

Vaccination of Zoo Birds against West Nile Virus—A Field Study
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Vaccination
2.3. Animals
2.4. Diagnostic Methods
2.5. Sequencing and Phylogenetic Analysis
2.6. Statistical Analysis
2.7. Ethical Statement
3. Results
3.1. Adverse Reactions of Vaccination
3.2. Molecular Biological Results
3.3. Serological Results
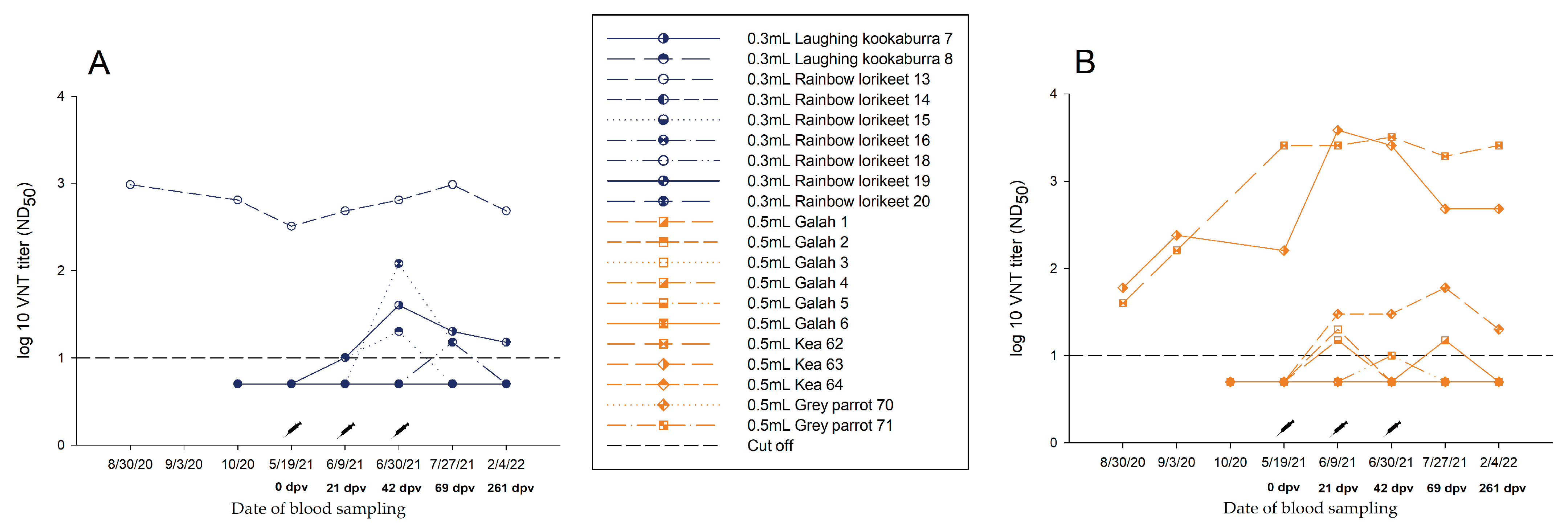
3.3.1. Group A—0.3 mL Dose Regimen Vaccinated Three Times
3.3.2. Group B—0.5 mL Dose Regimen Vaccinated Three Times
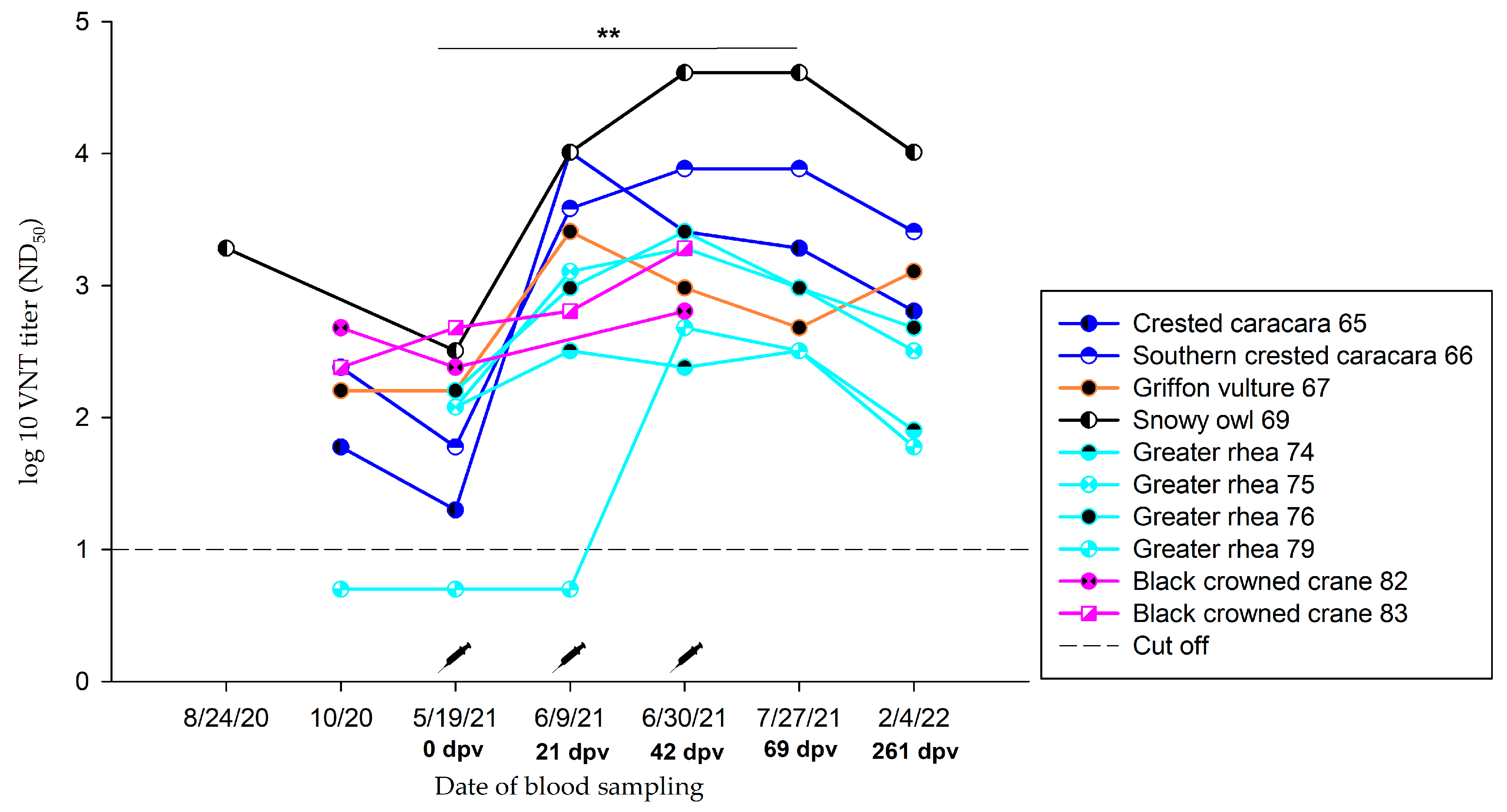
3.3.3. Group C.1—1.0 mL Dose Regimen Vaccinated Three Times
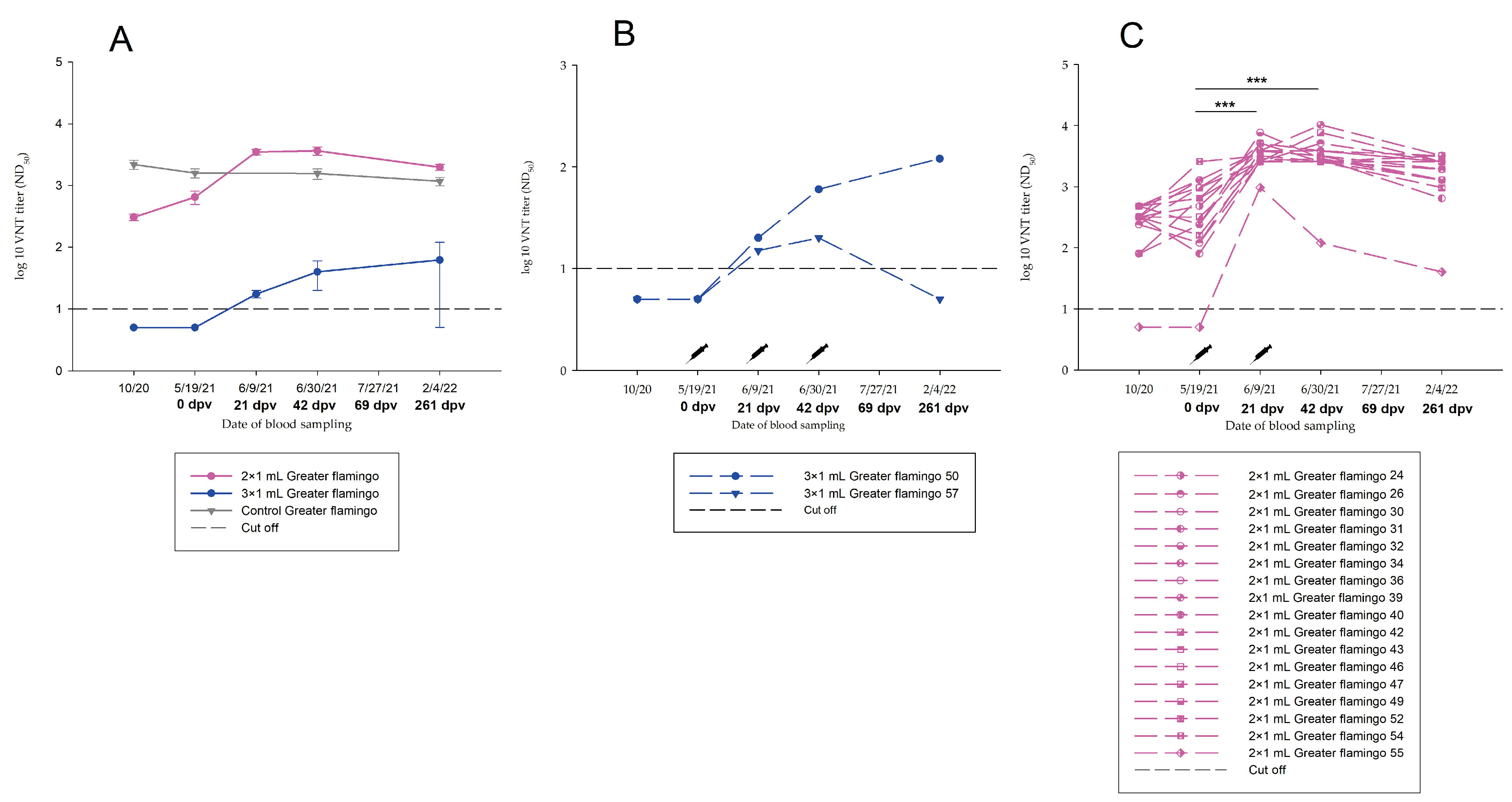
3.3.4. Group C.2—1.0 mL Dose Regimen Vaccinated Twice
3.3.5. Group C.3—1.0 mL Dose Regimen Vaccinated Once
3.3.6. Group D—Control Group without Vaccination
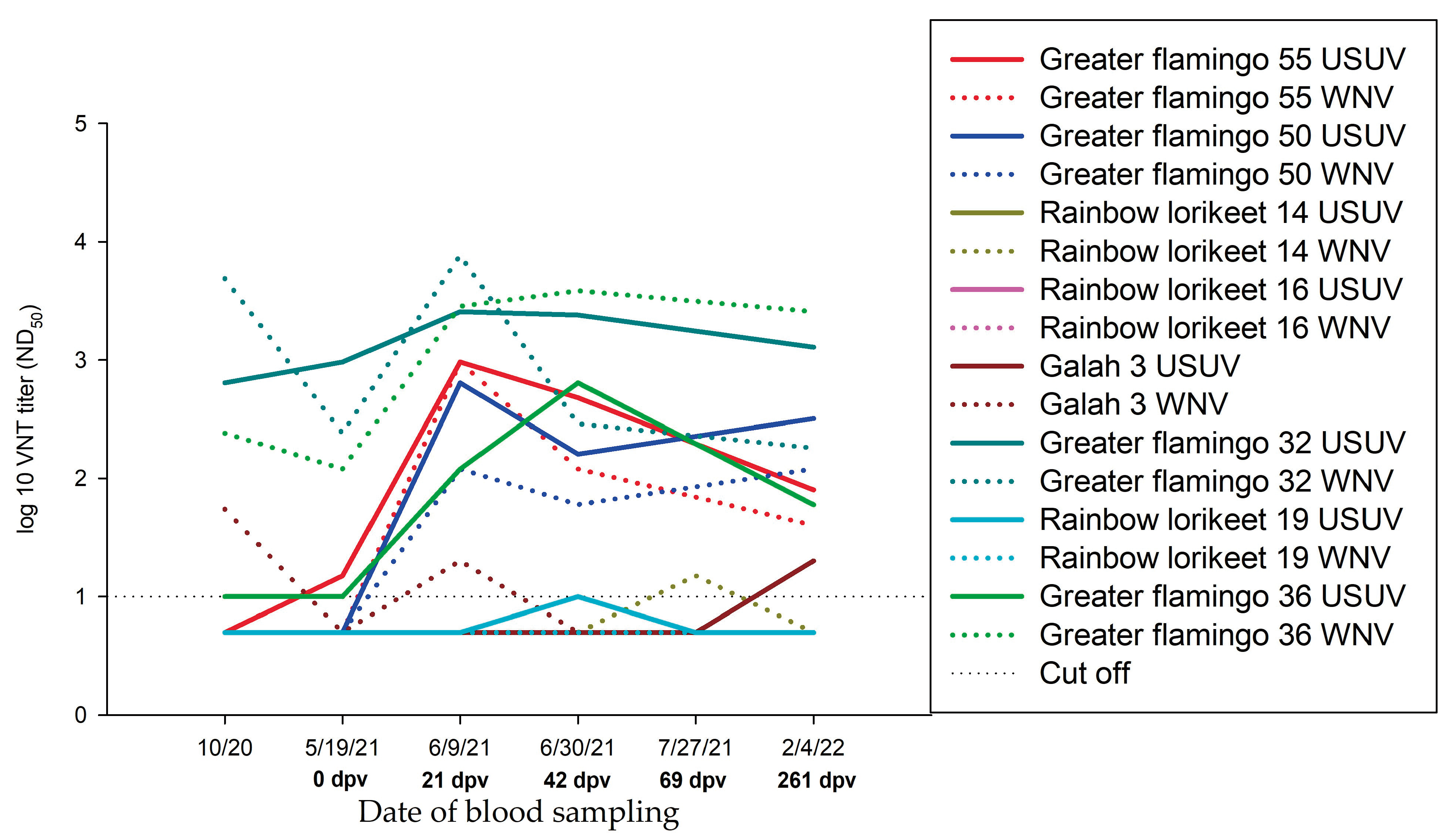
3.3.7. Summary of the Serological Results of Greater Flamingos
3.4. Usutu Virus Circulation in the Zoo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Engler, O.; Savini, G.; Papa, A.; Figuerola, J.; Groschup, M.H.; Kampen, H.; Medlock, J.; Vaux, A.; Wilson, A.J.; Werner, D.; et al. European surveillance for West Nile virus in mosquito populations. Int. J. Environ. Res. Public Health 2013, 10, 4869–4895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, G.L.; Marfin, A.A.; Lanciotti, R.S.; Gubler, D.J. West Nile virus. Lancet Infect. Dis. 2002, 2, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Nikolay, B. A review of West Nile and Usutu virus co-circulation in Europe: How much do transmission cycles overlap? Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 609–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Madrid, A.T.; Porterfield, J.S. The flaviviruses (group B arboviruses): A cross-neutralization study. J. Gen. Virol. 1974, 23, 91–96. [Google Scholar] [CrossRef]
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The global ecology and epidemiology of West Nile virus. Biomed Res. Int. 2015, 2015, 376230. [Google Scholar] [CrossRef] [Green Version]
- Chang, G.-J.J.; Davis, B.S.; Stringfield, C.; Lutz, C. Prospective immunization of the endangered California condors (Gymnogyps californianus) protects this species from lethal West Nile virus infection. Vaccine 2007, 25, 2325–2330. [Google Scholar] [CrossRef]
- Kramer, L.D.; Styer, L.M.; Ebel, G.D. A global perspective on the epidemiology of West Nile virus. Annu. Rev. Entomol. 2008, 53, 61–81. [Google Scholar] [CrossRef] [Green Version]
