Google Scholar] [CrossRef] [PubMed]
- Hyun, H.; Jang, A.Y.; Park, H.; Heo, J.Y.; Seo, Y.B.; Nham, E.; Yoon, J.G.; Seong, H.; Noh, J.Y.; Cheong, H.J.; et al. Humoral and cellular immunogenicity of homologous and heterologous booster vaccination in Ad26.COV2.S-primed individuals: Comparison by breakthrough infection. Front. Immunol. 2023, 14, 1131229. [Google Scholar] [CrossRef]
- Zou, J.; Kurhade, C.; Patel, S.; Kitchin, N.; Tompkins, K.; Cutler, M.; Cooper, D.; Yang, Q.; Cai, H. Neutralization of BA.4–BA.5, BA.4.6, BA.2.75.2, BQ.1.1, and XBB.1 with Bivalent Vaccine. N. Engl. J. Med. 2023, 388, 853–854. [Google Scholar] [CrossRef]
- Miller, J.; Hachmann, N.P.; Collier, A.r.Y.; Lasrado, N.; Mazurek, C.R.; Patio, R.C.; Powers, O.; Surve, N.; Theiler, J.; Korber, B.; et al. Substantial Neutralization Escape by SARSCoV-2 Omicron Variants BQ.1.1 and XBB.1. N. Engl. J. Med. 2023, 388, 660–662. [Google Scholar] [CrossRef]
- Wang, Q.; Bowen, A.; Tam, A.R.; Valdez, R.; Stoneman, E.; Mellis, I.A.; Gordon, A.; Liu, L.; Ho, D.D. SARS-CoV-2 neutralising antibodies after bivalent versus monovalent booster. Lancet Infect. Dis. 2023, 23, 527–528. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. Coronavirus (COVID-19) Update: FDA Authorizes Moderna, Pfizer-BioNTech Bivalent COVID-19 Vaccines for Use as a Booster Dose. 2022. Available online: https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-authorizes-moderna-pfizer-biontech-bivalent-covid-19-vaccines-use (accessed on 12 April 2023).
- Korea Disease Control and Prevention Agency. Regular Briefing: Transition to Once-a-Year COVID-19 Vaccination (March 22). 2023. Available online: https://www.kdca.go.kr/board/board.es?mid=a20501010000&bid=0015&list_no=722117&cg_code=&act=view&nPage=1# (accessed on 27 July 2023).
- Nextstrain Team. Genomic Epidemiology of SARS-CoV-2 with Subsampling Focused Globally over the Past 6 Months. Available online: https://nextstrain.org/ncov/gisaid/global/6m (accessed on 27 July 2023).
- Korea Disease Control and Prevention Agency. Current Status of SARS-CoV-2 Variants in South Korea. Available online: https://www.kdca.go.kr/contents.es?mid=a20107030000 (accessed on 12 April 2023).
- Hodcroft, E.B. CoVariants: SARS-CoV-2 Mutations and Variants of Interest. Available online: https://covariants.org (accessed on 12 April 2023).
