
Sex Differences and Cytokine Profiles among Patients Hospitalized for COVID-19 and during Their Recovery: The Predominance of Adhesion Molecules in Females and Oxidative Stress in Males
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. ELISA Assay
2.3. NETs Quantity from the Plasma of the Patients with COVID-19
2.4. Nitro Blue Tetrazolium (NBT) Test of ROS Activity
2.5. Statistical Analysis
3. Results
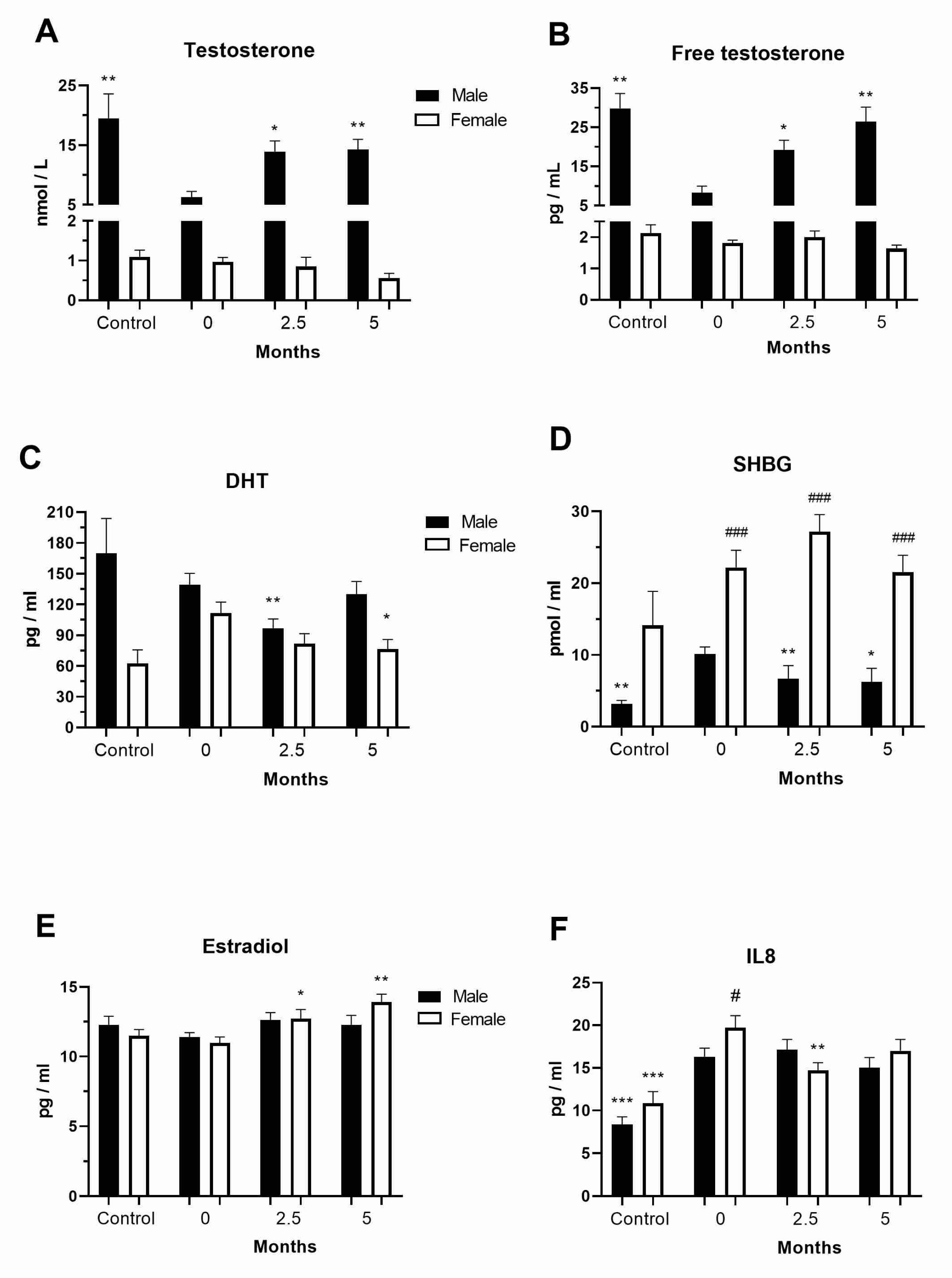
3.1. Levels of Sex Hormones in the COVID-19 and Post-COVID-19 Patients
3.2. Levels of Chemokines in the COVID-19 and Post-COVID-19 Patients
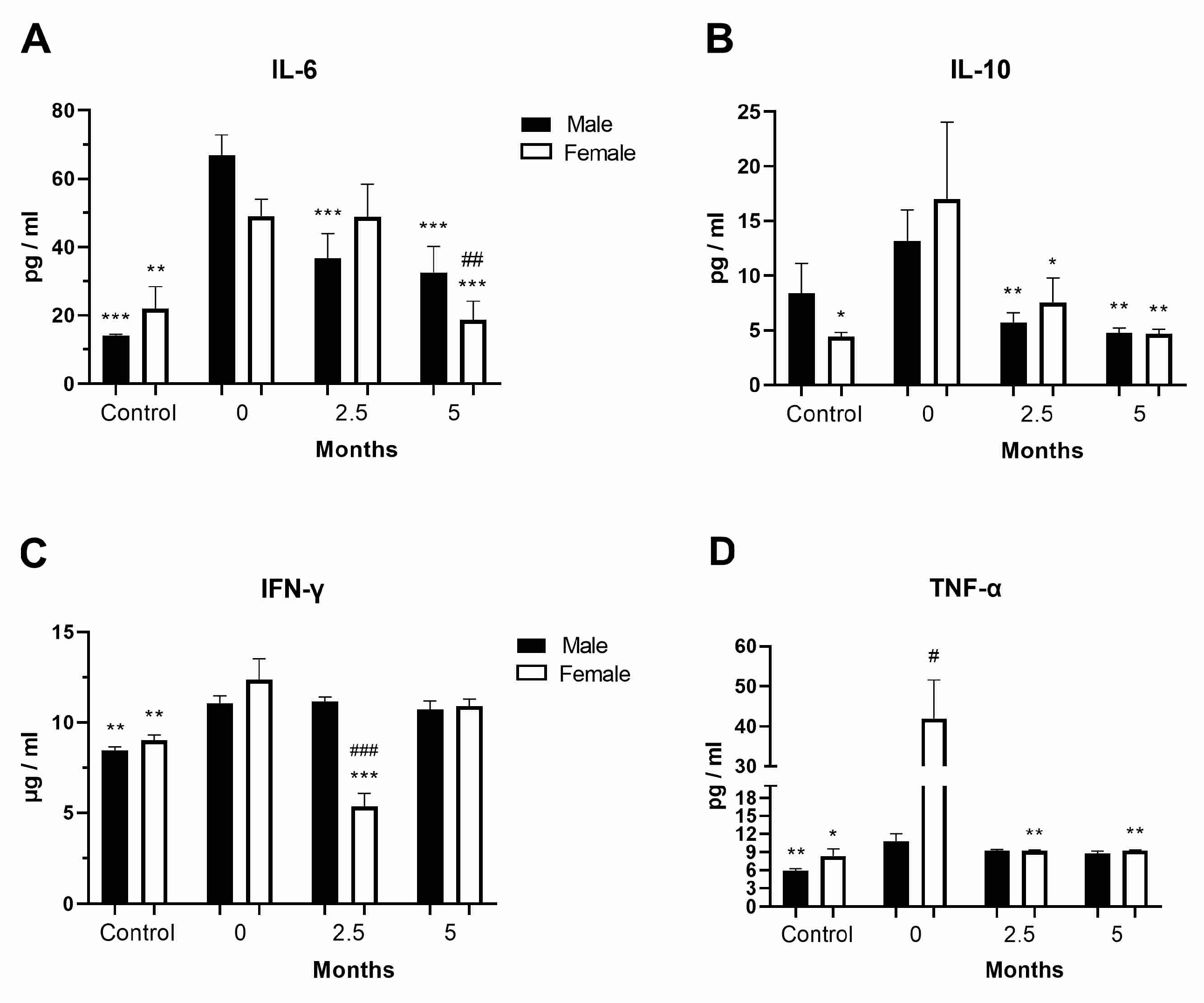
3.3. Levels of Inflammatory Cytokines in the COVID-19 and Post-COVID-19 Patients
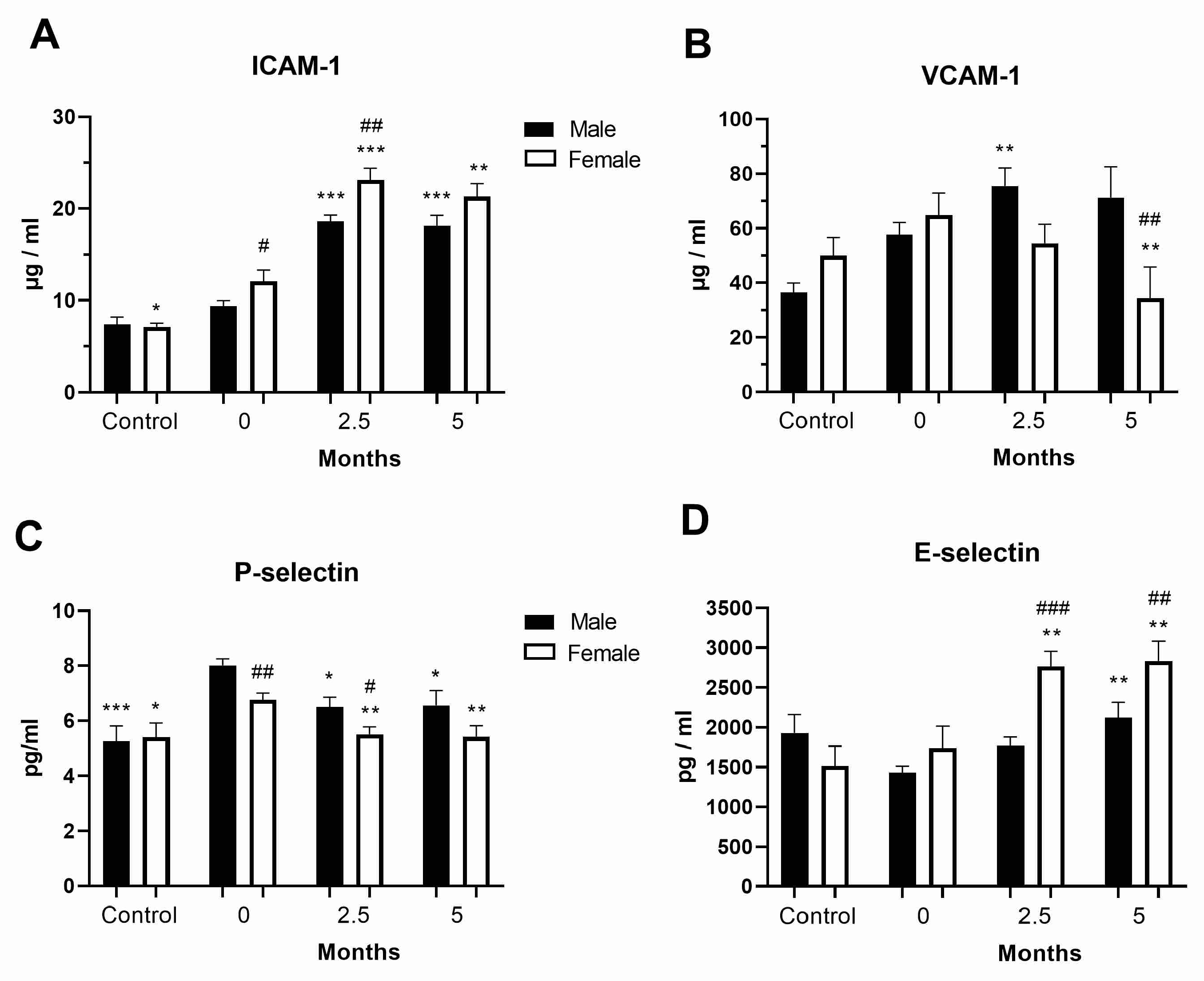
3.4. Levels of Cell Adhesion Molecules in the COVID-19 and Post-COVID-19 Patients
3.5. Reactive Oxygen Species Activity in Neutrophils
3.6. Quantification of NETs from the Plasma of Patients with COVID-19
3.7. Correlation of Biochemical Parameters with the Levels of Cell Adhesion Molecules and Sex Hormones in the COVID-19 and Post-COVID-19 Patients
3.8. Correlation of Vaccination and Biochemical Parameters with the Levels of Inflammatory Cytokines and Chemokines in COVID-19 and Post-COVID-19 Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yamamoto, S.; Shimizu, S.; Kiyonaka, S.; Takahashi, N.; Wajima, T.; Hara, Y.; Negoro, T.; Hiroi, T.; Kiuchi, Y.; Okada, T.; et al. TRPM2-mediated, Ca2+ influx induces chemokine production in monocytes that aggravates inflammatory neutrophil infiltration. Nat. Med. 2008, 14, 738–747. [Google Scholar] [CrossRef]
- Postiga, I.A.L.; Teixeira, P.C.; Neves, C.A.M.; Santana Filho, P.; Marmett, B.; Carvalho, R.; Peres, A.; Rotta, L.; Thompson, C.E.; Dorneles, G.P.; et al. Systemic redox imbalance in severe COVID-19 patients. Cell. Biochem. Funct. 2022, 40, 694–705. [Google Scholar] [CrossRef] [PubMed]
- Veras, F.P.; Pontelli, M.C.; Silva, C.M.; Toller-Kawahisa, J.E.; de Lima, M.; Nascimento, D.C.; Schneider, A.H.; Caetité, D.; Tavares, L.A.; Paiva, I.M.; et al. SARS-CoV-2-triggered neutrophil extracellular traps mediate COVID-19 pathology. J. Exp. Med. 2020, 217, e20201129. [Google Scholar] [CrossRef] [PubMed]
