1. Introduction
Salmonella is an important pathogenic bacterium in the poultry industry, public health, and human food safety worldwide. It is implicated in both animal and human salmonellosis cases. Unlike their typhoidal counterparts (
Salmonella enterica serovar Gallinarum and
Salmonella enterica serovar Pullorum) which are host-specific, non-typhoidal Salmonella (
Salmonella enterica serovar Enteritidis,
Salmonella enterica serovar Typhimurium,
Salmonella enterica serovar Infantis) can infect a range of hosts and are of zoonotic importance, being associated with human food poisoning cases. Infection with
Salmonella in humans occurs upon consumption of raw or undercooked poultry products, mostly meat and eggs, contaminated with the pathogen [
1]. In young chicks, whose immunity is still poorly developed and whose gut microbiome is immature, infection with non-typhoidal
Salmonella enterica can be systemic and deadly. Healthy older chickens, however, remain asymptomatic upon infection, silently propagating the pathogen in the flock, contaminating the products thereof and increasing the incidences of human salmonellosis [
2,
3,
4,
5]. Poultry is thus an important reservoir for non-typhoidal
Salmonella and control measures which limit their contamination with this pathogen at the farm level would greatly contribute to a reduction in human non-typhoidal salmonellosis cases [
6].
Generally, the most commonly used prophylaxes in poultry farms against
Salmonella involve biosecurity measures, flock testing and culling, use of antibiotics, and vaccination programs. For decades, the chemoprophylactic use of antibiotics as feed additives has been widely practiced and with considerable success in limiting intestinal pathogenic bacterial infections [
7,
8]. The increasing global problem of antimicrobial resistance, however, has led to the banning of antibiotics as growth promoters in poultry production in most countries. In Europe, this has been in effect since 2006 under the Commission regulation (EC) No 1177/2006 [
9]. Consequently, non-antibiotic alternatives for the control of important pathogenic bacteria such as
Salmonella in poultry are urgently needed. Prebiotics, probiotics, bacteriophages, phytobiotics and vaccines are among the developed non-antibiotic prophylactic measures. The mechanism of action of most of these new anti-
Salmonella strategies are, however, not fully understood [
10,
11].
Nevertheless, most of the developed anti-
Salmonella alternatives are administered orally to effectively reach the gut, which is the main route for
Salmonella entry in chicken. Successful colonization of the chicken gut by non-typhoidal
Salmonella involves induction of inflammation by invasion of the epithelial cells. This also generates metabolites such as tetrathionate, which acts as a terminal electron acceptor needed for ethanolamine and 1,2-propane diol utilization under anaerobic conditions [
12]. From this,
Salmonella gains a competitive advantage over the commensal gut bacteria. Consequently, this leads to an imbalanced gut microbiota, allowing for colonization of the chicken gut by
Salmonella. Thus, control approaches that lead to a strengthened or balanced protective microbiota in the chicken gut are essential to limiting
Salmonella contamination level in chickens. The commensal gut microbiota play an important role in protecting chickens against
Salmonella via mechanisms such as modulation of the host immune responses, colonization exclusion, short chain fatty acids (SCFA) release from the metabolism of non-fermentable carbon sources and antimicrobial peptides (such as bacteriocins) production. The latter two are, respectively, bacteriostatic and bactericidal to
Salmonella [
10,
13].
Live bacteria prophylactics, involving the use of attenuated
Salmonella vaccines and live bacteria with protective benefits (probiotics) to the chicken gut are commonly used methods in modern poultry farming. The varying efficacy of live
Salmonella vaccines in protecting chickens against the challenge of wild-type strains has been reported [
14]. Nevertheless, vaccination is still considered the most efficient approach to control
Salmonella contamination in chickens at the farm level [
15]. Both inactivated and live attenuated
Salmonella vaccines for poultry are commercially available and used in poultry farms. Whereas inactivated vaccines mainly induce humoral immunity, live attenuated
Salmonella vaccines can trigger both cellular and humoral immune responses, offering more protection in poultry [
14]. Cellular responses are also associated with an enhanced expression of interferon gamma (IFNγ), interleukin 8 (IL-8) and inducible nitric oxide synthase (iNOS) while downregulating interleukin 1β (IL-1β) [
16].
The effect of live attenuated
Salmonella vaccines on the poultry gut microbiota is not well established. A recent study by Park et al. [
17], using
Salmonella Typhimurium live attenuated vaccine candidates, reported compositional changes but not overall relative abundance in gut microbiota. To our knowledge, reports on other live attenuated
Salmonella vaccine types, particularly multivalent vaccines, are not available. In this study, we used three commercial live attenuated
Salmonella vaccine strains for
S. Typhimurium (AviPro
® Salmonella Vac T),
S. Enteritidis (AviPro
® Salmonella Vac E) and
S. Typhimurium +
S. Enteritidis (AviPro
® Salmonella DUO) to study their influence on cecal microbiota profiles. The cecal innate immune genes expressions and
Salmonella-specific humoral responses (IgG and IgA) were also investigated in both vaccinated and unvaccinated chickens.
2. Materials and Methods
2.1. Experimental Animals Description
A total of 16 day-old male Ross-308 broiler chicks were obtained from a commercial supplier (Belgabroed nv, Merksplas, Belgium). The chicks used for this study were pre-vaccinated for Newcastle disease by the supplier via spray with Nobilis® ND C2 (MSD Animal Health, Madison, NJ, USA), a live Newcastle disease vaccine. The use of animals for this study was evaluated and approved by the KU Leuven Ethical Committee for Animal Research, project number P040/2020.
2.2. Animal Handling Procedures and Housing
The experiment was carried out at TRANSfarm, the test facility of KU Leuven, located in Lovenjoel, Belgium. The chicks were housed in 4 pens (4 chicks/pen) with the floor covered by wood shavings. A starter diet and drinking water were provided ad libitum in a room with standard heating and light-dark cycles.
2.3. Experimental Setup and Treatments
Day-old broiler chicks were divided into four treatment groups: S. Typhimurium (AviPro® Salmonella Vac T), S. Enteritidis (AviPro® Salmonella Vac E), S. Typhimurium + S. Enteritidis (AviPro® Salmonella DUO) and the control group, each consisting of four chicks. The live attenuated Salmonella vaccines (AviPro® Salmonella Vac T, AviPro® Salmonella Vac E and AviPro® Salmonella DUO) used in this study were all obtained from Elanco Europe Ltd. (Bartley Wood Business Park, Hook, UK). At 2 days of age, cloacal swabs were collected from all chicks to check for the presence of Salmonella contamination prior to administration of the treatments. Then, each group received 0.5 mL of either a live attenuated Salmonella vaccine or sterile normal saline (for the control) by oral gavage. On day 14 post-vaccination (DPV14, 16 days post hatch), the experiment was terminated, and samples were collected.
2.4. Sample Collection and Processing
DPV14 (16 days post hatch), chickens were euthanized by cervical dislocation, dissected, ceca and cecal contents were aseptically collected into sterile 2 mL cryovials (SARSTEDT, Nümbrecht, Germany), immediately placed on dry ice for transportation and then stored at −80 °C until use. One chick from the
S. Enteritidis (AviPro
® Salmonella Vac E) group could, however, not be sampled as it died four days after vaccination and was thus excluded from further analysis. The specific cause of the chick’s death was not clear, as the bird did not show any visible physical indications of disease or deformation. Nevertheless, failure of adaptation to the rapid changes that occur during the first week of the chick’s life, where their immunity is still immature (transportation stress, feed, water, environmental, and microbiological changes), is the suspected cause. Prior to humane killing, whole blood was withdrawn from each chicken’s brachial wing vein into BD Vacutainer™ SST™ serum separator tubes (Fisher Scientific, Hampton, NH, USA) for serum collection. Serum was prepared by centrifugation at 2000×
g for 10 min at room temperature, collecting a clear supernatant into new tubes and storing at −20 °C until needed. The sampling in this study was intended to capture the cecal immune responses in the function of live
Salmonella vaccines and the vaccines interaction with commensal microbiota. Thus, the collection of samples was performed towards the broilers gut microbiota stabilizing period, which is suggested to be 14–21 days post hatch [
18].
2.5. Total Tissue RNA Extraction and Complementary DNA (cDNA) Synthesis
Total RNA was isolated from cecal tissues using the TRIzol™ (Invitrogen™, Waltham, MA, USA) method, following the manufacturer’s instructions. Briefly, approximately 80 mg cecal tissues were homogenized in TRIzol™ (Invitrogen) for 1 min at 6800 rpm, in the presence of sterile 2 mm Zirconia Beads (BioSpec, Bartlesville, OK, USA), using the Precellys® Evolution tissue homogenizer (Bertin instruments, Montigny-le-Bretonneux, France). Chloroform (Avantor, Radnor, PA, USA) was added to tissue homogenates, followed by centrifugation for 15 min at 12,000× g, 4 °C to separate the aqueous from the organic phase. Total RNA was precipitated from the aqueous phase by addition of isopropanol (Carl Roth, Karlsruhe, Schoemperlenstraße, Germany), followed by centrifugation for 10 min at 12,000× g, 4 °C. The RNA pellet was washed and resuspended in 75% ethanol (VWR, Geldenaaksebaan, Leuven, Belgium), vortexed briefly and centrifuged for 5 min at 7500× g, 4 °C. The supernatant was discarded with a micropipettor, and the RNA pellet allowed to air-dry for 10 min. The RNA pellet was finally solubilized via resuspension in 40 µL RNase-free water through gentle pipetting. The concentration and purity (A260/A280 ratio) of the RNA samples were checked using a SimpliNanoTM spectrophotometer (BioChrom, Hill Road Holliston, MA, USA), then stored at −80 °C until needed.