- Pietsch, C.; Michalski, D.; Münch, J.; Petros, S.; Bergs, S.; Trawinski, H.; Lübbert, C.; Liebert, U.G. Autochthonous West Nile virus infection outbreak in humans, Leipzig, Germany, August to September 2020. Eurosurveill 2020, 25, 2001786. [Google Scholar] [CrossRef]
- Ziegler, U.; Bergmann, F.; Fischer, D.; Müller, K.; Holicki, C.M.; Sadeghi, B.; Sieg, M.; Keller, M.; Schwehn, R.; Reuschel, M.; et al. Spread of West Nile Virus and Usutu Virus in the German Bird Population, 2019–2020. Microorganisms 2022, 10, 807. [Google Scholar] [CrossRef]
- Yeung, M.W.; Shing, E.; Nelder, M.; Sander, B. Epidemiologic and clinical parameters of West Nile virus infections in humans: A scoping review. BMC Infect. Dis. 2017, 17, 609. [Google Scholar] [CrossRef] [Green Version]
- Wünschmann, A.; Shivers, J.; Bender, J.; Carroll, L.; Fuller, S.; Saggese, M.; van Wettere, A.; Redig, P. Pathologic and immunohistochemical findings in goshawks (Accipiter gentilis) and great horned owls (Bubo virginianus) naturally infected with West Nile virus. Avian Dis. 2005, 49, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.S.; Roehrig, J.T.; Deubel, V.; Smith, J.; Parker, M.; Steele, K.; Crise, B.; Volpe, K.E.; Crabtree, M.B.; Scherret, J.H.; et al. Origin of the West Nile virus responsible for an outbreak of encephalitis in the northeastern United States. Science 1999, 286, 2333–2337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komar, N.; Langevin, S.; Hinten, S.; Nemeth, N.; Edwards, E.; Hettler, D.; Davis, B.; Bowen, R.; Bunning, M. Experimental infection of North American birds with the New York 1999 strain of West Nile virus. Emerg. Infect. Dis. 2003, 9, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, U.; Angenvoort, J.; Fischer, D.; Fast, C.; Eiden, M.; Rodriguez, A.V.; Revilla-Fernández, S.; Nowotny, N.; de La Fuente, J.G.; Lierz, M.; et al. Pathogenesis of West Nile virus lineage 1 and 2 in experimentally infected large falcons. Vet. Microbiol. 2013, 161, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Troupin, A.; Colpitts, T.M. Overview of West Nile Virus Transmission and Epidemiology. Methods Mol. Biol. 2016, 1435, 15–18. [Google Scholar] [CrossRef] [PubMed]
- McLean, R.G.; Ubico, S.R.; Bourne, D.; Komar, N. West Nile virus in livestock and wildlife. Curr. Top. Microbiol. Immunol. 2002, 267, 271–308. [Google Scholar] [CrossRef]
- Zannoli, S.; Sambri, V. West Nile Virus and Usutu Virus Co-Circulation in Europe: Epidemiology and Implications. Microorganisms 2019, 7, 184. [Google Scholar] [CrossRef] [Green Version]
