
BBIBP-CorV Vaccination against the SARS-CoV-2 Virus Affects the Gut Microbiome

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Fecal Sample Collection and DNA Preparation
2.3. PCR Amplicon and Sequencing of 16S rRNA Gene
2.4. Bioinformatics and Statistical Analysis
3. Results
3.1. Clinical Characteristics of the Recruited Subjects
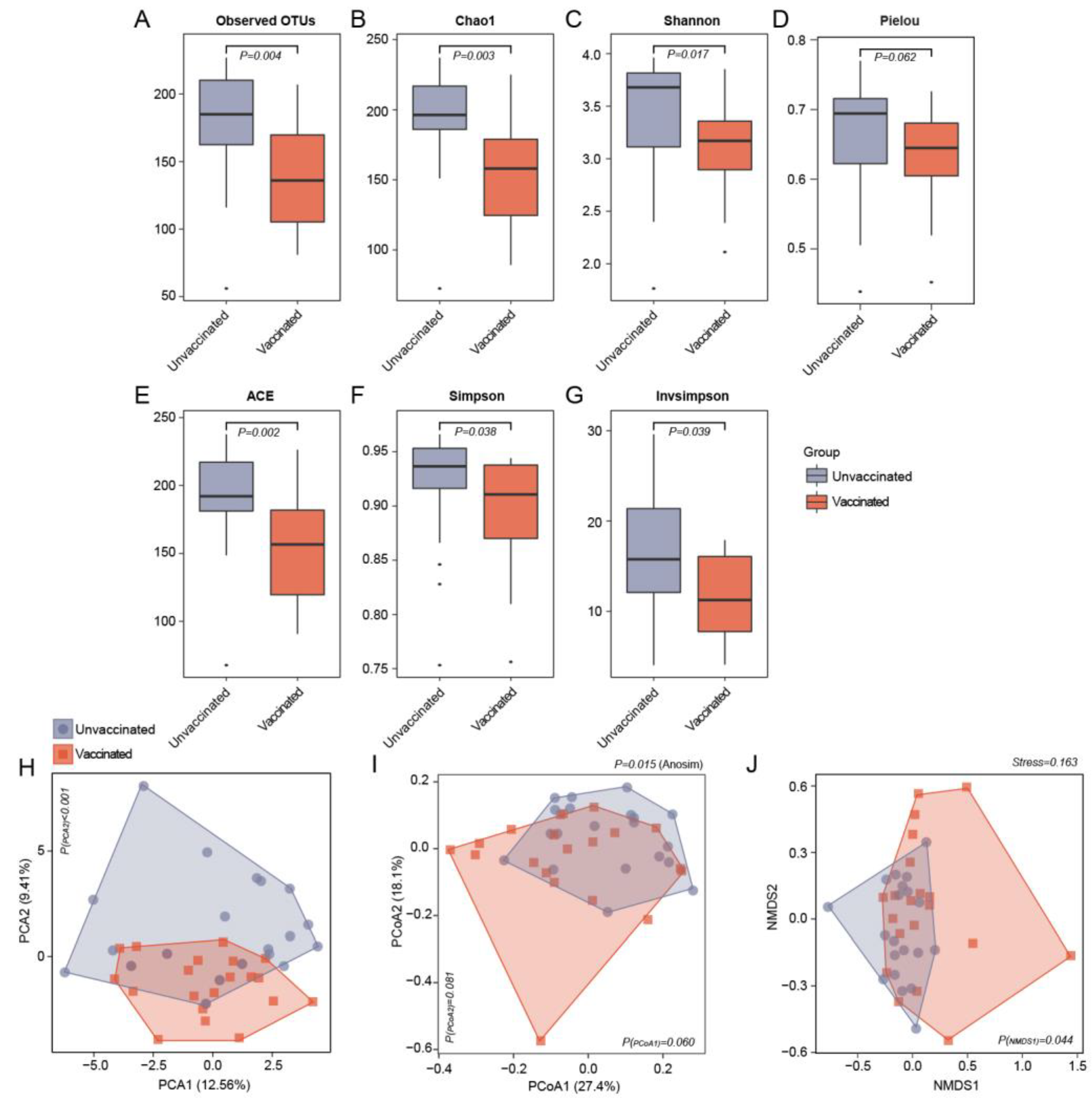
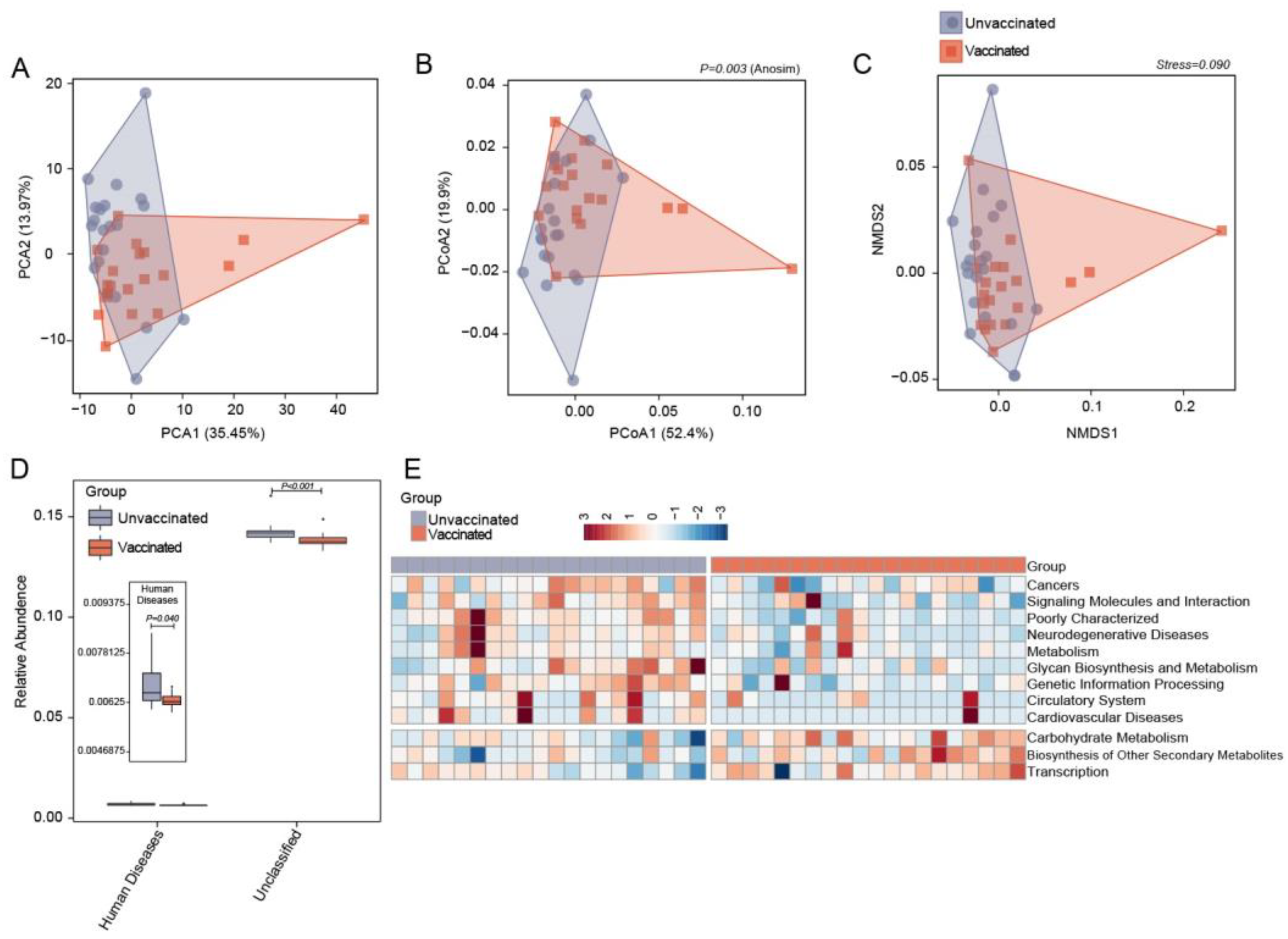
3.2. Differences in Gut Microbiome Community Structure in Unvaccinated and Vaccinated Subjects
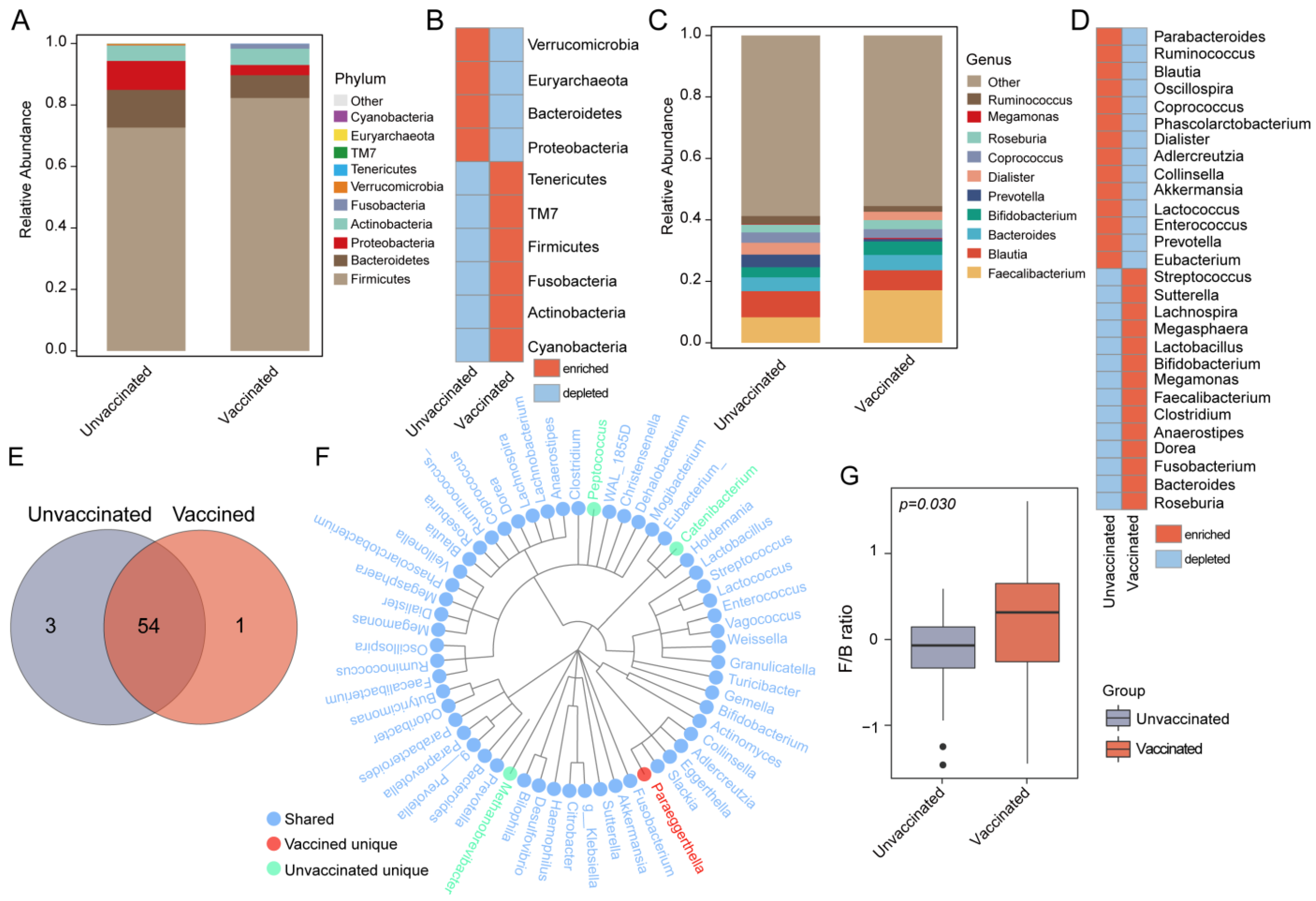
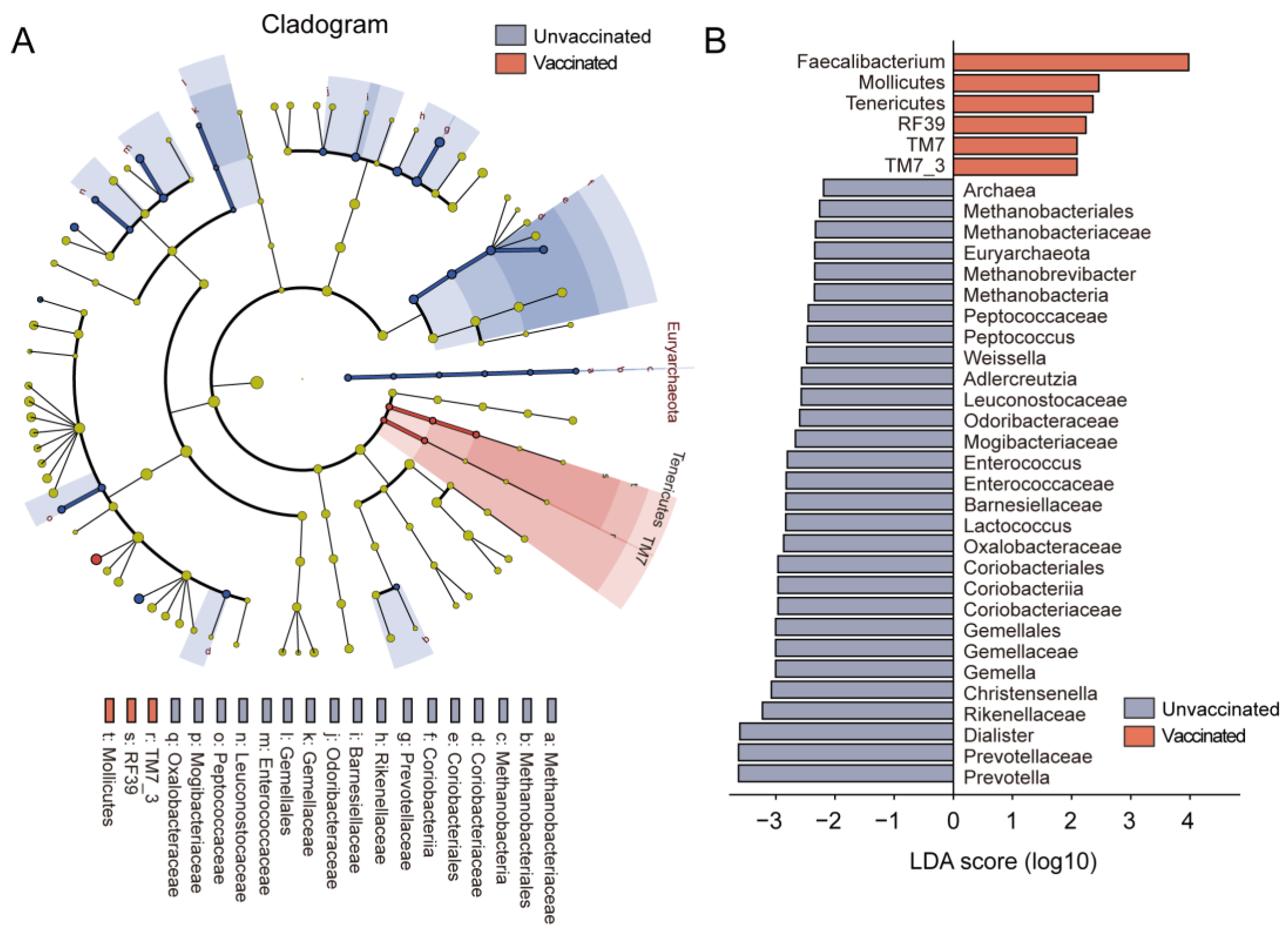
3.3. Gut Features and Comparison of Microbial Profiles in Taxa
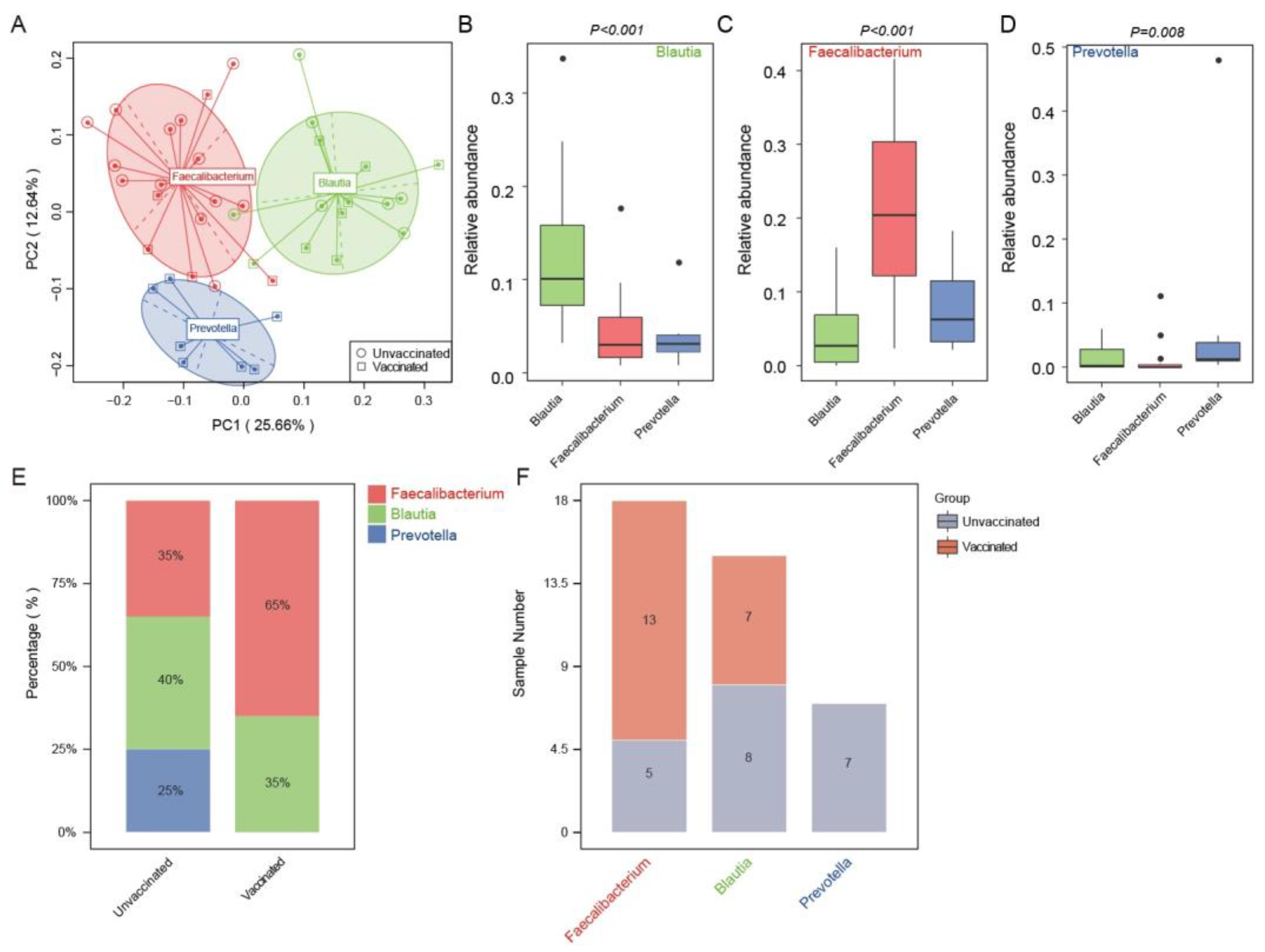