The cDNA was synthesized from 200 ng cecal total tissue RNA per sample, using oligo (dT) primers (Promega, Madison, Wisconsin, MA, USA) and the Maxima H minus reverse transcriptase enzyme (ThermoFisher, Waltham, MA, USA), following the manufacturer’s protocol. In brief, the cDNA synthesis was performed at 50 °C for 30 min in the presence of 200U Maxima H minus reverse transcriptase (ThermoFisher Scientific) and 20U RiboLock RNase inhibitor (ThermoFisher Scientific). The reaction was then terminated by heating at 85 °C, and samples were stored at −20 °C until use.
2.6. qPCR
A probe-based qPCR was performed using the GoTaq probe two-step-qPCR system (Promega), according to the manufacturer’s recommendations. The PCR reactions consisted of 10 µL GoTaq probe master mix (2X), 0.4 µL each primer per target (200 nM), 0.4 µL hydrolysis probe (200 nM), cDNA (70 ng per sample) and nuclease-free water to a final 20 µL reaction volume. The reactions were prepared in MicroAmp
TM Fast Optical 96-well reaction plates and sealed with MicroAmp
TM optical adhesive films (ThermoFisher Scientific). The PCR run was performed in the Applied Biosystems StepOne Plus (ThermoFisher Scientific) using the standard cycling conditions; 50 °C for 2 min, 95 °C for 2 min, then followed by 40 cycles of 95 °C for 15 s and 60 °C for 1 min. The primer pairs and hydrolysis probes used for this study are given in
Table 1. The design of primers and probes and the checking of their parameters were performed using the integrated DNA technology (IDT) OligoAnalyzer
TM tool (IDT, Leuven, Belgium) and were ordered from the same manufacturer.
2.7. ELISA
An overnight culture of a live attenuated Salmonella vaccine strain (AviPro® Salmonella DUO) was pelleted at 10,000× g (4 °C) for 5 min and washed twice with PBS at the same speed, each time for 1 min. The bacterial pellet was resuspended in PBS and absorbance (O.D600) was adjusted to 1, using a nanospectrophotometer (Westburg Life Sciences, Leusden-zuid, Utrecht, The Netherlands). This was then distributed in 96-well F-bottom, clear, high binding microplates (Greiner Bio-One, Kremsmünster, Austria) at 50 µL per well as a Salmonella vaccine coating solution. Plates were covered with lids, sealed with parafilm to prevent evaporation and incubated overnight at 4 °C to allow for coating. The coating solution was discarded and each well blocked with 50 µL blocking buffer (PBS + 2% BSA + 1% goat serum) for 1.5 h at room temperature. The blocking buffer was then discarded and each well rinsed twice with washing buffer (PBS + 0.05% Tween-20). Prior to use, the standard chicken sera and cecal extract samples were thawed and spun down at 11,000× g (1 min, room temperature). The standard samples were derived from 53 week-old Isa Brown layer chickens, obtained from Evap Proefbedrijf Pluimveehouderij (Provincie Antwerpen, Belgium) and were fully vaccinated orally with the AviPro® Salmonella DUO live attenuated vaccine. For cecal extracts preparation, 0.5 g cecal contents were homogenized in 500 µL extraction buffer (PBS + 0.02% sodium azide) by continuously vortexing for 15 min. The mixture was then centrifuged at 7000× g for 20 min (4 °C) and clear supernatant collected. Prior to use as standards, chicken sera and cecal extracts were tested for high absorbances (antibodies) by ELISA and then aliquoted into 200 µL and stored at −20 °C until needed. Fifty microliters of the diluted standard and test samples were pipetted into each well of the coated plates as primary antibodies and incubated at room temperature for 1 h. The unbound antibodies were washed by rinsing 3 times with washing buffer, followed by thorough blotting on paper towel. Fifty microliters horse-radish peroxidase (HRP)-conjugated rabbit anti-chicken IgG (EMD Millipore AP162P) or goat anti-chicken IgA (Bio-Rad, Hercules, CA, USA), were added as secondary antibodies (1:10,000, in blocking buffer) and plates incubated at room temperature for 30 min. Unbound secondary antibodies were washed by rinsing 4 times and thorough blotting of the plates on paper towel. To each well, 50 µL 3,3′5,5′-tetramethylbenzidine (TMB) developing substrate (ThermoFisher Scientific) was pipetted and plates incubated at room temperature for 10 min. The enzymatic reaction was finally stopped using 50 µL 0.18 M sulfuric acid and absorbance immediately measured at 450 nm using the Victor3 microplate reader (Perkin Elmer, Waltham, MA, USA).
The antibody units were assigned to standard and test samples as previously described by Miura et al. [
27], with some modifications. Briefly, an aliquot of the standard serum sample was thawed, centrifuged at 11,000×
g for 1 min and diluted in 2-fold steps from 1:100 to 1:51,200 in blocking buffer. For cecal extracts, the thawed aliquot was centrifuged for 1 min at 7000×
g, then diluted in ten 2-fold steps from 1:10 to 1:10,240. The serially diluted standards were applied on ELISA plates as primary antibodies, except for two wells per set of serially diluted standard which were left as blank wells and assigned the reciprocal number of dilution 0. The relation between reciprocal number of dilution and absorbance (OD
450) was approximated using MyCurveFit (
https://mycurvefit.com/, accessed on 15 March 2023) [
28], an online 4-parameter hyperbolic curve fitting software. As a quality check, only standard samples giving R
2 ≥ 0.994 were used for further analyses of the test samples. The constants of the hyperbolic curve fit equation were used to assign antibody units to the standard as reciprocal of the dilution giving an OD
450 = 1. After determining the antibody units of the standard samples, the number was used for all samples tested by ELISA against that standard. Such a reference standard was then used on each ELISA plate to make a standard curve. To this end, standard serum and cecal extract aliquots were thawed and used to prepare ten 2-fold dilution steps in duplicates serially, starting with a dilution of 20 antibody units (serum) and 2 antibody units (cecal extracts). The serially diluted reference standards and test samples were applied to the assigned wells on ELISA plates as primary antibodies and four were left as blank wells (antibody units assigned as 0). The absorbance (OD
450) values obtained were fitted to a 4-parameter standard curve (Antibody units = a[{(a − OD
450)/(OD
450 − d)}˄(1/b)]), using an online 4-parameter hyperbolic curve software (MyCurveFit). The generated standard curves were then used to calculate antibody units in test samples from their measured absorbance values. The antibody units in undiluted samples were finally determined based on the dilution factor for each sample.
2.8. Total Genomic DNA (gDNA) Isolation for 16S rRNA Sequencing
Genomic DNA was isolated from the chicken cecal contents using the QIAamp® Fast DNA Stool Mini kit (Qiagen, Hilden, Germany), according to the manufacturer’s protocol. The concentration and purity of the isolated genomic DNA from cecal contents were checked using the SimpliNanoTM spectrophotometer (BioChrom), and the A260/A280 ratios were within the recommended range (1.8–2.0) for pure DNA. The DNA samples were then stored at −20 °C until use.