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.; Phillips, D.J.; White, T.; Sayal, H.; Aley, P.K.; Bibi, S.; Dold, C.; Fuskova, M.; Gilbert, S.C.; Hirsch, I.; et al. Correlates of protection against symptomatic and asymptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 2032–2040. [Google Scholar] [CrossRef]
- Uraki, R.; Ito, M.; Kiso, M.; Yamayoshi, S.; Iwatsuki-Horimoto, K.; Sakai-Tagawa, Y.; Furusawa, Y.; Imai, M.; Koga, M.; Yamamoto, S.; et al. Efficacy of antivirals and bivalent mRNA vaccines against SARS-CoV-2 isolate CH.1.1. Lancet Infect. Dis. 2023, 23, 525–526. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, A.K.; Fox, A.; Tan, H.X.; Juno, J.A.; Davenport, M.P.; Subbarao, K.; Kent, S.J. Immune imprinting and SARS-CoV-2 vaccine design. Trends Immunol. 2021, 42, 956–959. [Google Scholar] [CrossRef] [PubMed]
- Arbel, R.; Peretz, A.; Sergienko, R.; Friger, M.; Beckenstein, T.; Duskin-Bitan, H.; Yaron, S.; Hammerman, A.; Bilenko, N.; Netzer, D. Effectiveness of a bivalent mRNA vaccine booster dose to prevent severe COVID-19 outcomes: A retrospective cohort study. Lancet Infect. Dis. 2023, 23, 914–921. [Google Scholar] [CrossRef] [PubMed]
- Lasrado, N.; Collier, A.-r.Y.; Miller, J.; Hachmann, N.P.; Liu, J.; Sciacca, M.; Wu, C.; Anand, T.; Bondzie, E.A.; Fisher, J.L.; et al. Waning Immunity Against XBB.1.5 Following Bivalent mRNA Boosters. bioRxiv 2023. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Chakraborti, S.; Gill, J.; Goswami, R.; Kumar, S.; Chandele, A.; Sharma, A. Structural Profiles of SARS-CoV-2 Variants in India. Curr. Microbiol. 2022, 80, 1. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Jian, F.; Wang, J.; Yu, Y.; Song, W.; Yisimayi, A.; Wang, J.; An, R.; Chen, X.; Zhang, N.; et al. Imprinted SARS-CoV-2 humoral immunity induces convergent Omicron RBD evolution. Nature 2023, 614, 521–529. [Google Scholar] [CrossRef]