- Ng, H.; Havervall, S.; Rosell, A.; Aguilera, K.; Parv, K.; von Meijenfeldt, F.A.; Lisman, T.; Mackman, N.; Thålin, C.; Phillipson, M. Circulating markers of neutrophil extracellular traps are of prognostic value in patients with COVID-19. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 988–994. [Google Scholar] [CrossRef]
- Tong, M.; Jiang, Y.; Xia, D.; Xiong, Y.; Zheng, Q.; Chen, F.; Zou, L.; Xiao, W.; Zhu, Y. Elevated Expression of Serum Endothelial Cell Adhesion Molecules in COVID-19 Patients. J. Infect. Dis. 2020, 222, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Jain, V.; Kumar, P.; Panda, P.K.; Suresh, M.; Kaushal, K.; Mirza, A.A.; Raina, R.; Saha, S.; Omar, B.J.; Subbiah, V. Utility of IL-6 in the diagnosis, treatment and prognosis of COVID-19 patients: A longitudinal study. Vaccines 2022, 10, 1786. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef]
- Yin, S.W.; Zhou, Z.; Wang, J.L.; Deng, Y.F.; Jing, H.; Qiu, Y. Viral loads, lymphocyte subsets and cytokines in asymptomatic mildly and critical symptomatic patients with SARS-CoV-2 infection: A retrospective study. Virol. J. 2021, 18, 126. [Google Scholar] [CrossRef]
- Ghazavi, A.; Ganji, A.; Keshavarzian, N.; Rabiemajd, S.; Mosayebi, G. Cytokine profile and disease severity in patients with COVID-19. Cytokine 2021, 137, 155323. [Google Scholar] [CrossRef] [PubMed]
- Khalil, B.A.; Elemam, N.M.; Maghazachi, A.A. Chemokines and chemokine receptors during COVID-19 infection. Comput. Struct. Biotechnol. J. 2021, 19, 976–988. [Google Scholar] [CrossRef]
- Takahashi, T.; Ellingson, M.K.; Wong, P.; Israelow, B.; Lucas, C.; Klein, J.; Silva, J.; Mao, T.; Oh, J.E.; Tokuyama, M.; et al. Sex differences in immune responses that underlie COVID-19 disease outcomes. Nature 2020, 588, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Bienvenu, L.A.; Noonan, J.; Wang, X.; Peter, K. Higher mortality of COVID-19 in males: Sex differences in immune response and cardiovascular comorbidities. Cardiovasc. Res. 2020, 116, 2197–2206. [Google Scholar] [CrossRef] [PubMed]
- Rastrelli, G.; Di Stasi, V.; Inglese, F.; Beccaria, M.; Garuti, M.; Di Costanzo, D.; Spreafico, F.; Greco, G.F.; Cervi, G.; Pecoriello, A.; et al. Low testosterone levels predict clinical adverse outcomes in SARS-CoV-2 pneumonia patients. Andrology 2021, 9, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Enikeev, D.; Taratkin, M.; Morozov, A.; Petov, V.; Korolev, D.; Shpikina, A.; Spivak, L.; Kharlamova, S.; Shchedrina, I.; Mestnikov, O.; et al. Prospective two–arm study of the testicular function in patients with COVID-19. Andrology 2022, 10, 1047–1056. [Google Scholar] [CrossRef] [PubMed]