2.9. Sample Preparation and 16S rRNA Next Generation Sequencing
DNA concentration measurements were performed using the Qubit double-stranded DNA (dsDNA) high-sensitivity assay kit and Qubit
TM 3.0 fluorometer (ThermoFisher Scientific). To normalize for concentration, all samples were diluted to the lowest measured concentration. The V4 region of the 16S rRNA gene was amplified as previously described [
29], with few modifications. Briefly, 10 µL of the template DNA, 5 µL Phusion HF Buffer (5X), 0.5 µL dNTPs mix (10 nM), 0.25 µL Phusion HF DNA polymerase (2 U/µL), 2.5 µL forward primer (5 µM), 2.5 µL reverse primer (5 µM) and 4.25 µL nuclease free water were used at a final 25 µL reaction volume per sample. Both the Phusion HF Buffer (5X) and Phusion HF DNA polymerase were obtained from New England BioLabs
® Inc. (Ipswich, MA, USA). For PCR amplification of the V4 region, unique barcoded primers 515F (5′-AATGATACGGCGACCACCGAGATCTACAC NNNNNNNN TATGGTAATT GT
GTGCCAGCMGCCGCGGTAA-3′) and 806R (5′-CAAGCAGAAGACGGCATACGAGAT NNNNNNNN AGTCAGTCAG CC
GGACTACH VGGGTWTCTAAT-3′) were used. The following PCR program was used; initial denaturation (98 °C, 30 s), 24 cycles of denaturation (98 °C, 10 s), annealing (55 °C, 30 s) and extension (72 °C, 30 s), and final extension (72 °C, 5 min). The PCR products were purified using AMPure XP beads (Illumina, San Diego, CA, USA), following the manufacturer’s instructions. The purification and correct amplicon (385 bp) were confirmed by electrophoresis on a 1% agarose gel in tris-base acetic acid EDTA (TAE) buffer. Equimolar (8 nM) amplicons at a final volume 10 µL for each sample were then pooled into a sterile 1.5 mL microcentrifuge tube (SARSTEDT), in Illumina resuspension buffer. The pooled samples were denatured and diluted to 1.5 pM. Finally, 350 µL of the 16S library (1.5 pM), 150 µL genomic library prepared using the Illumina DNA prep (20060060) and 15 µL of PhiX (1.5 pM) were combined. This combination was loaded on the MiniSeq Mid output (300 cycles) reagent cartridge (Illumina) and custom primers were added to the reagent cartridge as follows; 16.5 µL of 10 µM Read1.515F (5′-TATGGTAATTGTGTGCCAGCMGCCGCGGTAA-3′) was added to position 24; 18.3 µL of 10 µM Read2.806R (5′-AGTCAGTCAGCCGGACTACHVGGGTWTCTAAT-3′) added to position 25; 24.6 µL of 10 µM Index1.806R (5′-ATTAGAWACCCBDGTAGTCCGGCTGACTGACT-3′) and 25.3 µL of 10 µM Index2.515F (5′-TTACCGCGGCKGCTGGCACACAATTACCATA-3′) were added to position 28. During sequencing, paired-end reads (2 × 150 bp) were generated. The sequencing was performed at the Laboratory of Gene Technology (KU Leuven, Belgium).
2.10. Data Analysis
The demultiplexed files acquired from the sequencing platform were processed using the LotuS pipeline (version 1.62.1) [
30], with default parameters. In brief, the pipeline was used for reads1 and reads2 assembly, sequences quality checking, clustering, operational taxonomic units (OTUs) generation, and taxa assignments by Basic Local Alignment Search Tool (BLAST) against the SILVA database (version SILVA 138.1 SSU). The R package phyloseq (version 1.38.0) was used to import and organize the microbiota data. The aggregate rare function from microbiota package (version 1.16.0) was used to combine rare taxa into “others” category. The packages ranacapa (version 0.1.0) and ggplot2 (version 3.3.6) were used to generate the rarefaction curve. The R packages microbiota (version 1.16.0), ggplot2 (version 3.3.6) and hrbrthemes (version 0.8.0) were used for plotting the taxa abundances at genus and phylum levels. The microbiota R package was also used to plot alpha diversity while phyloseq R package was used to plot beta diversity. The adonis2 function from vegan package (version 2.5-7) was used to test if the control and the treatment groups were significantly different from each other via permutational multivariate analysis of variance (PERMANOVA), using distance matrices. All R-based analyses were performed in R software version 4.2.1 [
31].
The qPCR data were analyzed using the ΔΔCt (fold change) method on Microsoft Excel (version Microsoft 365) to compute the cecal immune-related genes expression. The data were then further analyzed using the GraphPad Prism 9.0.0 (San Diego, CA, USA) software for statistical testing in different treatment groups, in which two-way ANOVA was used. GraphPad Prism was also used to compare the cecal microbiotas taxa abundances, and antibodies level in different treatment groups. For these purposes, the Kruskal–Wallis test was used and p values ≤ 0.05 were considered significant. Data are presented as mean and standard error of the mean (mean ± S.E.M).
4. Discussion
The ban on antibiotics use in many parts of the world due to the increasing problem of antibiotic resistance has resulted in an urgent search for non-antimicrobial alternatives. Various alternatives have been developed. These include vaccines, which are widely applied in the poultry industry to limit
Salmonella contamination at the farm level. Nevertheless, the mechanisms of action for most antimicrobial alternatives are yet to be fully understood [
10,
11], especially in the host–microbiota–pathogen context. Both inactivated and live