3.4. Enterotype Distribution Indicated an Inclination for Faecalibacterium-Dominated Types upon Vaccination
3.5. Predicting Functional Capacities of Gut Microbiota Specific to Vaccinated Status via PICRUST Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, C.; Horby, P.W.; Hayden, F.G.; Gao, G.F. A novel coronavirus outbreak of global health concern. Lancet 2020, 395, 470–473. [Google Scholar] [CrossRef] [PubMed]
- COVID-19 Coronavirus Pamdemic. 2023. Available online: https://www.worldometers.info/coronavirus/ (accessed on 28 April 2023).
- Xiao, F.; Tang, M.; Zheng, X.; Liu, Y.; Li, X.; Shan, H. Evidence for Gastrointestinal Infection of SARS-CoV-2. Gastroenterology 2020, 158, 1831–1833. [Google Scholar] [CrossRef] [PubMed]
- Lamers, M.M.; Beumer, J.; Van Der Vaart, J.; Knoops, K.; Puschhof, J.; Breugem, T.I.; Ravelli, R.B.; Paul van Schayck, J.; Mykytyn, A.Z.; Duimel, H.Q.; et al. SARS-CoV-2 productively infects human gut enterocytes. Science 2020, 369, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Wölfel, R.; Corman, V.M.; Guggemos, W.; Seilmaier, M.; Zange, S.; Müller, M.A.; Niemeyer, D.; Jones, T.C.; Vollmar, P.; Rothe, C.; et al. Virological assessment of hospitalized patients with COVID-2019. Nature 2020, 581, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, X.; Zhu, B.; Liang, H.; Fang, C.; Gong, Y.; Guo, Q.; Sun, X.; Zhao, D.; Shen, J.; et al. Characteristics of pediatric SARS-CoV-2 infection and potential evidence for persistent fecal viral shedding. Nat. Med. 2020, 26, 502–505. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xu, Y.; Gao, R.; Lu, R.; Han, K.; Wu, G.; Tan, W. Detection of SARS-CoV-2 in Different Types of Clinical Specimens. JAMA 2020, 323, 1843–1844. [Google Scholar] [CrossRef]
- Schmidt, T.S.B.; Raes, J.; Bork, P. The Human Gut Microbiome: From Association to Modulation. Cell 2018, 172, 1198–1215. [Google Scholar] [CrossRef]
- Kelly, D.; Conway, S.; Aminov, R. Commensal gut bacteria: Mechanisms of immune modulation. Trends Immunol. 2005, 26, 326–333. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef]
- Ralli, T.; Saifi, Z.; Rathee, A.; Aeri, V.; Kohli, K. Decoding the bidirectional relationship between gut microbiota and COVID-19. Heliyon 2023, 9, e13801. [Google Scholar] [CrossRef]
- Chen, Y.; Gu, S.; Chen, Y.; Lu, H.; Shi, D.; Guo, J.; Wu, W.R.; Yang, Y.; Li, Y.; Xu, K.J.; et al. Six-month follow-up of gut microbiota richness in patients with COVID-19. Gut 2022, 71, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Yeoh, Y.K.; Zuo, T.; Lui, G.C.Y.; Zhang, F.; Liu, Q.; Li, A.Y.; Chung, A.C.; Cheung, C.P.; Tso, E.Y.; Fung, K.S.; et al. Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID-19. Gut 2021, 70, 698–706. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. COVID-19 Vaccine Tracker and Landscape. Available online: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 30 March 2023).