- Santos, P.D.; Michel, F.; Wylezich, C.; Höper, D.; Keller, M.; Holicki, C.M.; Szentiks, C.A.; Eiden, M.; Muluneh, A.; Neubauer-Juric, A.; et al. Co-infections: Simultaneous detections of West Nile virus and Usutu virus in birds from Germany. Transbound. Emerg. Dis. 2021, 69, 776–792. [Google Scholar] [CrossRef]
- Weissenböck, H.; Kolodziejek, J.; Url, A.; Lussy, H.; Rebel-Bauder, B.; Nowotny, N. Emergence of Usutu virus, an African mosquito-borne flavivirus of the Japanese encephalitis virus group, central Europe. Emerg. Infect. Dis. 2002, 8, 652–656. [Google Scholar] [CrossRef]
- Becker, N.; Jöst, H.; Ziegler, U.; Eiden, M.; Höper, D.; Emmerich, P.; Fichet-Calvet, E.; Ehichioya, D.U.; Czajka, C.; Gabriel, M.; et al. Epizootic emergence of Usutu virus in wild and captive birds in Germany. PLoS ONE 2012, 7, e32604. [Google Scholar] [CrossRef]
- Cadar, D.; Lühken, R.; van der Jeugd, H.; Garigliany, M.; Ziegler, U.; Keller, M.; Lahoreau, J.; Lachmann, L.; Becker, N.; Kik, M.; et al. Widespread activity of multiple lineages of Usutu virus, western Europe, 2016. Eurosurveill 2017, 22, 30452. [Google Scholar] [CrossRef] [Green Version]
- Jöst, H.; Bialonski, A.; Maus, D.; Sambri, V.; Eiden, M.; Groschup, M.H.; Günther, S.; Becker, N.; Schmidt-Chanasit, J. Isolation of usutu virus in Germany. Am. J. Trop. Med. Hyg. 2011, 85, 551–553. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, U.; Lühken, R.; Keller, M.; Cadar, D.; van der Grinten, E.; Michel, F.; Albrecht, K.; Eiden, M.; Rinder, M.; Lachmann, L.; et al. West Nile virus epizootic in Germany, 2018. Antivir. Res. 2019, 162, 39–43. [Google Scholar] [CrossRef]
- Michel, F.; Sieg, M.; Fischer, D.; Keller, M.; Eiden, M.; Reuschel, M.; Schmidt, V.; Schwehn, R.; Rinder, M.; Urbaniak, S.; et al. Evidence for West Nile Virus and Usutu Virus Infections in Wild and Resident Birds in Germany, 2017 and 2018. Viruses 2019, 11, 674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, U.; Jöst, H.; Müller, K.; Fischer, D.; Rinder, M.; Tietze, D.T.; Danner, K.-J.; Becker, N.; Skuballa, J.; Hamann, H.-P.; et al. Epidemic Spread of Usutu Virus in Southwest Germany in 2011 to 2013 and Monitoring of Wild Birds for Usutu and West Nile Viruses. Vector-Borne Zoonotic Dis. 2015, 15, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Seidowski, D.; Ziegler, U.; von Rönn, J.A.C.; Müller, K.; Hüppop, K.; Müller, T.; Freuling, C.; Mühle, R.