Salmonella vaccines are commercially available and used in poultry farms, but live vaccines are considered more protective due to their ability to induce both cellular and humoral immune responses. Live attenuated
Salmonella vaccines are mostly orally administered in poultry, exposing them to a complex environment comprising the host immune factors and commensal gut microbiota. Being live bacteria, it is expected that the vaccine strains will need to colonize the gut and establish a niche, although for a limited period of time. The effect that live attenuated
Salmonella vaccine has on the poultry gut microbiota is not so clear. In this study, we used three commercial live attenuated
Salmonella vaccine strains for
S. Typhimurium (AviPro
® Salmonella Vac T),
S. Enteritidis (AviPro
® Salmonella Vac E) and
S. Typhimurium +
S. Enteritidis (AviPro
® Salmonella DUO) to study their influence on chicken cecal microbiota profiles. Chicks were orally exposed to the attenuated vaccines and ceca, cecal contents, and sera sampled 14 days post-vaccination for analysis of cecal immune related gene expression,
Salmonella-specific antibodies (IgG and IgA), and microbiota population structure.
The effect of live
Salmonella vaccines on cecal immune genes was studied by comparing the expression of the cytokines involved in type 1 (IL-18, TNFα), type 2 (IL-4, IL-10, IL-6) and type 3 (IL-17A, IL-8) immune responses. The avian β-defensin (AvBD1), a cationic antimicrobial peptide secreted by gut epithelial cells as an effector signature of IL-17 was also analyzed. Forkhead Box P3 (FoxP3) and inducible nitric oxide synthase (iNOS) were also included to study the regulatory T-cells (Tregs) levels (indicative of the non-inflammatory, tolerogenic state, important for persistent
Salmonella colonization) and oxidative burst status (iNOS), respectively. In chickens, FoxP3, is a recently discovered member of the FOX proteins family and acts as a pivotal regulator in the regulatory pathway for biosynthesis and for the functioning of Treg cells [
24].
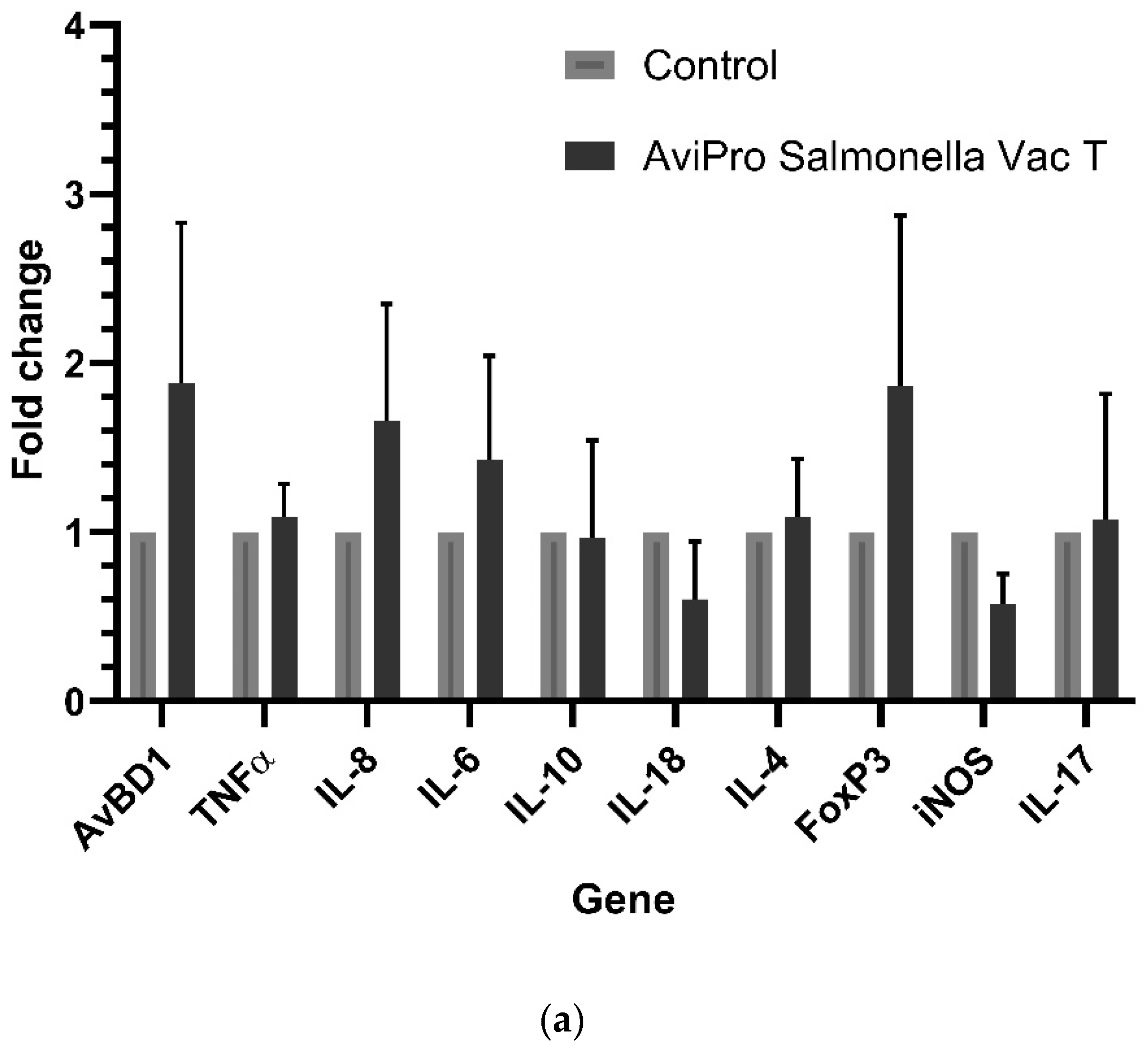
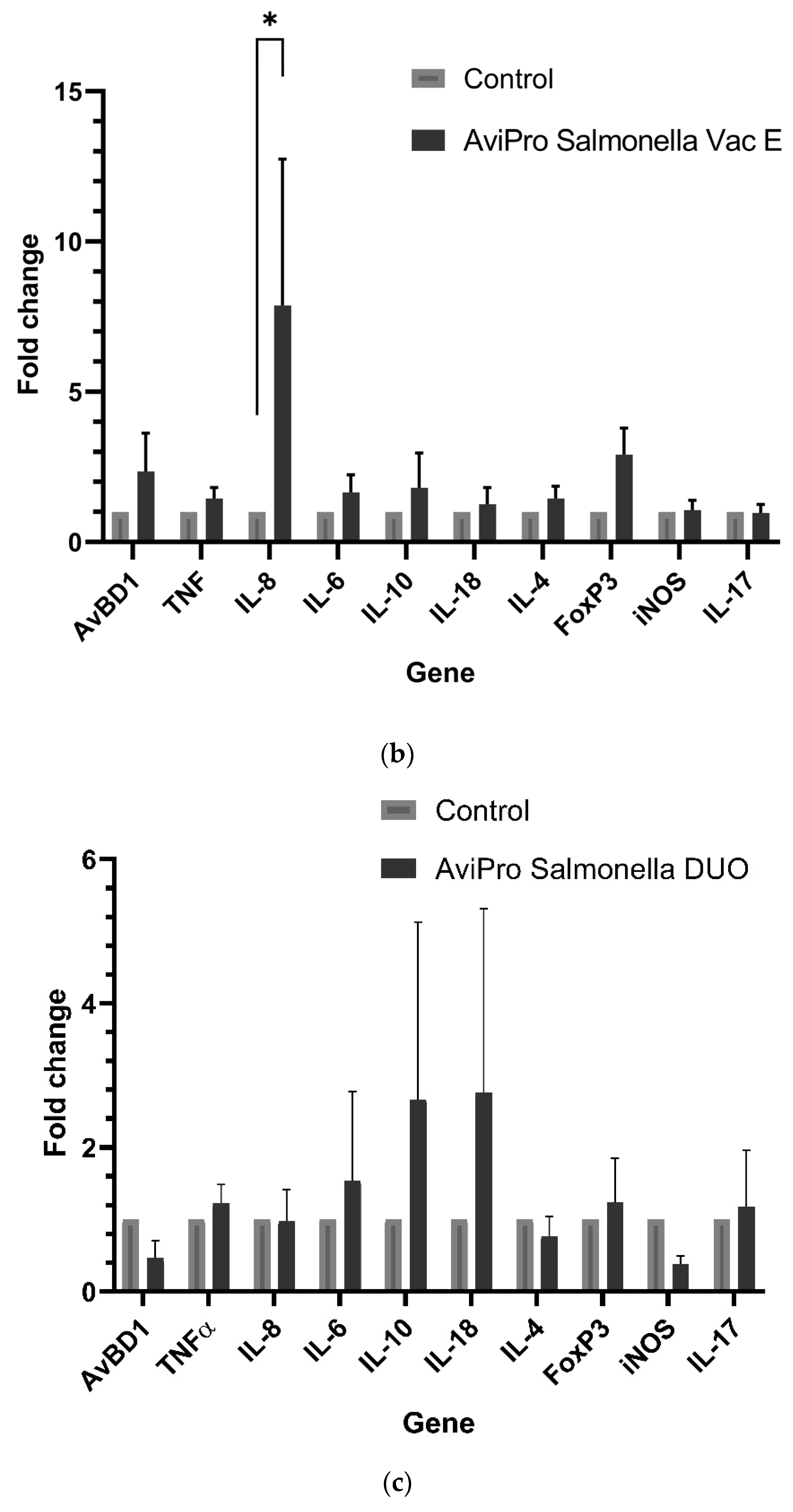
Generally, it is expected that invasive live
Salmonella vaccines should induce similar immune responses as their pathogenic counterparts. Our data show a significantly high IL-8 expression in the AviPro
® Salmonella Vac E vaccinated chickens (
Figure 1b). Following this cytokine’s role in the recruitment of heterophils to the inflammatory site, it could be suggested that this vaccine induced higher inflammation compared with the other two, although this might need further confirmation of heterophils infiltration into the cecal tissues. Furthermore, an oral immunization with attenuated
Salmonella vaccine strains during this study triggered an upregulation of some cecal immune genes in broiler chickens, although the fold changes in their expression were not significant. This was the case for the AviPro
® Salmonella Vac E (
Figure 1b), but not for the AviPro
® Salmonella Vac T (
Figure 1a) or the AviPro
® Salmonella DUO (
Figure 1c) in which iNOS and IL-18, and IL-4, iNOS and AvBD1 were downregulated. This is partly in agreement with the findings by Carvajal et al. [
32] and Pan et al. [
33], in which IL-8 (CXCLi2), IFNγ, IL-6, and IL-2 expression were induced in ceca following an oral immunization of chickens with an attenuated recombinant
Salmonella enterica serovar Typhimurium vaccine candidate. Additionally, a study by Kogut et al. (2016) reported highly induced IL-6 expression on day 1, followed by a decrease until day 14 post infection with
Salmonella Enteritidis in chicken ceca. The IL-10 expression, on the other hand, increased towards day 14 post infection [
3]. Although our study was not time-point designed as was the case for Kogut et al., we also observed (
Figure 1b,c) similar trends of IL-10 and IL-6 expression at sampling (14 days since vaccination).
The expression of IL-17A (
Figure 1a–c) was not induced above the control level following an oral exposure to all the studied live
Salmonella vaccines. Congruent to our observation, an oral immunization with the SPI-1 mutant
S. Enteritidis triggered low expression of IL-17 and IL-22 in chicken ceca [
34]. It has been proposed that the interaction between
Salmonella and the chicken gut triggers immune responses that can be divided into three stages, depending on the duration of interaction since exposure. The stages are associated with changes not only of the inflammatory status but also metabolic reprogramming towards less responsiveness against the pathogen [
2]. Considering the sample collection timing post immunization and our cecal immune related gene expression data, it can be suggested that the anti-inflammatory (stage 2) and homeostatic (stage 3) responses were mostly dominant, as also indicated by the IL-10, FoxP3, iNOS, IL-4, IL-17A expression profiles.
Taken together, our data suggest that the live attenuated Salmonella vaccine AviPro® Salmonella DUO induced a more anti-inflammatory cecal environment (IL-10) and Th1 responses (IL-18) both of which are crucial for limiting the contamination level with Salmonella in chickens. Furthermore, the tolerogenic environment (FoxP3 expression), necessary for persistent colonization of chickens with Salmonella was lower in the AviPro® Salmonella DUO vaccinated chickens, which might be indicative of improved protection.