- Xia, S.; Zhang, Y.; Wang, Y.; Wang, H.; Yang, Y.; Gao, G.F.; Tan, W.; Wu, G.; Xu, M.; Lou, Z.; et al. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBIBP-CorV: A randomised, double-blind, placebo-controlled, phase 1/2 trial. Lancet Infect. Dis. 2021, 21, 39–51. [Google Scholar] [CrossRef]
- Nadeem, I.; Ul Munamm, S.A.; Ur Rasool, M.; Fatimah, M.; Abu Bakar, M.; Rana, Z.K.; Khatana, U.F.; Jordon, L.; Saqlain, M.; Mahdi, N.; et al. Safety and efficacy of Sinopharm vaccine (BBIBP-CorV) in elderly population of Faisalabad district of Pakistan. Postgrad. Med. J. 2022. [Google Scholar] [CrossRef]
- Castelli, J.M.; Rearte, A.; Olszevicki, S.; Voto, C.; Del Valle Juarez, M.; Pesce, M.; Iovane, A.N.; Paz, M.; Chaparro, M.E.; Buyayisqui, M.P.; et al. Effectiveness of mRNA-1273, BNT162b2, and BBIBP-CorV vaccines against infection and mortality in children in Argentina, during predominance of delta and omicron covid-19 variants: Test negative, case-control study. BMJ 2022, 379, e073070. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Tang, L.; He, W.; Jiang, X.; Hu, C.; Li, Y.; Zhang, Y.; Pang, K.; Lei, Y.; Li, S.; et al. Correlation of gut microbiota and metabolic functions with the antibody response to the BBIBP-CorV vaccine. Cell Rep. Med. 2022, 3, 100752. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Weng, S.; Wu, J.; Tian, X.; Zhang, Y.; Wang, X.; Wang, J.; Yan, D.; Wang, W.; Fang, F.; et al. Preexisting antibodies targeting SARS-CoV-2 S2 cross-react with commensal gut bacteria and impact COVID-19 vaccine induced immunity. Gut Microbes 2022, 14, 2117503. [Google Scholar] [CrossRef]
- Ng, H.Y.; Leung, W.K.; Cheung, K.S. Association between Gut Microbiota and SARS-CoV-2 Infection and Vaccine Immunogenicity. Microorganisms 2023, 11, 452. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5367. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed]
- Maddah, R.; Goodarzi, V.; Asadi-Yousefabad, S.L.; Abbasluo, M.; Shariati, P.; Shafiei Kafraj, A. Evaluation of the gut microbiome associated with COVID-19. Inform. Med. Unlocked 2023, 38, 101239. [Google Scholar] [CrossRef] [PubMed]
- National Health Commission of the People’s Republic of China. 2023. Available online: http://www.nhc.gov.cn/xcs/yqfkdt/202211/39d316d555094db5b2a6a9eb97214a8b.shtml (accessed on 5 November 2022).
- Dong, Y.; Dai, T.; Wei, Y.; Zhang, L.; Zheng, M.; Zhou, F. A systematic review of SARS-CoV-2 vaccine candidates. Signal Transduct. Target. Ther. 2020, 5, 237. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.N.; Zhao, Y.Y.; Li, L.; Gao, H.D.; Cai, Q.; Sun, X.X.; Zhang, F.S.; Su, J.F.; Zhang, Y.N.; Shu, X.; et al. Evaluation of safety of two inactivated COVID-19 vaccines in a large-scale emergency use. Zhonghua Liu Xing Bing Xue Za Zhi 2021, 42, 977–982. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Chen, Y.; Wu, Z.; Chen, Y.; Gao, H.; Lv, L.; Guo, F.; Zhang, X.; Luo, R.; Huang, C.; et al. Alterations of the Gut Microbiota in Patients With Coronavirus Disease 2019 or H1N1 Influenza. Clin. Infect. Dis. 2020, 71, 2669–2678. [Google Scholar] [CrossRef]
- Zuo, T.; Zhang, F.; Lui, G.C.; Yeoh, Y.K.; Li, A.Y.; Zhan, H.; Wan, Y.; Chung, A.C.; Cheung, C.P.; Chen, N.; et al. Alterations in Gut Microbiota of Patients With COVID-19 During Time of Hospitalization. Gastroenterology 2020, 159, 944–955. [Google Scholar] [CrossRef]
- Lewandowski, K.; Kaniewska, M.; Rosołowski, M.; Rydzewska, G. Gastrointestinal symptoms in COVID-19. Prz. Gastroenterol. 2023, 18, 61–66. [Google Scholar] [CrossRef]
- Dingeo, G.; Brito, A.; Samouda, H.; Iddir, M.; La Frano, M.R.; Bohn, T. Phytochemicals as modifiers of gut microbial communities. Food Funct. 2020, 11, 8444–8471. [Google Scholar] [CrossRef]