- Kurhade, C.; Zou, J.; Xia, H.; Liu, M.; Chang, H.C.; Ren, P.; Xie, X.; Shi, P.Y. Low neutralization of SARS-CoV-2 Omicron BA.2.75.2, BQ.1.1 and XBB.1 by parental mRNA vaccine or a BA.5 bivalent booster. Nat. Med. 2023, 29, 344–347. [Google Scholar] [CrossRef]
- Yue, C.; Song, W.; Wang, L.; Jian, F.; Chen, X.; Gao, F.; Shen, Z.; Wang, Y.; Wang, X.; Cao, Y. ACE2 binding and antibody evasion in enhanced transmissibility of XBB.1.5. Lancet Infect. Dis. 2023, 23, 278–280. [Google Scholar] [CrossRef]
- Uriu, K.; Ito, J.; Zahradnik, J.; Fujita, S.; Kosugi, Y.; Schreiber, G.; Genotype to Phenotype Japan, C.; Sato, K. Enhanced transmissibility, infectivity, and immune resistance of the SARS-CoV-2 omicron XBB.1.5 variant. Lancet Infect. Dis. 2023, 23, 280–281. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Statement on the Antigen Composition of COVID-19 Vaccines. Available online: https://www.who.int/news/item/18-05-2023-statement-on-the-antigen-composition-of-covid-19-vaccines (accessed on 30 June 2023).
- Yamasoba, D.; Uriu, K.; Plianchaisuk, A.; Kosugi, Y.; Pan, L.; Zahradnik, J.; Ito, J.; Sato, K. Virological characteristics of the SARS-CoV-2 omicron XBB.1.16 variant. Lancet Infect. Dis. 2023, 23, 655–656. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1 | Group 2 | Group 3 | |
|---|---|---|---|
| SARS-CoV-2 Infection-Naïve (n = 5) | Prior BA.1/BA.2-Infected (n = 10) | Prior BA.5-Infected (n = 6) | |
| Demographics | |||
| Male, n (%) | 2 (40.0) | 2 (20.0) | 3 (50.0) |
| Median age (IQR), y | 65 (61–71) | 63 (62–67) | 64 (50–67) |
| Median BMI (IQR), kg/m2 | 21.8 (21.2–23.7) | 25.4 (21.3–26.0) | 24.8 (24.4–30.5) |
| Medical comorbidities, n (%) | 2 (40.0) | 5 (50.0) | 2 (33.3) |
| Chronic heart disease | 0 (0) | 1 (10.0) | 1 (16.7) |
| Chronic lung disease | 0 (0) | 0 (0) | 0 (0) |
| Chronic liver disease | 0 (0) | 0 (0) | 0 (0) |
| Chronic renal disease | 0 (0) | 0 (0) | 0 (0) |
| Rheumatologic disease | 0 (0) | 1 (10.0) | 0 (0) |
| Hypertension | 1 (20.0) | 1 (10.0) | 1 (16.7) |
| Diabetes mellitus | 2 (40.0) | 2 (20.0) | 2 (33.3) |
| Dyslipidemia | 0 (0) | 0 (0) | 0 (0) |