-U.; Nowotny, N.; Ulrich, R.G.; et al. West Nile virus monitoring of migratory and resident birds in Germany. Vector-Borne Zoonotic Dis. 2010, 10, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Michel, F.; Fischer, D.; Eiden, M.; Fast, C.; Reuschel, M.; Müller, K.; Rinder, M.; Urbaniak, S.; Brandes, F.; Schwehn, R.; et al. West Nile Virus and Usutu Virus Monitoring of Wild Birds in Germany. Int. J. Environ. Res. Public Health 2018, 15, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, F.; Trachsel, D.S.; Stoeckle, S.D.; Bernis Sierra, J.; Lübke, S.; Groschup, M.H.; Gehlen, H.; Ziegler, U. Seroepidemiological Survey of West Nile Virus Infections in Horses from Berlin/Brandenburg and North Rhine-Westphalia, Germany. Viruses 2022, 14, 243. [Google Scholar] [CrossRef]
- Petersen, L.R. Epidemiology of West Nile Virus in the United States: Implications for Arbovirology and Public Health. J. Med. Entomol. 2019, 56, 1456–1462. [Google Scholar] [CrossRef] [Green Version]
- Frank, C.; Schmidt-Chanasit, J.; Ziegler, U.; Lachmann, R.; Preußel, K.; Offergeld, R. West Nile Virus in Germany: An Emerging Infection and Its Relevance for Transfusion Safety. Transfus. Med. Hemother. 2022, 49, 192–204. [Google Scholar] [CrossRef]
- Friedrich-Loeffler-Institut. West-Nil-Virus. Available online: https://www.fli.de/de/aktuelles/tierseuchengeschehen/west-nil-virus/ (accessed on 13 October 2022).
- Nasci, R.S.; Mutebi, J.-P. Reducing West Nile Virus Risk Through Vector Management. J. Med. Entomol. 2019, 56, 1516–1521. [Google Scholar] [CrossRef] [PubMed]
- StIKoVet. Stellungnahme zur Immunisierung von Pferden gegen das West-Nil-Virus. Available online: https://www.openagrar.de/servlets/MCRFileNodeServlet/openagrar_derivate_00017232/Stellungnahme_WNV-Impfung_Pferde_2018-10-22.pdf (accessed on 6 February 2023).
- Davis, M.R.; Langan, J.N.; Johnson, Y.J.; Ritchie, B.W.; van Bonn, W. West Nile virus seroconversion in penguins after vaccination with a killed virus vaccine or a DNA vaccine. J. Zoo Wildl. Med. 2008, 39, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Okeson, D.M.; Llizo, S.Y.; Miller, C.L.; Glaser, A.L. Antibody Response of Five Bird Species after Vaccination with a Killed West Nile Virus Vaccine. J. Zoo Wildl. Med. 2007, 38, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Nusbaum, K.E.; Wright, J.C.; Johnston, W.B.; Allison, A.B.; Hilton, C.D.; Staggs, L.A.; Stallknecht, D.E.; Shelnutt, J.L. Absence of humoral response in flamingos and red-tailed hawks to experimental vaccination with a killed West Nile virus vaccine. Avian Dis. 2003, 47, 750–752. [Google Scholar] [CrossRef]
- Siegal-Willott, J.L.; Carpenter, J.W.; Glaser, A.L. Lack of Detectable Antibody Response in Greater Flamingos (Phoenicopterus ruber ruber) after Vaccination against West Nile Virus with a Killed Equine Vaccine. J. Avian Med. Surg. 2006, 20, 89–93. [Google Scholar] [CrossRef]