- Cao, J.; Wang, C.; Zhang, Y.; Lei, G.; Xu, K.; Zhao, N.; Lu, J.; Meng, F.; Yu, L.; Yan, J.; et al. Integrated gut virome and bacteriome dynamics in COVID-19 patients. Gut Microbes 2021, 13, 1–21. [Google Scholar] [CrossRef]
- Madison, A.A.; Shrout, M.R.; Renna, M.E.; Kiecolt-Glaser, J.K. Psychological and Behavioral Predictors of Vaccine Efficacy: Considerations for COVID-19. Perspect. Psychol. Sci. 2021, 16, 191–203. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Nageshwar Reddy, D. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Li, A.; Huang, T.; Lai, J.; Li, J.; Sublette, M.E.; Lu, H.; Lu, Q.; Du, Y.; Hu, Z.; et al. Gut Microbiota Changes in Patients with Bipolar Depression. Adv. Sci. 2019, 6, 1900752. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Ling, Z.; Zhang, Y.; Mao, H.; Ma, Z.; Yin, Y.; Wang, W.; Tang, W.; Tan, Z.; Shi, J.; et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav. Immun. 2015, 48, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Demirci, M.; Tokman, H.B.; Taner, Z.; Keskin, F.E.; Çağatay, P.; Bakar, Y.O.; Özyazar, M.; Kiraz, N.; Kocazeybek, B.S. Bacteroidetes and Firmicutes levels in gut microbiota and effects of hosts TLR2/TLR4 gene expression levels in adult type 1 diabetes patients in Istanbul, Turkey. J. Diabetes Complicat. 2020, 34, 107449. [Google Scholar] [CrossRef]
- Tanca, A.; Abbondio, M.; Palomba, A.; Fraumene, C.; Manghina, V.; Cucca, F.; Fiorillo, E.; Uzzau, S. Potential and active functions in the gut microbiota of a healthy human cohort. Microbiome 2017, 5, 79. [Google Scholar] [CrossRef]
- Leonel, A.J.; Alvarez-Leite, J.I. Butyrate: Implications for intestinal function. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 474–479. [Google Scholar] [CrossRef]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef]
- Dillon, S.M.; Lee, E.J.; Kotter, C.V.; Austin, G.L.; Dong, Z.; Hecht, D.K.; Gianella, S.; Siewe, B.; Smith, D.M.; Landay, A.L.; et al. An altered intestinal mucosal microbiome in HIV-1 infection is associated with mucosal and systemic immune activation and endotoxemia. Mucosal Immunol. 2014, 7, 983–994. [Google Scholar] [CrossRef]
- Vázquez-Castellanos, J.F.; Serrano-Villar, S.; Latorre, A.; Artacho, A.; Ferrús, M.L.; Madrid, N.; Vallejo, A.; Sainz, T.; Martínez-Botas, J.; Ferrando-Martínez, S.; et al. Altered metabolism of gut microbiota contributes to chronic immune activation in HIV-infected individuals. Mucosal Immunol. 2015, 8, 760–772. [Google Scholar] [CrossRef]
- Dillon, S.M.; Lee, E.J.; Kotter, C.V.; Austin, G.L.; Gianella, S.; Siewe, B.; Smith, D.M.; Landay, A.L.; McManus, M.C.; Robertson, C.E.; et al. Gut dendritic cell activation links an altered colonic microbiome to mucosal and systemic T-cell activation in untreated HIV-1 infection. Mucosal Immunol. 2016, 9, 24–37. [Google Scholar] [CrossRef]
- Hu, H.J.; Park, S.G.; Jang, H.B.; Choi, M.G.; Park, K.H.; Kang, J.H.; Park, S.I.; Lee, H.J.; Cho, S.H. Obesity Alters the Microbial Community Profile in Korean Adolescents. PLoS ONE 2015, 10, e0134333. [Google Scholar] [CrossRef] [PubMed]
- Michail, S.; Lin, M.; Frey, M.R.; Fanter, R.; Paliy, O.; Hilbush, B.; Reo, N.V. Altered gut microbial energy and metabolism in children with non-alcoholic fatty liver disease. FEMS Microbiol. Ecol. 2015, 91, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sandle, G.I.; Herod, M.R.; Fontana, J.; Lippiat, J.D.; Stockley, P.G. Is intestinal transport dysfunctional in COVID-19-related diarrhea? Am. J. Physiol. Gastrointest. Liver Physiol. 2023, 324, G415–G418. [Google Scholar] [CrossRef] [PubMed]