- Swerdloff, R.S.; Dudley, R.E.; Page, S.T.; Wang, C.; Salameh, W.A. Dihydrotestosterone: Biochemistry, physiology and clinical implications of elevated blood levels. Endocr. Rev. 2017, 38, 220–254. [Google Scholar] [CrossRef]
- Shepherd, R.; Cheung, A.S.; Pang, K.; Saffery, R.; Novakovic, B. Sexual Dimorphism in Innate Immunity: The Role of Sex Hormones and Epigenetics. Front. Immunol. 2021, 11, 604000. [Google Scholar] [CrossRef]
- Viveiros, A.; Rasmuson, J.; Vu, J.; Mulvagh, S.L.; Yip, C.Y.Y.; Norris, C.M.; Oudit, G.Y. Sex differences in COVID-19: Candidate pathways, genetics of ACE2, and sex hormones. Am. J. Physiol. Heart. Circ. Physiol. 2021, 320, H296–H304. [Google Scholar] [CrossRef]
- Scully, E.P.; Haverfield, J.; Ursin, R.L.; Tannenbaum, C.; Klein, S.L. Considering how biological sex impacts immune responses and COVID-19 outcomes. Nat. Rev. Immunol. 2020, 20, 442–447. [Google Scholar] [CrossRef]
- Gebhard, C.; Regitz-Zagrosek, V.; Neuhauser, H.K.; Morgan, R.; Klein, S.L. Impact of sex and gender on COVID-19 outcomes in Europe. Biol. Sex. Differ. 2020, 11, 29. [Google Scholar] [CrossRef]
- Hikmet, F.; Méar, L.; Edvinsson, Å.; Micke, P.; Uhlén, M.; Lindskog, C. The protein expression profile of ACE2 in human tissues. Mol. Syst. Biol. 2020, 16, e9610. [Google Scholar] [CrossRef]
- Flaifel, A.; Guzzetta, M.; Occidental, M.; Najari, B.B.; Melamed, J.; Thomas, K.M.; Deng, F.M. Testicular changes associated with severe acute respiratory syndrome Coronavirus 2 (SARS-CoV-2). Arch. Pathol. Lab. Med. 2021, 145, 8–9. [Google Scholar] [CrossRef] [PubMed]
- Dhindsa, S.; Champion, C.; Deol, E.; Lui, M.; Campbell, R.; Newman, J.; Yeggalam, A.; Nadella, S.; Ahir, V.; Shrestha, E.; et al. Association of male hypogonadism with risk of hospitalization for COVID-19. JAMA Netw. Open. 2022, 5, e2229747. [Google Scholar] [CrossRef] [PubMed]
- Çayan, S.; Uğuz, M.; Saylam, B.; Akbay, E. Effect of serum total testosterone and its relationship with other laboratory parameters on the prognosis of coronavirus disease 2019 (COVID-19) in SARS–CoV–2 infected male patients: A cohort study. Aging Male. 2020, 23, 1493–1503. [Google Scholar] [CrossRef] [PubMed]