- Baitchman, E.J.; Tlusty, M.F.; Murphy, H.W. Passive Transfer of Maternal Antibodies to West Nile Virus in Flamingo Chicks (Phoenicoperius Chilensis and Phoenicopterus Ruber Ruber). J. Zoo Wildl. Med. 2007, 38, 337–340. [Google Scholar] [CrossRef]
- Johnson, S. Avian titer development against West nile virus after extralabel use of an equine vaccine. J. Zoo Wildl. Med. 2005, 36, 257–264. [Google Scholar] [CrossRef]
- Redig, P.T.; Tully, T.N.; Ritchie, B.W.; Roy, A.F.; Baudena, M.A.; Chang, G.-J.J. Effect of West Nile virus DNA-plasmid vaccination on response to live virus challenge in red-tailed hawks (Buteo jamaicensis). Am. J. Vet. Res. 2011, 72, 1065–1070. [Google Scholar] [CrossRef]
- Angenvoort, J.; Fischer, D.; Fast, C.; Ziegler, U.; Eiden, M.; de La Fuente, J.G.; Lierz, M.; Groschup, M.H. Limited efficacy of West Nile virus vaccines in large falcons (Falco spp.). Vet. Res. 2014, 45, 41. [Google Scholar] [CrossRef] [Green Version]
- Bertelsen, M.F.; Olberg, R.-A.; Crawshaw, G.J.; Dibernardo, A.; Lindsay, L.R.; Drebot, M.; Barker, I.K. West Nile virus infection in the eastern loggerhead shrike (Lanius ludovicianus migrans): Pathology, epidemiology, and immunization. J. Wildl. Dis. 2004, 40, 538–542. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, S.S.; Langevin, S.; Woods, L.; Carroll, B.D.; Vickers, W.; Morrison, S.A.; Chang, G.-J.J.; Reisen, W.K.; Boyce, W.M. Efficacy of three vaccines in protecting Western Scrub-Jays (Aphelocoma californica) from experimental infection with West Nile virus: Implications for vaccination of Island Scrub-Jays (Aphelocoma insularis). Vector-Borne Zoonotic Dis. 2011, 11, 1069–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glavis, J.; Larsen, R.S.; Lamberski, N.; Gaffney, P.; Gardner, I. Evaluation of antibody response to vaccination against West Nile virus in thick billed parrots (Rhynchopsitta pachyrhyncha). J. Zoo Wildl. Med. 2011, 42, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Olsen, G.H.; Miller, K.J.; Docherty, D.E.; Bochsler, V.S.; Sileo, L. Pathogenicity of West Nile virus and response to vaccination in sandhill cranes (Grus canadensis) using a killed vaccine. J. Zoo Wildl. Med. 2009, 40, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Fynmore, N.; Lühken, R.; Maisch, H.; Risch, T.; Merz, S.; Kliemke, K.; Ziegler, U.; Schmidt-Chanasit, J.; Becker, N. Rapid assessment of West Nile virus circulation in a German zoo based on honey-baited FTA cards in combination with box gravid traps. Parasit. Vectors 2021, 14, 449. [Google Scholar] [CrossRef]