- Mannan, A.; Hoque, M.N.; Noyon, S.H.; Hamidullah Mehedi, H.M.; Foisal, J.; Salauddin, A.; Rafiqul Islam, S.M.; Sharmen, F.; Tanni, A.A.; Siddiki, A.Z.; et al. SARS-CoV-2 infection alters the gut microbiome in diabetes patients: A cross-sectional study from Bangladesh. J. Med. Virol. 2023. [Google Scholar] [CrossRef]
- Copoiu, L.; Malhotra, S. The current structural glycome landscape and emerging technologies. Curr. Opin. Struct. Biol. 2020, 62, 132–139. [Google Scholar] [CrossRef]
- Mattila, J.; Hietakangas, V. Regulation of Carbohydrate Energy Metabolism in Drosophila melanogaster. Genetics 2017, 207, 1231–1253. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, H.; Zhang, L.; Li, R.; Luo, L.; Yu, Z.; Wan, Z. Effects of high-intensity interval training on gut microbiota profiles in 12 months’ old ICR mice. J. Physiol. Biochem. 2020, 76, 539–548. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, Q.; Yang, J.; Zhang, J.; Fu, J.; Dang, C.; Liu, M.; Wang, S.; Lin, Y.; Hao, J.; et al. Significant alterations of intestinal symbiotic microbiota induced by intraperitoneal vaccination mediate changes in intestinal metabolism of NEW Genetically Improved Farmed Tilapia (NEW GIFT, Oreochromis niloticus). Microbiome 2022, 10, 221. [Google Scholar] [CrossRef]
- Petrović, V.; Vuković, V.; Patić, A.; Marković, M.; Ristić, M. Immunogenicity of BNT162b2, BBIBP-CorV and Gam-COVID-Vac vaccines and immunity after natural SARS-CoV-2 infection-A comparative study from Novi Sad, Serbia. PLoS ONE 2022, 17, e0263468. [Google Scholar] [CrossRef]
- Vályi-Nagy, I.; Matula, Z.; Gönczi, M.; Tasnády, S.; Bekő, G.; Réti, M.; Ajzner, É.; Uher, F. Comparison of antibody and T cell responses elicited by BBIBP-CorV (Sinopharm) and BNT162b2 (Pfizer-BioNTech) vaccines against SARS-CoV-2 in healthy adult humans. Geroscience 2021, 43, 2321–2331. [Google Scholar] [CrossRef]
- Kamel, N.A.; El Wakeel, L.M.; Aboshanab, K.M. Exploring SARS-CoV-2 Spikes Glycoproteins for Designing Potential Antiviral Targets. Viral Immunol. 2021, 34, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Hamed, S.M.; Sakr, M.M.; El-Housseiny, G.S.; Wasfi, R.; Aboshanab, K.M. State of the art in epitope mapping and opportunities in COVID-19. Future Sci. OA 2023, 16, FSO832. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unvaccinated (n = 20) | Vaccinated (n = 20) | P Value | |
|---|---|---|---|
| Age, years | 38.5 (29.5–49.0) | 38.5 (30.5–42.5) | 0.799 |
| Male/Female | 12/8 | 9/11 | 0.527 |
| BMI, kg/m2 | 23.4 (20.2–25.5) | 22.9 (21.3–26.5) | 0.745 |
| SBP, mmHg | 120.5 (116.3–126.6) | 120.0 (111.2–127.5) | 0.489 |
| DBP, mmHg | 73.2 (70.0–80.0) | 70.5 (65.3–77.3) | 0.290 |
| FBG, mmol/L | 5.07 (4.63–5.39) | 4.59 (4.38–5.30) | 0.239 |
| Total cholesterol, mmol/L | 4.65 (3.83–5.12) | 4.61 (4.11–5.24) | 0.685 |
| Triglyceride, mmol/L | 1.29 (0.82–1.60) | 0.98 (0.62–1.35) | 0.074 |
| HDLC, mmol/L | 1.15 (0.92–1.57) | 1.37 (1.02–1.70) | 0.318 |
| LDLC, mmol/L | 2.55 (2.25–3.37) | 2.83 (2.39–3.70) | 0.196 |
| Uric acid, umol/L | 321.25 (233.98–386.50) | 378.00 (243.75–407.25) | 0.626 |
| WBC, uL | 6.95 (6.20–7.86) | 5.89 (5.12–7.85) | 0.062 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Y.; Dong, Y.; Jiao, J.; Wang, P.; Chen, M.; Li, J. BBIBP-CorV Vaccination against the SARS-CoV-2 Virus Affects the Gut Microbiome. Vaccines 2023, 11, 942. https://doi.org/10.3390/vaccines11050942
Shen Y, Dong Y, Jiao J, Wang P, Chen M, Li J. BBIBP-CorV Vaccination against the SARS-CoV-2 Virus Affects the Gut Microbiome. Vaccines. 2023; 11(5):942. https://doi.org/10.3390/vaccines11050942
Chicago/Turabian StyleShen, Yang, Ying Dong, Jie Jiao, Pan Wang, Mulei Chen, and Jing Li. 2023. "BBIBP-CorV Vaccination against the SARS-CoV-2 Virus Affects the Gut Microbiome" Vaccines 11, no. 5: 942. https://doi.org/10.3390/vaccines11050942