- Lanser, L.; Burkert, F.R.; Thommes, L.; Egger, A.; Hoermann, G.; Kaser, S.; Pinggera, G.M.; Anliker, M.; Griesmacher, A.; Weiss, G.; et al. Testosterone deficiency is a risk factor for severe COVID-19. Front. Endocrinol. 2021, 12, 694083. [Google Scholar] [CrossRef]
- Schroeder, M.; Schaumburg, B.; Mueller, Z.; Parplys, A.; Jarczak, D.; Roedl, K.; Nierhaus, A.; de Heer, G.; Grensemann, J.; Schneider, B.; et al. High estradiol and low testosterone levels are associated with critical illness in male but not in female COVID-19 patients: A retrospective cohort study. Emerg. Microbes. Infect. 2021, 10, 1807–1818. [Google Scholar] [CrossRef]
- Sharma, A.; Jayasena, C.N.; Dhillo, W.S. Regulation of the Hypothalamic–Pituitary–Testicular Axis: Pathophysiology of Hypogonadism. Endocrinol. Metab. Clin. North. Am. 2022, 51, 29–45. [Google Scholar] [CrossRef]
- Cai, Z.; Zhong, J.; Jiang, Y.; Zhang, J. Associations between COVID-19 infection and sex steroid hormones. Front. Endocrinol. 2022, 13, 940675. [Google Scholar] [CrossRef]
- Van Zeggeren, I.E.; Boelen, A.; van de Beek, D.; Heijboer, A.C.; Vlaar, A.P.J.; Brouwer, M.C.; Amsterdam UMC COVID-19 Biobank. Sex steroid hormones are associated with mortality in COVID-19 patients: Level of sex hormones in severe COVID-19. Medicine 2021, 100, e27072. [Google Scholar] [CrossRef]
- Paoli, D.; Pallotti, F.; Anzuini, A.; Bianchini, S.; Caponecchia, L.; Carraro, A.; Ciardi, M.R.; Faja, F.; Fiori, C.; Gianfrilli, D.; et al. Male reproductive health after 3 months from SARS-CoV-2 infection: A multicentric study. J. Endocrinol Investig. 2023, 46, 89–101. [Google Scholar] [CrossRef]
- Aribas, E.; Roeters van Lennep, J.E.; De Rijke, Y.B.; Laven, J.S.E.; Ikram, M.A.; Peeters, R.P.; Kavousi, M. Sex steroids and sex steroid-binding globulin levels amongst middle-aged and elderly men and women from general population. Eur. J. Clin. Investig. 2022, 52, e13866. [Google Scholar] [CrossRef]
- Infante, M.; Pieri, M.; Lupisella, S.; D’Amore, L.; Bernardini, S.; Fabbri, A.; Iannetta, M.; Andreoni, M.; Morello, M. Low testosterone levels and high estradiol to testosterone ratio are associated with hyperinflammatory state and mortality in hospitalized men with COVID-19. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 5889–5903. [Google Scholar] [PubMed]