| Previous vaccination, n (%) | |||
| ChAd/ChAd/BNT/NVX | 3 (60.0) | 7 (70.0) | 3 (50.0) |
| ChAd/ChAd/M/NVX | 1 (20.0) | 2 (20.0) | 2 (33.3) |
| BNT/BNT/BNT/NVX | 1 (20.0) | 0 (0) | 0 (0) |
| BNT/BNT/NVX | 0 (0) | 0 (0) | 1 (16.7) |
| M/M/NVX | 0 (0) | 1 (10.0) | 0 (0) |
| Median interval between previous dose and bivalent vaccination (IQR), days | 190 (188–219) | 216 (189–237) | 230 (221–272) |
| Bivalent vaccine, n (%) | |||
| BA.4/5 | 2 (40.0) | 4 (40.0) | 6 (100) |
| BA.1 | 3 (60.0) | 6 (60.0) | 0 (0) |
| Group 1 | Group 2 | Group 3 | p-Value | |||
|---|---|---|---|---|---|---|
| SARS-CoV-2 Infection-Naïve (n = 5) | Prior BA.1/BA.2-Infected (n = 10) | Prior BA.5-Infected (n = 6) | Group 1 vs. Group 2 | Group 2 vs. Group 3 | Group 3 vs. Group 1 | |
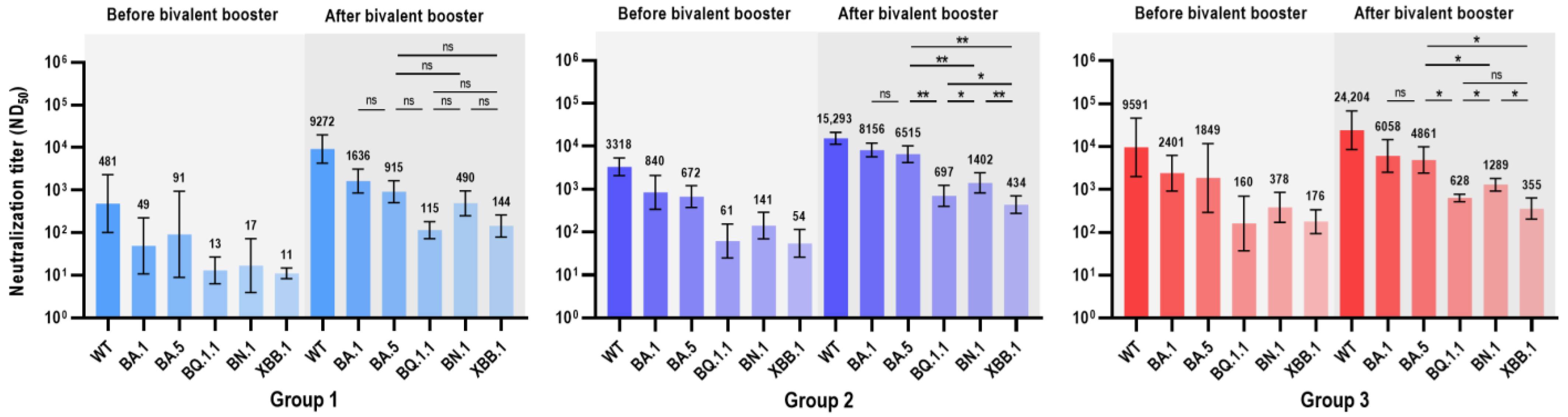
| Before Bivalent COVID-19 Vaccine, GMT (95% CI) | ||||||
| Wild type | 481 (100–2300) | 3318 (2061–5342) | 9591 (1987–46,290) | 0.028 | 0.056 | 0.009 |
| BA.1 | 49 (11–224) | 840 (339–2084) | 2401 (920–6264) | 0.003 | 0.093 | 0.004 |
| BA.5 | 91 (9–939) | 672 (376–1200) | 1849 (290–11,799) | 0.055 | 0.042 | 0.017 |
| BQ.1.1 | 13 (6–27) | 61 (25–152) | 160 (37–693) | 0.024 | 0.072 | 0.022 |
| BN.1 | 17 (4–73) | 141 (69–290) | 378 (169–847) | 0.007 | 0.056 | 0.009 |
| XBB.1 | 11 (8–15) | 55 (26–115) | 176 (93–334) | 0.016 | 0.007 | 0.004 |
| After Bivalent COVID-19 Vaccine, GMT (95% CI) | ||||||
| Wild type | 9272 (4319–19,905) | 15,293 (11,082–21,104) | 24,204 (8558–68,450) | 0.099 | 0.492 | 0.178 |
| BA.1 | 1636 (850–3150) | 8156 (5627–11,823) | 6058 (2520–14,561) | 0.001 | 0.264 | 0.009 |
| BA.5 | 915 (506–1655) | 6515 (4154–10,217) | 4861 (2393–9875) | 0.001 | 0.492 | 0.004 |
| BQ.1.1 | 115 (73–183) | 697 (397–1222) | 628 (512–770) | 0.001 | 0.713 | 0.004 |
| BN.1 | 490 (251–958) | 1402 (818–2404) | 1289 (921–1804) | 0.028 | 0.492 | 0.004 |
| XBB.1 | 144 (79–261) | 434 (271–695) | 355 (201–626) | 0.005 | 0.562 | 0.009 |
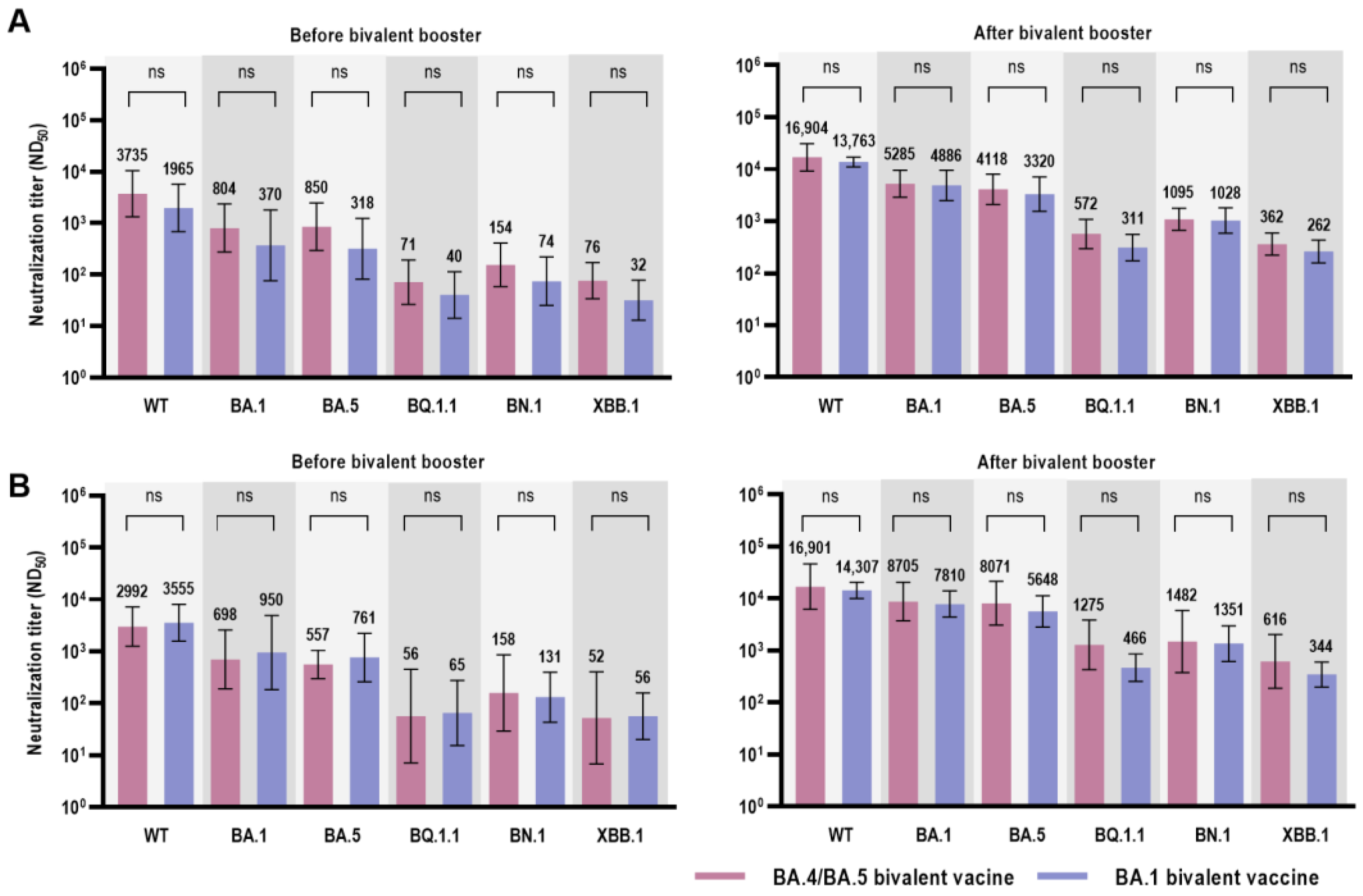