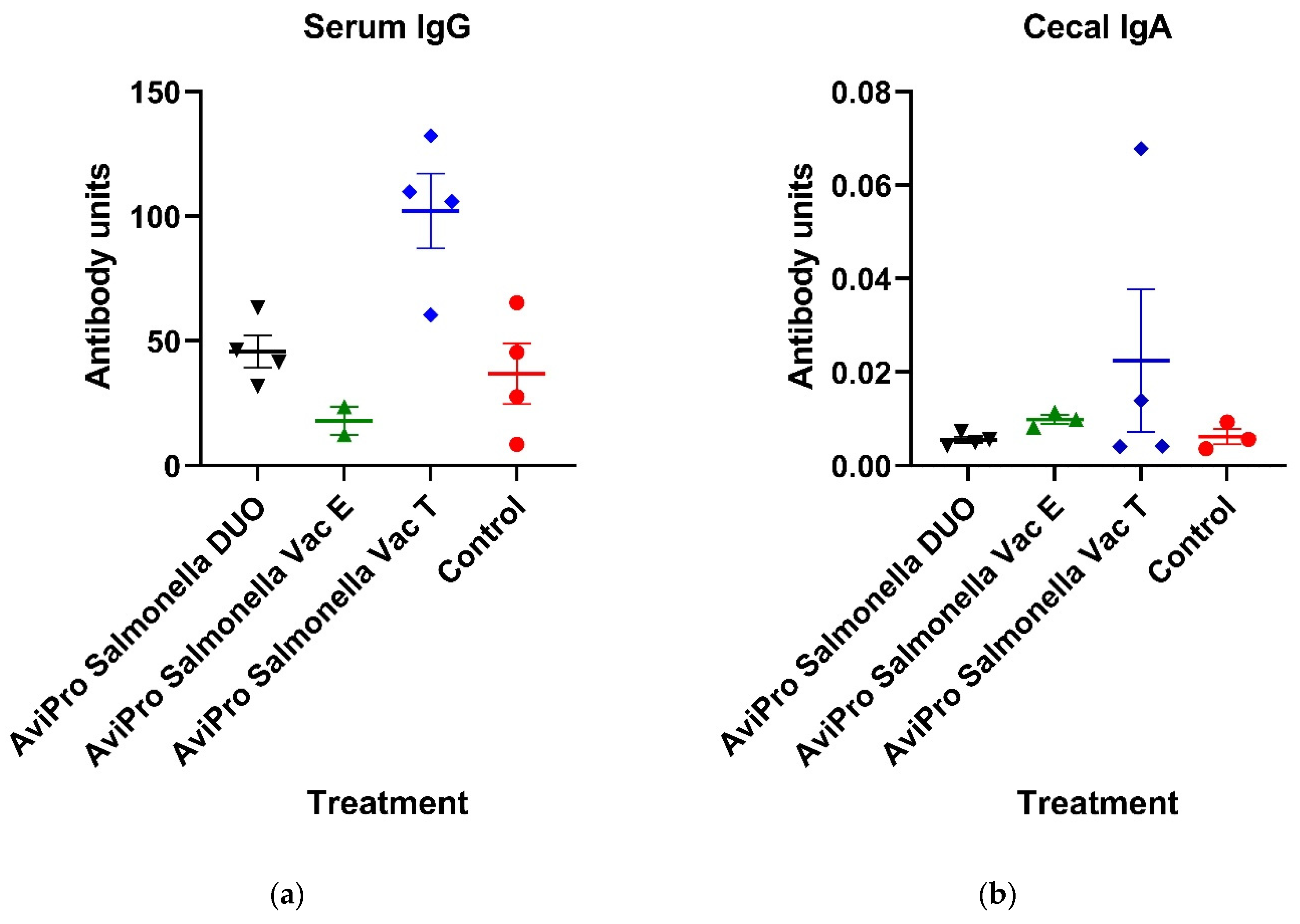
To study the vaccine-induced humoral responses, whole
Salmonella-specific antibodies were quantified and compared in vaccinated and unvaccinated chickens. Specifically, IgG and IgA, which are the most dominant chicken immunoglobulins in serum and gut mucosa, respectively, were measured by ELISA. Although not significantly, our data show that the AviPro
® Salmonella Vac T induced an increase in serum IgG above the control level (
Figure 2a). On the other hand, the cecal IgA level in all vaccinated chickens were generally low and not different from unvaccinated controls (
Figure 2a). This is in contrast with the findings by Matsui et al. [
35], in which oral administration of a live virulence plasmid-cured
S. Typhimurium vaccine candidate in mice induced high anti-
S. Typhimurium IgA and IgG levels in serum, cecal homogenates, intestinal and lung lavage fluids and in bile as compared with unvaccinated mice. Moreover, higher antibodies were observed following three oral vaccination rounds than following two rounds or one. Similar findings were also reported by Bridge et al. [
36] in their study using mice orally or intraperitoneally vaccinated with a live
Salmonella vaccine candidate. Immunization increased
Salmonella-specific IgG and sIgA as compared with the unvaccinated controls, 25–27 days post vaccination. Furthermore, similar trends of IgA and IgG in serum and fecal pellets of mice [
37] or swine [
38] immunized with live attenuated
Salmonella vaccines have been reported. The low and non-significant IgG and IgA levels in vaccinated chickens observed in our study could partly be contributed by a single oral administration of the live
Salmonella vaccines prior to sampling. Nevertheless, the single oral vaccination round performed in this study was according to the manufacturers’ recommendation for broiler chickens. It is also specified by the manufacturer that low seropositivity reactivity of individual birds in a flock are possible following oral vaccination with the AviPro
® Salmonella Vac T, AviPro
® Salmonella Vac T, and AviPro
® Salmonella DUO (AviPro
® Salmonella DUO user manual, 2020) [
39], which could be another possible explanation for our antibody ELISA data.
Next, the live
Salmonella vaccine’s effect on chicken cecal microbiota was investigated. The chicken gut is inhabited by millions of commensal microbiota which vary in composition and abundances with gut section, age of the bird, feed, water, breed, environmental factors (hygiene or biosecurity levels), and the host immune factors. The highest abundance is found in ceca, the two blind pouches located at the posterior end of the chicken gut, consisting of up to 10
10–10
11/g cecal content [
40]. Microbiota are known to play various roles in the chicken gut, which determine the bird’s health via symbiotic interactions. These include the digestion of non-fermentable fibers, making them available for uptake by the bird; the inducing of gut development [
40]; and the promotion of the development, maturation, training and functioning of chicken immunity [
41,
42].
There are conflicting data on the effect of
Salmonella infection on gut microbiota. A study by Mon et al. [
43] showed that
S. Enteritidis induced a reduction in microbiota diversity in chicken, with members of the
Enterobacteriaceae (Proteobacteria) family being dominant while the
Lachnospiraceae family (particularly butyrate producers) and
Ruminococcus were significantly reduced and negatively correlated with the
Enterobacteriaceae family expansion due to competitive interaction between the two taxa, 7 days post infection. Early (2 days) post infection, Proteobacteria phylum expansion resulted in Firmicutes reduction. Similarly, a study by Khan and Chousalkar [
44] reported reduced abundances in the genera Lactobacillus,
Faecalibacterium, Bifidobacterium,
Alistipes and Butyricimonas following an infection with virulent
S. Typhimurium while increasing the Butyricicoccus, Oscillibacter and Eryscipelatoclostridium genera abundances.
On the other hand, Videnska et al. [
45] reported a minor effect on microbiota alteration upon
Salmonella infection in chickens.
Enterobacteriaceae and
Ruminococcaceae were shown to both increase in
S. Enteritidis-infected chickens as compared with uninfected chickens, although this was not statistically significant. Likewise, the study by Zeng et al. [
46] showed that at day 0 post infection with
S. Enteritidis, the family
Enterobacteriaceae dominated the chicken ceca. Ten days later, the microbiota were more diverse and stabilized, consisting of