- Salonia, A.; Pontillo, M.; Capogrosso, P.; Pozzi, E.; Ferrara, A.M.; Cotelessa, A.; Belladelli, F.; Corsini, C.; Gregori, S.; Rowe, I.; et al. Testosterone in males with COVID-19: A 12-month cohort study. Andrology 2022, 11, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Amodio, G.; Capogrosso, P.; Pontillo, M.; Tassara, M.; Boeri, L.; Carenzi, C.; Cignoli, D.; Ferrara, A.M.; Ramirez, G.A.; Tresoldi, C.; et al. Combined plasma levels of IL-10 and testosterone, but not soluble HLA-G5, predict the risk of death in COVID-19 patients. Andrology 2023, 11, 32–44. [Google Scholar] [CrossRef]
- Tramontana, F.; Battisti, S.; Napoli, N.; Strollo, R. Immuno-Endocrinology of COVID-19: The Key Role of Sex Hormones. Front. Endocrinol. 2021, 12, 726696. [Google Scholar] [CrossRef]
- Kumar, P.; Osahon, O.; Vides, D.B.; Hanania, N.; Minard, C.G.; Sekhar, R.V. Severe Glutathione Deficiency Oxidative Stress and Oxidant Damage in Adults Hospitalized with COVID-19: Implications for GlyNAC (Glycine and N-Acetylcysteine) Supplementation. Antioxidants 2021, 11, 50. [Google Scholar] [CrossRef]
- Montiel, V.; Lobysheva, I.; Gérard, L.; Vermeersch, M.; Perez–Morga, D.; Castelein, T.; Mesland, J.B.; Hantson, P.; Collienne, C.; Gruson, D.; et al. Oxidative stress–induced endothelial dysfunction and decreased vascular nitric oxide in COVID-19 patients. EBioMedicine 2022, 77, 103893. [Google Scholar] [CrossRef] [PubMed]
- Middleton, E.A.; He, X.Y.; Denorme, F.; Campbell, R.A.; Ng, D.; Salvatore, S.P.; Mostyka, M.; Baxter–Stoltzfus, A.; Borczuk, A.C.; Loda, M.; et al. Neutrophil extracellular traps contribute to immunothrombosis in COVID-19 acute respiratory distress syndrome. Blood 2020, 136, 1169–1179. [Google Scholar] [CrossRef]
- Huckriede, J.; de Vries, F.; Hultström, M.; Wichapong, K.; Reutelingsperger, C.; Lipcsey, M.; Garcia de Frutos, P.; Frithiof, R.; Nicolaes, G.A.F. Histone H3 Cleavage in Severe COVID-19 ICU Patients. Front. Cell Infect. Microbiol. 2021, 11, 694186. [Google Scholar] [CrossRef] [PubMed]
- Skendros, P.; Mitsios, A.; Chrysanthopoulou, A.; Mastellos, D.C.; Metallidis, S.; Rafailidis, P.; Ntinopoulou, M.; Sertaridou, E.; Tsironidou, V.; Tsigalou, C.; et al. Complement and tissue factor-enriched neutrophil extracellular traps are key drivers in COVID-19 immunothrombosis. J. Clin. Investig. 2020, 130, 6151–6157. [Google Scholar] [CrossRef]