- Eiden, M.; Vina-Rodriguez, A.; Hoffmann, B.; Ziegler, U.; Groschup, M.H. Two new real-time quantitative reverse transcription polymerase chain reaction assays with unique target sites for the specific and sensitive detection of lineages 1 and 2 West Nile virus strains. J. Vet. Diagn. Investig. 2010, 22, 748–753. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef]
- Mayr, A.; Bachmann, P.A.; Bibrack, B.; Wittmann, G. Neutralisationstest: Virologische Arbeitsmethoden (Serologie), 1st ed.; Gustav Fischer Verlag: Jena, Germany, 1977. [Google Scholar]
- Yeh, J.-Y.; Lee, J.-H.; Park, J.-Y.; Seo, H.-J.; Moon, J.-S.; Cho, I.-S.; Kim, H.-P.; Yang, Y.-J.; Ahn, K.-M.; Kyung, S.-G.; et al. A diagnostic algorithm to serologically differentiate West Nile virus from Japanese encephalitis virus infections and its validation in field surveillance of poultry and horses. Vector-Borne Zoonotic Dis. 2012, 12, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Holicki, C.M.; Bergmann, F.; Stoek, F.; Schulz, A.; Groschup, M.H.; Ziegler, U.; Sadeghi, B. Expedited retrieval of high-quality Usutu virus genomes via Nanopore sequencing with and without target enrichment. Front. Microbiol. 2022, 13, 1044316. [Google Scholar] [CrossRef]
- Quick, J.; Grubaugh, N.D.; Pullan, S.T.; Claro, I.M.; Smith, A.D.; Gangavarapu, K.; Oliveira, G.; Robles-Sikisaka, R.; Rogers, T.F.; Beutler, N.A.; et al. Multiplex PCR method for MinION and Illumina sequencing of Zika and other virus genomes directly from clinical samples. Nat. Protoc. 2017, 12, 1261–1276. [Google Scholar] [CrossRef] [Green Version]
- Sikkema, R.S.; Schrama, M.; van den Berg, T.; Morren, J.; Munger, E.; Krol, L.; van der Beek, J.G.; Blom, R.; Chestakova, I.; van der Linden, A.; et al. Detection of West Nile virus in a common whitethroat (Curruca communis) and Culex mosquitoes in The Netherlands, 2020. Eurosurveill 2020, 25, 2001704. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Library of Medicine; National Center for Biotechnology Information. Basic Local Alignment Search Tool (BLAST). Version 2.13.0. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 25 January 2023).
- Santos, P.D.; Günther, A.; Keller, M.; Homeier-Bachmann, T.; Groschup, M.H.; Beer, M.; Höper, D.; Ziegler, U. An advanced sequence clustering and designation workflow reveals the enzootic maintenance of a dominant West Nile virus subclade in Germany. bioRxiv 2022. Pre-print. [Google Scholar] [CrossRef]
- Friedrich-Loeffler-Institut. West-Nil-Virus Auch in der Saison 2021 Wieder Aktiv. Available online: https://www.fli.de/de/aktuelles/kurznachrichten/neues-einzelansicht/west-nil-virus-auch-in-der-saison-2021-wieder-aktiv/ (accessed on 19 October 2022).