Ruminococcaceae (Firmicutes),
Enterobacteriaceae (Proteobacteria),
Lachnospiraceae (Firmicutes) and
Clostridiaceae (Firmicutes). Overall,
S. Enteritidis infection in 2-day old chickens did not significantly alter the cecal microbiota structure.
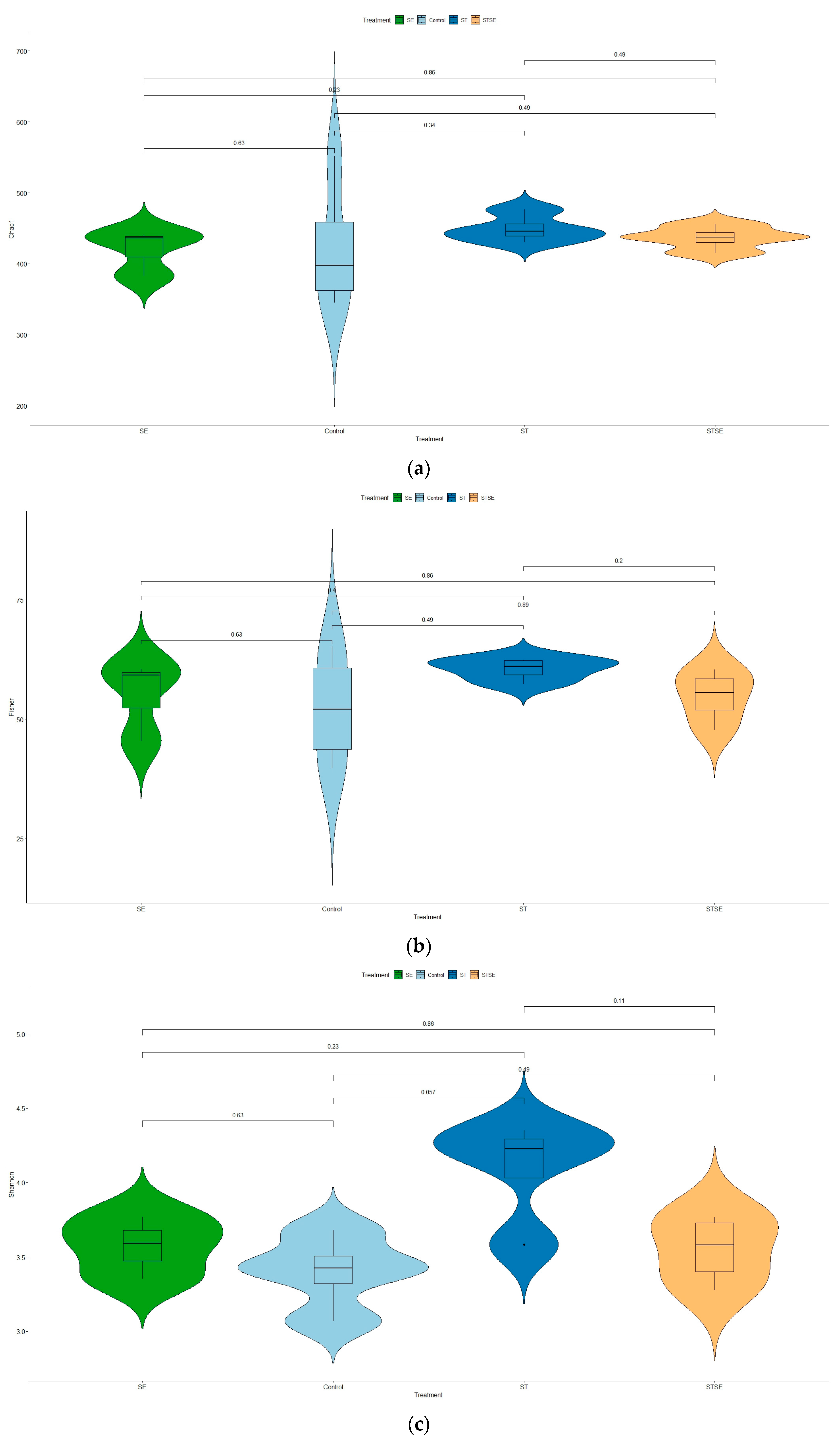
Looking into the influence of live attenuated
Salmonella vaccines on cecal microbiotas, our data show that variability within samples (α-diversity measure) was only marginally significant in the AviPro
® Salmonella Vac T vaccinated chickens (
Figure 4c). The other two vaccines (AviPro
® Salmonella DUO and AviPro
® Salmonella Vac E), however, did not indicate a significant influence on cecal microbiota variability (
Figure 4a,b).
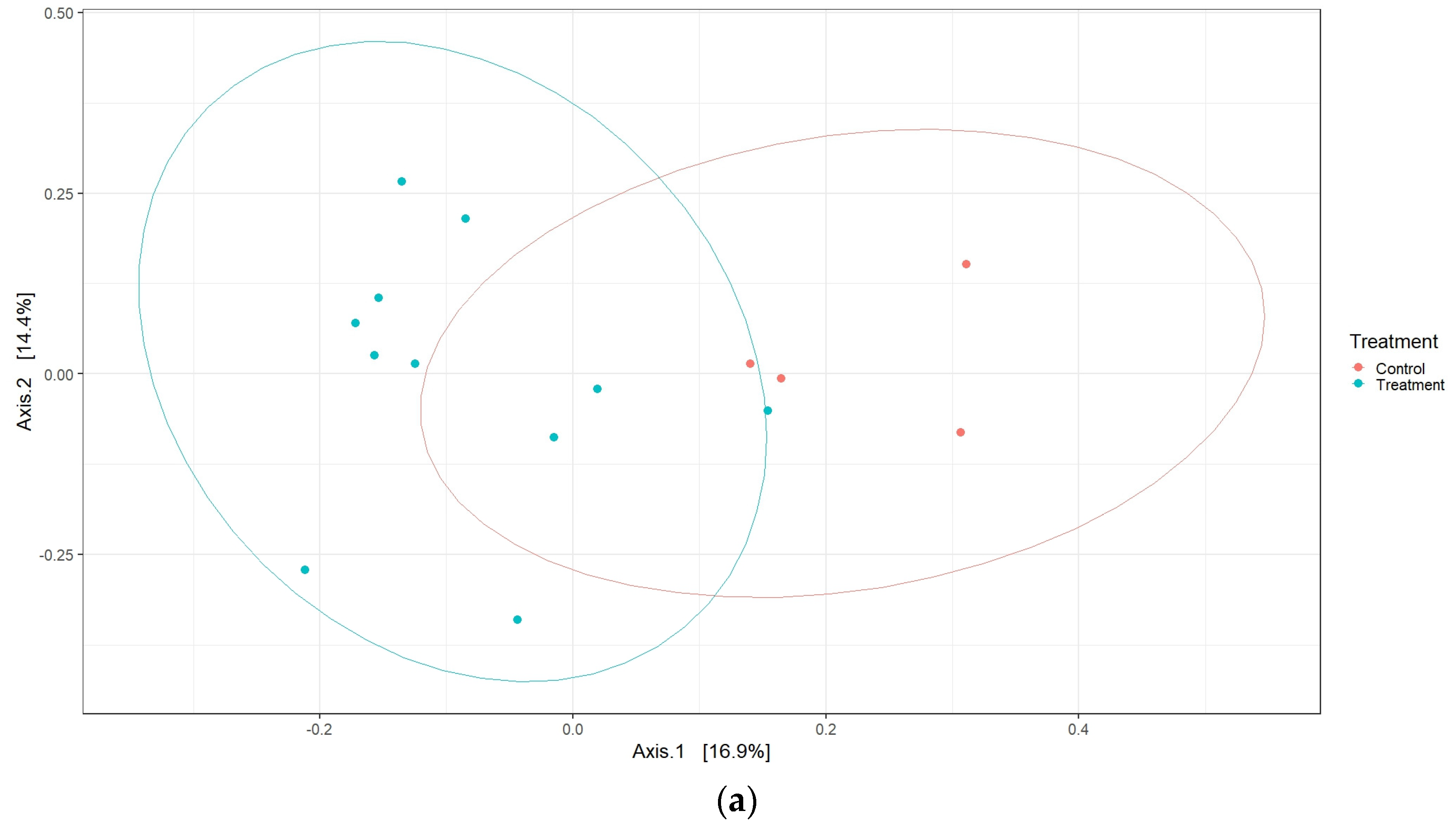
Although there were no differences in alpha diversity, interesting effects were observed for the β-diversity. Data from this study show that live vaccines in general had a significant influence on the cecal microbiota shaping, as suggested by the different clustering between vaccine and control chicken samples (
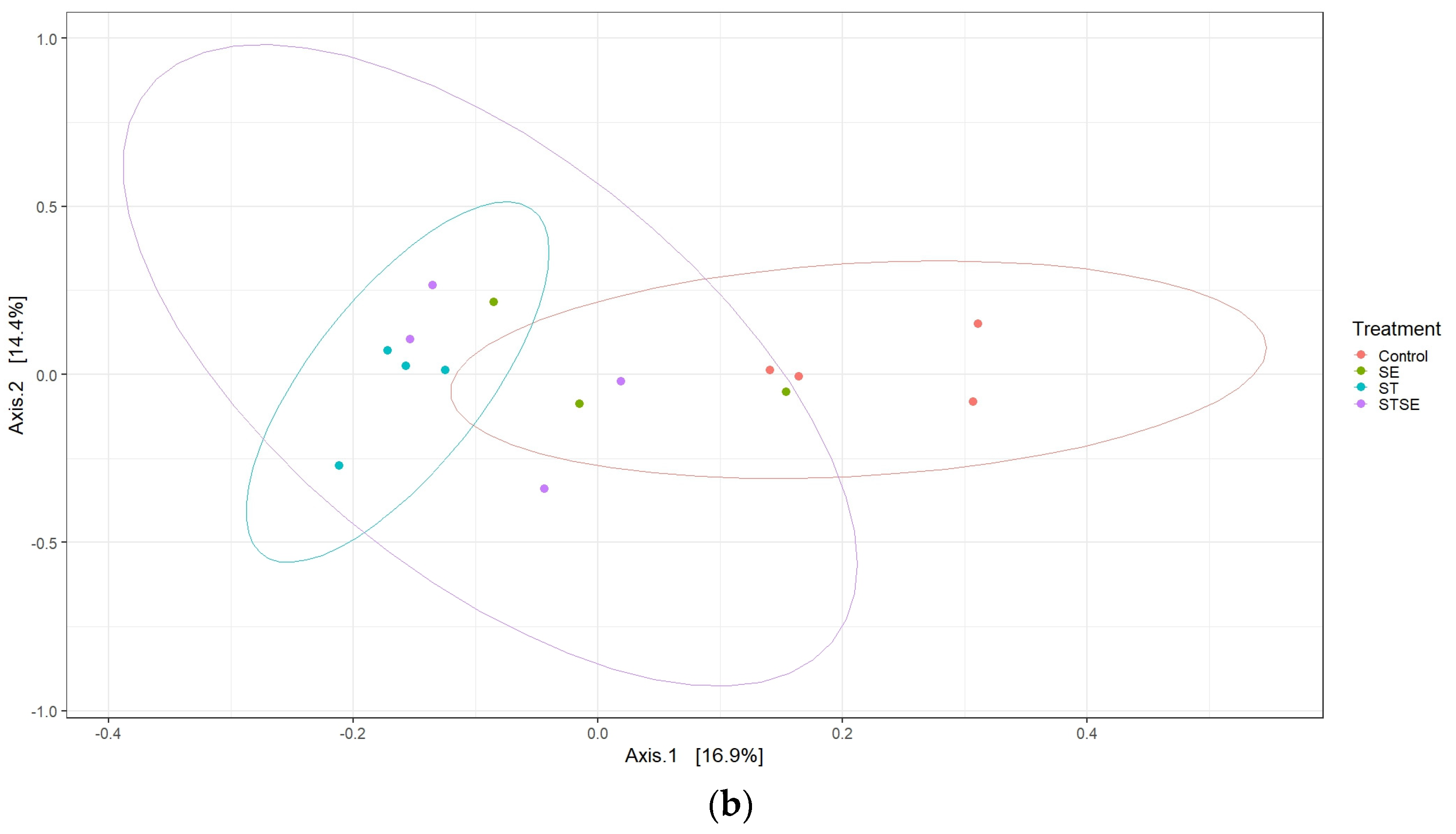
Figure 5a). Next, we indicated that, individually, the AviPro
® Salmonella Vac T and AviPro
® Salmonella DUO, but not the AviPro
® Salmonella Vac E vaccine, had significant effects on the chicken cecal microbiota variability (
Figure 5b).