- Goshua, G.; Pine, A.B.; Meizlish, M.L.; Chang, C.H.; Zhang, H.; Bahel, P.; Baluha, A.; Bar, N.; Bona, R.D.; Burns, A.J.; et al. Endotheliopathy in COVID-19-associated coagulopathy: Evidence from a single-centre cross-sectional study. Lancet Haematol. 2020, 7, e575–e582. [Google Scholar] [CrossRef]
- Fan, B.E.; Wong, S.W.; Sum, C.L.L.; Lim, G.H.; Leung, B.P.; Tan, C.W.; Ramanathan, K.; Dalan, R.; Cheung, C.; Lim, X.R.; et al. Hypercoagulability, endotheliopathy and inflammation approximating 1 year after recovery: Assessing the long-term outcomes in COVID-19 patients. Am. J. Hematol. 2022, 97, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Watany, M.M.; Abdou, S.; Elkolaly, R.; Elgharbawy, N.; Hodeib, H. Evaluation of admission levels of P, E and L selectins as predictors for thrombosis in hospitalized COVID-19 patients. Clin. Exp. Med. 2022, 22, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Ben–Batalla, I.; Vargas-Delgado, M.E.; von Amsberg, G.; Janning, M.; Loges, S. Influence of Androgens on Immunity to Self and Foreign: Effects on Immunity and Cancer. Front. Immunol. 2020, 11, 1184. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COVID-19 | Sex | INR | PTS | UREA mmol/L | CRT μmol/L | CK U/L | D- DIMER mg/L | CRP mg/L | AST U/L | ALT U/L | GGT U/L | FBN g/L | LDH U/L | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Diagnosis | M | AV | 1.20 | 12.98 | 6.70 | 93.19 | 380.08 | 21.38 | 90.56 | 59.80 | 74.06 | 65.42 | 5.91 | 318.85 |
| SD | 1.07 | 1.03 | 2.64 | 33.88 | 589.24 | 146.6 | 68.46 | 50.97 | 56.76 | 58.83 | 2.12 | 138.45 | ||
| M * | 1.08 | 13 | 6.05 | 95 | 157 | 0.75 | 82.9 | 42 | 55.5 | 50 | 5.4 | 282 | ||
| F | AV | 1.11 | 13.00 | 6.37 | 81.20 | 122.00 | 2.18 | 63.62 | 42.80 | 45.08 | 43.72 | 5.13 | 268.49 | |
| SD | 0.28 | 1.58 | 4.79 | 50.03 | 104.77 | 7.65 | 48.49 | 36.42 | 27.84 | 27.05 | 1.67 | 96.01 | ||
| M * | 1.05 | 12.7 | 5.4 | 74.5 | 77 | 0.72 | 47.5 | 31 | 37 | 36 | 4.7 | 258.5 | ||
| 2.5 months | M | AV | 2.56 3 | 11.95 3 | 6.91 | 93.75 3 | 131.5 3 | 0.56 3 | 5.40 3 | 23.15 3 | 48.61 3 | 37.15 3 | 3.17 3 | 179.18 3 |
| SD | 12.32 | 1.06 | 9.30 | 26.19 | 153.62 | 0.65 | 11.78 | 11.98 | 35.17 | 21.50 | 1.20 | 47.60 | ||