- Poidinger, M.; Hall, R.A.; Mackenzie, J.S. Molecular characterization of the Japanese encephalitis serocomplex of the flavivirus genus. Virology 1996, 218, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, G.V.; Calle, P.P.; Mangiafico, J.A.; Raphael, B.L.; Danner, D.K.; Hile, J.A.; Clippinger, T.L.; Smith, J.F.; Cook, R.A.; McNamara, T. An outbreak of West Nile virus in a New York City captive wildlife population. Am. J. Trop. Med. Hyg. 2002, 67, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, N.M.; Bosco-Lauth, A.M.; Bowen, R.A. Cross-protection between West Nile and Japanese encephalitis viruses in red-winged blackbirds (Agelaius phoeniceus). Avian Dis. 2009, 53, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, N.M.; Kratz, G.E.; Bates, R.; Scherpelz, J.A.; Bowen, R.A.; Komar, N. Naturally induced humoral immunity to West Nile virus infection in raptors. Ecohealth 2008, 5, 298–304. [Google Scholar] [CrossRef]
- Wodak, E.; Richter, S.; Bagó, Z.; Revilla-Fernández, S.; Weissenböck, H.; Nowotny, N.; Winter, P. Detection and molecular analysis of West Nile virus infections in birds of prey in the eastern part of Austria in 2008 and 2009. Vet. Microbiol. 2011, 149, 358–366. [Google Scholar] [CrossRef] [Green Version]
- Carboni, D.A.; Nevarez, J.G.; Tully, T.N.; Evans, D.E. West Nile virus infection in a sun conure (Aratinga solstitialis). J. Avian Med. Surg. 2008, 22, 240–245. [Google Scholar] [CrossRef]
- Burt, F.J.; Grobbelaar, A.A.; Leman, P.A.; Anthony, F.S.; Gibson, G.V.F.; Swanepoel, R. Phylogenetic relationships of southern African West Nile virus isolates. Emerg. Infect. Dis. 2002, 8, 820–826. [Google Scholar] [CrossRef]
- Fischer, D.; Muir, A.; Aparici Plaza, D.; Herrmann, K.; Pynnonen-Oudman, K. EAZA Usutu and West Nile Virus Management Guidelines; EAZA Executive Office: Amsterdam, The Netherlands, 2019; pp. 1–27. [Google Scholar]
- Calisher, C.H.; Karabatsos, N.; Dalrymple, J.M.; Shope, R.E.; Porterfield, J.S.; Westaway, E.G.; Brandt, W.E. Antigenic relationships between flaviviruses as determined by cross-neutralization tests with polyclonal antisera. J. Gen. Virol. 1989, 70, 37–43. [Google Scholar] [CrossRef]
- Nikolay, B.; Fall, G.; Boye, C.S.B.; Sall, A.A.; Skern, T. Validation of a structural comparison of the antigenic characteristics of Usutu virus and West Nile virus envelope proteins. Virus Res. 2014, 189, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Oliphant, T.; Nybakken, G.E.; Engle, M.; Xu, Q.; Nelson, C.A.; Sukupolvi-Petty, S.; Marri, A.; Lachmi, B.-E.; Olshevsky, U.; Fremont, D.H.; et al. Antibody recognition and neutralization determinants on domains I and II of West Nile Virus envelope protein. J. Virol. 2006, 80, 12149–12159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, S.; Redfern, J.B.; Lidbury, B.A.; Mahalingam, S. Antibody-dependent enhancement and vaccine development. Expert Rev. Vaccines 2006, 5, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Blázquez, A.-B.; Escribano-Romero, E.; Martín-Acebes, M.A.; Petrovic, T.; Saiz, J.-C. Limited susceptibility of mice to Usutu virus (USUV) infection and induction of flavivirus cross-protective immunity. Virology 2015, 482, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Sinigaglia, A.; Pacenti, M.; Martello, T.; Pagni, S.; Franchin, E.; Barzon, L. West Nile virus infection in individuals with pre-existing Usutu virus immunity, northern Italy, 2018. Eurosurveill 2019, 24, 1900261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porterfield, J.S. Antibody-Dependent Enhancement of Viral Infectivity. Adv. Virus Res. 1986, 31, 335–355. [Google Scholar]
- Percivalle, E.; Cassaniti, I.; Sarasini, A.; Rovida, F.; Adzasehoun, K.M.G.; Colombini, I.; Isernia, P.; Cuppari, I.; Baldanti, F. West Nile or Usutu Virus? A Three-Year Follow-Up of Humoral and Cellular Response in a Group of Asymptomatic Blood Donors. Viruses 2020, 12, 157. [Google Scholar] [CrossRef] [Green Version]
- Price, W.H.; Thind, I.S. The mechanism of cross-protection afforded by dengue virus against West Nile virus in hamsters. J. Hyg. 1972, 70, 611–617. [Google Scholar] [CrossRef] [Green Version]
- Tesh, R.B.; Arroyo, J.; Da Travassos Rosa, A.P.A.; Guzman, H.; Xiao, S.-Y.; Monath, T.P. Efficacy of killed virus vaccine, live attenuated chimeric virus vaccine, and passive immunization for prevention of West Nile virus encephalitis in hamster model. Emerg. Infect. Dis. 2002, 8, 1392–1397. [Google Scholar] [CrossRef]