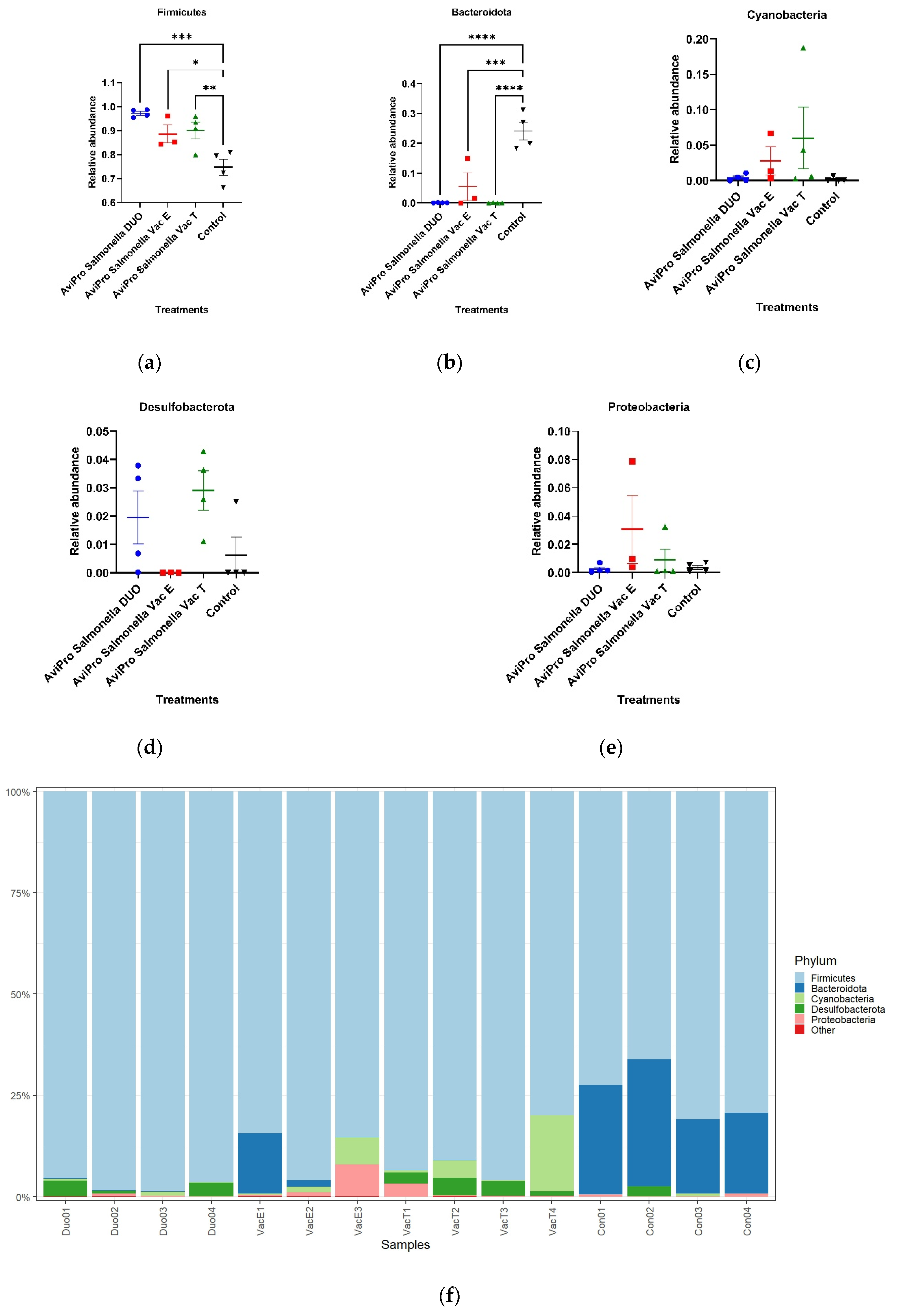
At the phylum level, the current study revealed high Firmicutes abundance in broiler chickens irrespective of their vaccination status with the live attenuated
Salmonella vaccines. The dominance of Firmicutes in broilers ceca observed here has also been reported by Oakley et al. [
21], and that this was almost an exclusive phylum in broiler chickens older than a week. Similar results by Qi et al. [
47], also showed that Firmicutes and Bacteroidota were the most dominant broiler cecal taxa at the phylum level. Nevertheless, our study showed that the live attenuated
Salmonella vaccines significantly increased the Firmicutes relative abundances as compared with unvaccinated chickens (
Figure 6). Contrary to our data, Park et al. [
17] reported the highest abundance for Firmicutes in unvaccinated broiler chickens and the oral challenge with live attenuated
Salmonella Typhimurium vaccine candidates (PBAD-mviN
S. Typhimurium UK-1 and ΔΔ
metRmetD S. Typhimurium UK-1) did not significantly alter this trend. A decrease in Firmicutes abundances has also been reported in layer chickens [
48]. Unlike our study, however, the findings by Joat and colleagues followed an intramuscular injection with a wild-type
Salmonella Typhimurium UK-1 strain.