| M * | 0.99 | 11.8 | 5.85 | 91 | 104 | 0.36 | 1.2 | 20.5 | 35 | 31 | 2.9 | 172 | ||
| F | AV | 0.99 3 | 12.25 3 | 5.51 | 70.40 | 89.46 | 0.59 1 | 4.81 3 | 21.10 3 | 34.07 | 33.52 1 | 3.87 3 | 173.75 3 | |
| SD | 0.08 | 2.05 | 1.79 | 17.13 | 61.57 | 0.38 | 6.46 | 8.75 | 13.90 | 18.13 | 1.40 | 48.39 | ||
| M * | 0.97 | 11.70 | 5.15 | 70.50 | 70.50 | 0.49 | 1.70 | 18 | 31 | 28 | 3.45 | 170 | ||
| 5 months | M | AV | 0.98 3 | 11.71 3 | 6.40 | 92.10 | 163.44 | 0.55 3 | 6.09 3 | 25.71 3 | 47.88 3 | 32.28 3 | 2.96 3 | 166.82 3 |
| SD | 0.07 | 0.72 | 1.98 | 27.23 | 153.89 | 0.78 | 14.69 | 20.06 | 43.22 | 11.70 | 1.11 | 32.02 | ||
| M * | 0.97 | 11.6 | 6 | 87 | 116.5 | 0.33 | 1.6 | 21 | 33 | 29 | 2.7 | 159 | ||
| F | AV | 1.03 3 | 12.36 3 | 5.85 | 60.27 2 | 95.50 | 3.84 1 | 5.38 3 | 17.96 3 | 29.67 1 | 26.50 3 | 3.77 3 | 153.24 3 | |
| SD | 0.26 | 2.93 | 1.72 | 19.43 | 50.59 | 14.88 | 12.05 | 6.37 | 11.03 | 17.84 | 1.61 | 42.40 | ||
| M * | 0.97 | 11.7 | 5.55 | 60 | 87 | 0.425 | 2.25 | 17 | 26 | 19.5 | 3.2 | 160 | ||
| Normal range | 0.8–1.1 | 11–13.5 | 2.5–7.5 | 59–104 | 0–200 | 0.5 | 0–3 | 0–37 | 14–63 | 0–55 | 1.8–3.5 | 85–227 | ||
| p Values | Sex | INR | PTS | Urea | Creatinine | CK | D-dimer | CRP | AST | ALT | GGT | Fibrinogen | LDH | Vaccine | CO | Co-M |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E-SELECTIN | F | 0.015 2 | 0.006 1 | 0.048 0.001 2 | 0.021 | |||||||||||
| M | 0.009 2 | 0.025 2 | ||||||||||||||
| T | 0.033 | 0.048 | 0.023 | 0.029 | 0.026 0.021 1 | |||||||||||
| P- SELECTIN | F | 0.000 1 | 0.049 | 0.005 | 0.044 | 0.005 | ||||||||||
| M | 0.040 2 | 0.024 2 | ||||||||||||||
| T | 0.022 2 | 0.031 | ||||||||||||||
| VCAM-1 | F | 0.046 1 | 0.030 2 | 0.037 | ||||||||||||
| M | 0.020 2 | 0.037 2 | 0.048 2 | 0.001 1 | 0.022 1 | |||||||||||
| T | 0.007 2 | 0.007 2 | 0.008 2 | 0.023 2 | 0.008 1 | |||||||||||
| ICAM-1 | F | 0.043 1 | ||||||||||||||
| M | 0.032 | 0.024 1 | ||||||||||||||
| T | 0.005 1 | |||||||||||||||
| ESTRADIOL | F | 0.013 1 | 0.009 1 | 0.011 1 | 0.016 1 | 0.030 1 | ||||||||||
| M | 0.002 2 | 0.006 1 0.018 2 | ||||||||||||||
| T | 0.046 1 | |||||||||||||||
| DHT | F | 0.017 | 0.025 | 0.028 | ||||||||||||
| M | 0.036 | 0.028 | ||||||||||||||
| T | 0.004 0.018 2 | 0.013 2 | 0.049 2 | 0.005 1 | ||||||||||||
| SHBG | F | 0.046 | 0.003 1 | 0.024 | 0.037 1 | |||||||||||
| M | 0.035 | 0.011 2 | ||||||||||||||