- Avian Medicine: Principles and Application: Defense Mechanisms of the Avian Host; Gerlach, H. (Ed.) Wingers Publishing Inc.: Lake Worth, FL, USA, 1994; ISBN 0-9636996-0-1. [Google Scholar]
- Brien, J.D.; Uhrlaub, J.L.; Nikolich-Zugich, J. Protective capacity and epitope specificity of CD8(+) T cells responding to lethal West Nile virus infection. Eur. J. Immunol. 2007, 37, 1855–1863. [Google Scholar] [CrossRef]
- Engle, M.J.; Diamond, M.S. Antibody prophylaxis and therapy against West Nile virus infection in wild-type and immunodeficient mice. J. Virol. 2003, 77, 12941–12949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purtha, W.E.; Myers, N.; Mitaksov, V.; Sitati, E.; Connolly, J.; Fremont, D.H.; Hansen, T.H.; Diamond, M.S. Antigen-specific cytotoxic T lymphocytes protect against lethal West Nile virus encephalitis. Eur. J. Immunol. 2007, 37, 1845–1854. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, S.E.J.; Hoffman, D.M.; Stark, L.M.; Marlenee, N.L.; Blitvich, B.J.; Beaty, B.J.; Stallknecht, D.E. Persistence of antibodies to West Nile virus in naturally infected rock pigeons (Columba livia). Clin. Diagn. Lab. Immunol. 2005, 12, 665–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Species Order | Common Name | Scientific Name | No. Birds | 1st Vaccination | 2nd Vaccination | 3rd Vaccination |
|---|---|---|---|---|---|---|---|
| A | Coraciiformes | Laughing kookaburra | Dacelo novaeguineae | 2 | 0.3 mL | 0.3 mL | 0.3 mL |
| A | Coraciiformes | Rainbow lorikeet | Trichoglossus haematodus | 7 | 0.3 mL | 0.3 mL | 0.3 mL |
| B | Psittaciformes | Galah | Eolophus roseicapilla | 6 | 0.5 mL | 0.5 mL | 0.5 mL |
| B | Psittaciformes | Kea | Nestor notabilis | 3 | 0.5 mL | 0.5 mL | 0.5 mL |
| B | Psittaciformes | Grey parrot | Psittacus erithacus | 2 | 0.5 mL | 0.5 mL | 0.5 mL |
| C.1 | Falconiformes | Southern crested caracara | Caracara plancus | 1 | 1.0 mL | 1.0 mL | 1.0 mL |
| C.1 | Falconiformes | Crested caracara | Caracara cheriway | 1 | 1.0 mL | 1.0 mL | 1.0 mL |
| C.1 | Accipitriformes | Griffon vulture | Gyps fulvus | 1 * | 1.0 mL | 1.0 mL | 1.0 mL |
| C.1 | Rheiformes | Greater rhea | Rhea americana | 4 | 1.0 mL | 1.0 mL | 1.0 mL |
| C.1 | Gruiformes | Black crowned crane | Balearica pavonina | 2 | 1.0 mL | 1.0 mL | 1.0 mL |
| C.1 | Strigiformes | Snowy owl | Bubo scandiacus | 1 | 1.0 mL | 1.0 mL | 1.0 mL |
| C.1 | Phoenicopteriformes | Greater flamingo | Phoenicopterus roseus | 2 | 1.0 mL | 1.0 mL | 1.0 mL |
| C.2 | Phoenicopteriformes | Greater flamingo | Phoenicopterus roseus | 17 | 1.0 mL | 1.0 mL | NV |
| C.3 | Pelecaniformes | Northern bald ibis | Geronticus eremita | 13 | 1.0 mL | NV | NV |
| D.1 | Rheiformes | Greater rhea | Rhea americana | 1 | NV | NV | NV |
| D.1 | Anseriformes | Northern shoveler | Spatula clypeata | 5 | NV | NV | NV |
| D.1 | Galliformes | Thuringer chicken | Gallus gallus domesticus | 5 | NV | NV | NV |
| D.1 | Galliformes | Sebright bantam chicken | Gallus gallus domesticus | 3 | NV | NV | NV |
| D.1 | Anseriformes | Indian runner duck | Anas platyrhynchos f. domestica | 4 | NV | NV | NV |
| D.1 | Anseriformes | Steinbach fighting goose | Anser anser f. domestica | 4 | NV | NV | NV |
| D.1 | Passeriformes | Bali myna | Leucopsar rothschildi | 2 | NV | NV | NV |
| D.1 | Galliformes | Common peafowl | Pavo cristatus | 2 | NV | NV | NV |
| D.1 | Coraciiformes | Rainbow lorikeet | Trichoglossus haematodus | 1 | NV | NV | NV |
| D.1 | Coraciiformes | Laughing kookaburra | Dacelo novaeguineae | 4 | NV | NV | NV |
| D.1 | Accipitriformes | Griffon vulture | Gyps fulvus | 1 | NV | NV | NV |
| D.2 | Phoenicopteriformes | Greater flamingo | Phoenicopterus roseus | 20 | NV | NV | NV |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bergmann, F.; Fischer, D.; Fischer, L.; Maisch, H.; Risch, T.; Dreyer, S.; Sadeghi, B.; Geelhaar, D.; Grund, L.; Merz, S.; et al. Vaccination of Zoo Birds against West Nile Virus—A Field Study. Vaccines 2023, 11, 652. https://doi.org/10.3390/vaccines11030652
Bergmann F, Fischer D, Fischer L, Maisch H, Risch T, Dreyer S, Sadeghi B, Geelhaar D, Grund L, Merz S, et al. Vaccination of Zoo Birds against West Nile Virus—A Field Study. Vaccines. 2023; 11(3):652. https://doi.org/10.3390/vaccines11030652
Chicago/Turabian StyleBergmann, Felicitas, Dominik Fischer, Luisa Fischer, Heike Maisch, Tina Risch, Saskia Dreyer, Balal Sadeghi, Dietmar Geelhaar, Lisa Grund, Sabine Merz, and et al. 2023. "Vaccination of Zoo Birds against West Nile Virus—A Field Study" Vaccines 11, no. 3: 652. https://doi.org/10.3390/vaccines11030652