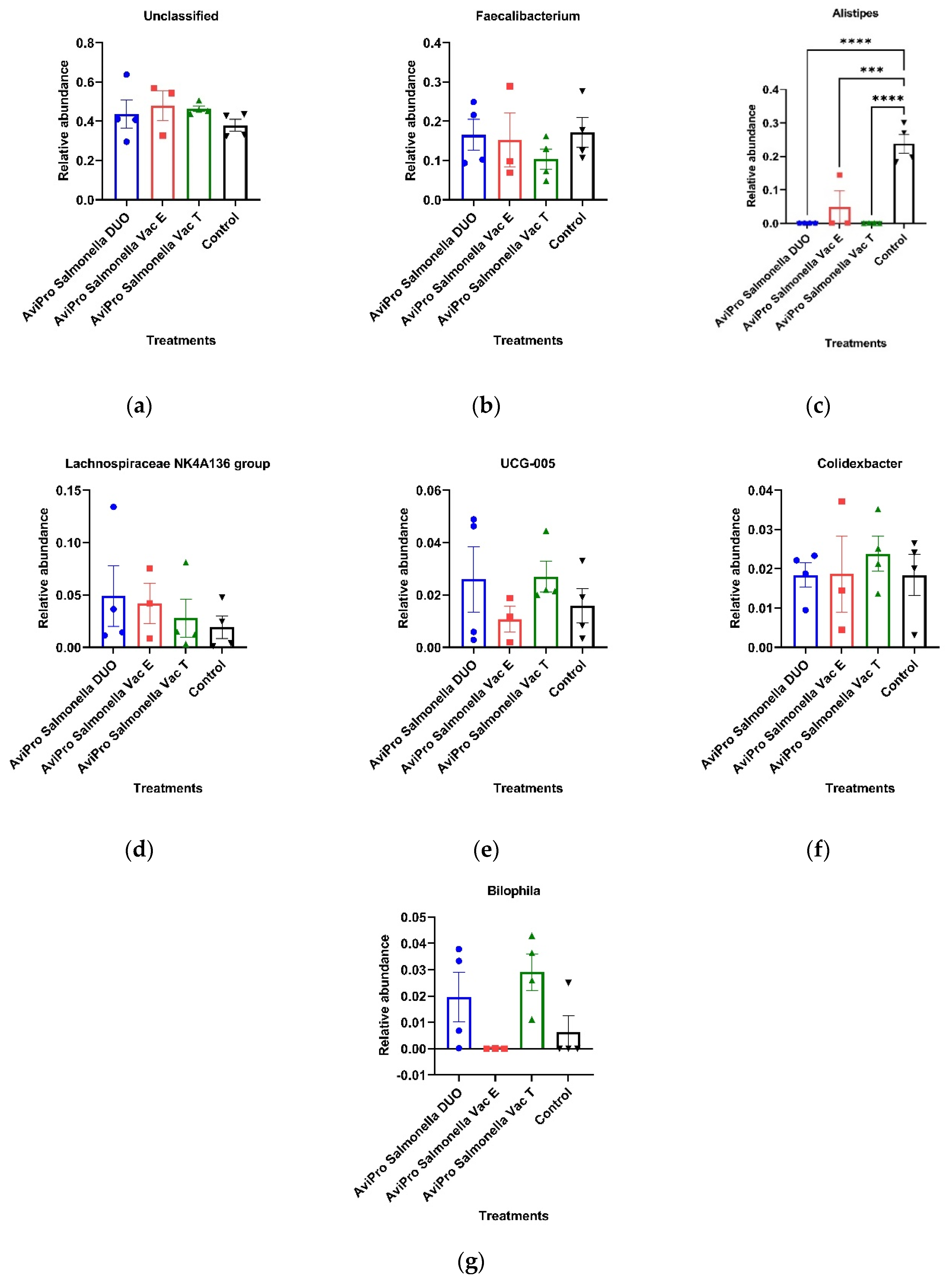
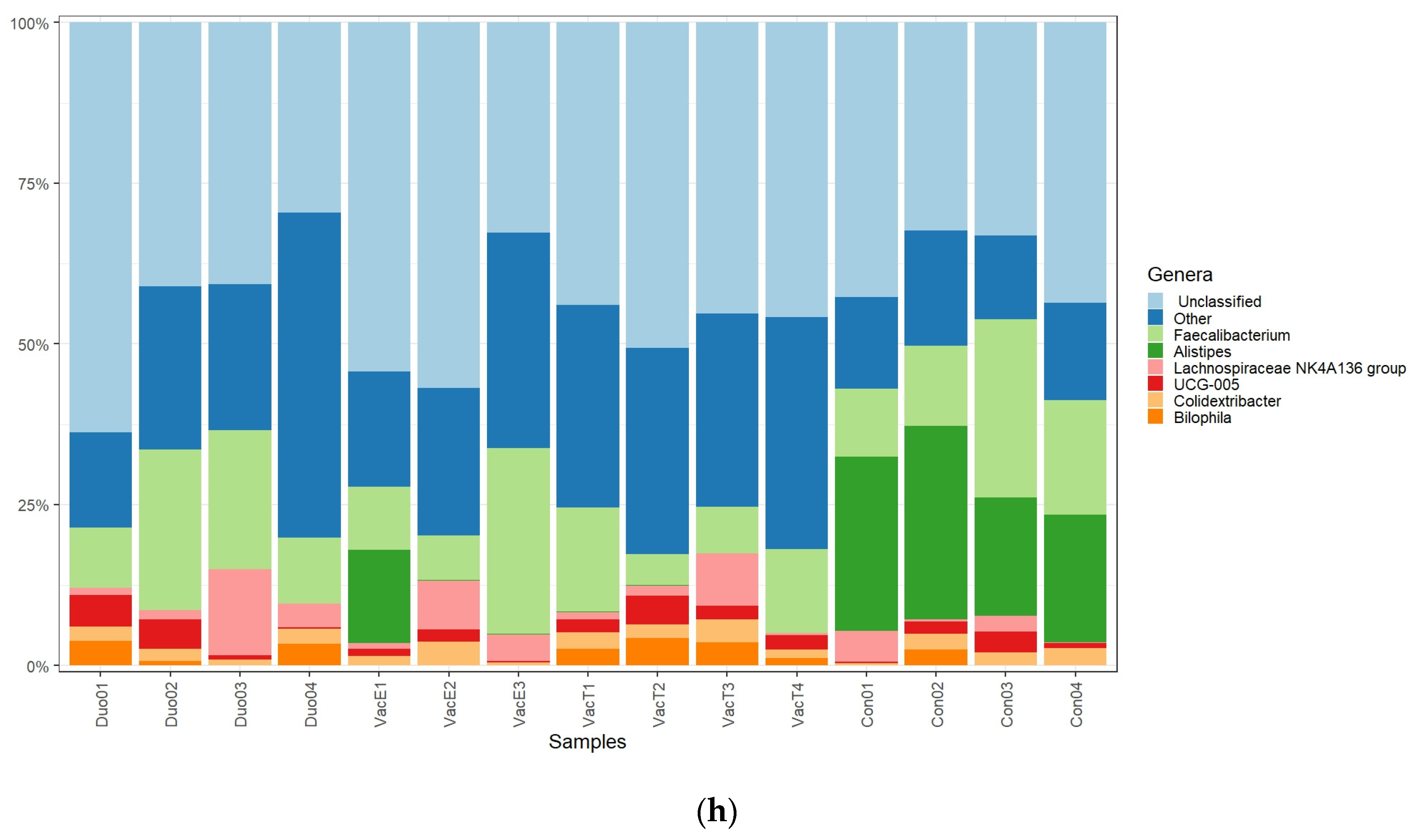
Consistent with our observations at phylum level, most high abundant genera identified were also members of the Firmicutes. The genus
Faecalibacterium is a well-known abundant cecal genus of the phylum Firmicutes [
49] and with well-known commensal benefits to chickens. Some
Faecalibacterium members have been described to have anti-inflammatory properties in human and mice [
50]. As shown in
Figure 7b, our data suggest that the different live
Salmonella vaccines used in this study did not significantly influence this genus, which is in agreement with the previous report by Park et al. [
17].
Similarly, the genera
Colidextribacter, UCG-005, and the
Lachnospiraceae NK4A136 group, which also all belong to the phylum Firmicutes showed a common distribution among the vaccinated and unvaccinated chickens. Additionally, the relative abundances for these genera were not significantly different among the treatment groups (
Figure 7d–f). The findings by Park et al. [
17], contrary to ours, however, showed an increase in
Lachnospiraceae abundances in broiler chickens challenged with a PBAD-mviN
S. Typhimurium UK-1 vaccine candidate as compared with the unvaccinated. This could be contributed by the difference in age at sampling (16 days) in the current study, as compared with six weeks in the study by Park and colleagues. Generally, the enhanced broiler cecal Firmicutes abundances by live
Salmonella vaccines can be considered an improvement in protection and health following the phylum members’ roles in protecting the host against pathogens and complex carbohydrates degradation [
50].
The phylum Bacteroidota, was more represented in the unvaccinated than in the vaccinated chickens (
Figure 6f). Our data, thus, suggest that the live vaccines used significantly reduced the abundance of this phylum (
Figure 6b). Some Bacteroidota genera, such as
Alistipes, are known to be short chain fatty acid (SCFA) producers from indigestible fiber fermentation and hence have an anti-inflammatory role in the animal gut and can considered beneficial to the host [
51,
52]. A similar trend was also observed at the genus level, in which
Alistipes was mostly common in the unvaccinated chickens for which it appeared at the highest abundance compared with the vaccinated groups (
Figure 7). In line with our data, Orso et al. [
53] reported a decrease in Bacteroidetes and SCFAs producing genera abundances following a live coccidiosis vaccine in chickens as compared with the unvaccinated control.
The present study also identified Cyanobacteria, Proteobacteria and Desulfobacterota among the abundant phyla. Congruent to our data, Cyanobacteria and Proteobacteria have also been reported by Orso et al. [
53], as next to Firmicutes and Bacteroidetes in abundances for broiler chickens. Additionally, in their work, an increased Proteobacteria abundance following a live vaccine against coccidiosis is reported. Proteobacteria are mostly associated with the generation of an inflammatory environment in the gut leading to dysbiosis, following their fermentative metabolism which mostly favors pathogenic bacteria [
51] such as
Salmonella. This is contradictory to the vaccination purpose in
Salmonella (a Proteobacteria member) control programs. Our findings are in agreement with those of Orso et al. (2021), but the high abundances of Proteobacteria in the vaccinated chickens observed in the current study (although not significant) could be explained by the live attenuated
Salmonella vaccines in the ceca, as the sampling was performed within the vaccines shedding window (up to 28 days post vaccination).
Our data are partly in agreement with the findings of Park et al. [
17] in which the live
Salmonella Typhimurium vaccine candidates affected only the presence of some cecal microbiota without influencing their overall relative abundances. However, here, we also observed that some taxa relative abundances were significantly altered with the vaccination status, and this was the case for all the vaccines used in this study (
Figure 6 and
Figure 7).













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}