| T | 0.001 0.030 1 | 0.048 1 | 0.003 | |||||||||||||
| INF-γ | F | 0.021 2 | 0.001 1 | 0.03 1 | ||||||||||||
| M | 0.015 | 0.006 2 | 0.041 1 0.040 2 | |||||||||||||
| T | 0.001 1 | 0.015 | 0.014 | |||||||||||||
| TNF-α | F | 0.028 1 | ||||||||||||||
| T | 0.016 1 | |||||||||||||||
| IL-10 | F | 0.012 1 | 0.011 2 | 0.037 2 | ||||||||||||
| T | 0.028 2 | |||||||||||||||
| IL-6 | F | 0.04 | ||||||||||||||
| M | 0.035 2 | |||||||||||||||
| T | 0.024 2 | 0.011 2 | 0.004 2 | |||||||||||||
| IL-1β | F | 0.003 1 | ||||||||||||||
| M | 0.006 1 | 0.027 1 | ||||||||||||||
| T | 0.004 1 | |||||||||||||||
| MCP-1 | F | 0.027 | ||||||||||||||
| M | 0.022 1 | 0.013 2 | ||||||||||||||
| T | 0.003 1 | 0.004 2 | 0.019 2 | |||||||||||||
| TGF-β | M | 0.016 | ||||||||||||||
| T | 0.021 2 | 0.012 2 | 0.042 1 | 0.046 | ||||||||||||
| IL-8 | F | 0.032 | ||||||||||||||
| M | 0.029 | 0.047 2 | ||||||||||||||
| T | 0.0292 | 0.007 | 0.036 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mitrović-Ajtić, O.; Đikić, D.; Subotički, T.; Bižić-Radulović, S.; Beleslin-Čokić, B.; Dragojević, T.; Živković, E.; Miljatović, S.; Vukotić, M.; Stanisavljević, D.; et al. Sex Differences and Cytokine Profiles among Patients Hospitalized for COVID-19 and during Their Recovery: The Predominance of Adhesion Molecules in Females and Oxidative Stress in Males. Vaccines 2023, 11, 1560. https://doi.org/10.3390/vaccines11101560
Mitrović-Ajtić O, Đikić D, Subotički T, Bižić-Radulović S, Beleslin-Čokić B, Dragojević T, Živković E, Miljatović S, Vukotić M, Stanisavljević D, et al. Sex Differences and Cytokine Profiles among Patients Hospitalized for COVID-19 and during Their Recovery: The Predominance of Adhesion Molecules in Females and Oxidative Stress in Males. Vaccines. 2023; 11(10):1560. https://doi.org/10.3390/vaccines11101560
Chicago/Turabian StyleMitrović-Ajtić, Olivera, Dragoslava Đikić, Tijana Subotički, Sandra Bižić-Radulović, Bojana Beleslin-Čokić, Teodora Dragojević, Emilija Živković, Sanja Miljatović, Milica Vukotić, Dejana Stanisavljević, and et al. 2023. "Sex Differences and Cytokine Profiles among Patients Hospitalized for COVID-19 and during Their Recovery: The Predominance of Adhesion Molecules in Females and Oxidative Stress in Males" Vaccines 11, no. 10: 1560. https://doi.org/10.3390/vaccines11101560