| BNT162b2 (BA.4/BA.5 Bivalent) | mRNA-1273.214 (BA.1 Bivalent) | p-Value | |
|---|---|---|---|
| Before Bivalent COVID-19 Vaccine, GMT (95% CI) | |||
| Wild type | 2992 (1252–7148) | 3555 (1573–8035) | 0.914 |
| BA.1 | 698 (189–2579) | 950 (184–4916) | 0.914 |
| BA.5 | 557 (298–1042) | 761 (260–2222) | 0.762 |
| BQ.1.1 | 57 (7–450) | 65 (15–274) | 0.657 |
| BN.1 | 158 (29–855) | 131 (43–398) | 0.762 |
| XBB.1 | 52 (7–400) | 56 (20–158) | 0.814 |
| After Bivalent COVID-19 Vaccine, GMT (95% CI) | |||
| Wild type | 16,901 (6204–46,047) | 14,307 (10,047–20,372) | 0.762 |
| BA.1 | 8705 (3722–20,358) | 7810 (4362–13,984) | 0.914 |
| BA.5 | 8071 (3060–21,287) | 5648 (2842–11,224) | 0.476 |
| BQ.1.1 | 1275 (424–3837) | 466 (254–855) | 0.067 |
| BN.1 | 1482 (373–5894) | 1351 (613–2977) | 0.762 |
| XBB.1 | 616 (187–2023) | 344 (198–599) | 0.352 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hyun, H.-J.; Choi, M.-J.; Nham, E.; Seong, H.; Yoon, J.-G.; Noh, J.-Y.; Cheong, H.-J.; Kim, W.-J.; Yoon, S.-K.; Park, S.-J.; et al. Neutralizing Activity against BQ.1.1, BN.1, and XBB.1 in Bivalent COVID-19 Vaccine Recipients: Comparison by the Types of Prior Infection and Vaccine Formulations. Vaccines 2023, 11, 1320. https://doi.org/10.3390/vaccines11081320
Hyun H-J, Choi M-J, Nham E, Seong H, Yoon J-G, Noh J-Y, Cheong H-J, Kim W-J, Yoon S-K, Park S-J, et al. Neutralizing Activity against BQ.1.1, BN.1, and XBB.1 in Bivalent COVID-19 Vaccine Recipients: Comparison by the Types of Prior Infection and Vaccine Formulations. Vaccines. 2023; 11(8):1320. https://doi.org/10.3390/vaccines11081320
Chicago/Turabian StyleHyun, Hak-Jun, Min-Joo Choi, Eliel Nham, Hye Seong, Jin-Gu Yoon, Ji-Yun Noh, Hee-Jin Cheong, Woo-Joo Kim, Sun-Kyung Yoon, Se-Jin Park, and et al. 2023. "Neutralizing Activity against BQ.1.1, BN.1, and XBB.1 in Bivalent COVID-19 Vaccine Recipients: Comparison by the Types of Prior Infection and Vaccine Formulations" Vaccines 11, no. 8: 1320. https://doi.org/10.3390/vaccines11081320