

Protective Efficacy of Multiple Epitope-Based Vaccine against Hyalomma anatolicum, Vector of Theileria annulata and Crimean–Congo Hemorrhagic Fever Virus
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Maintenance of H. anatolicum Ticks for Challenge Study
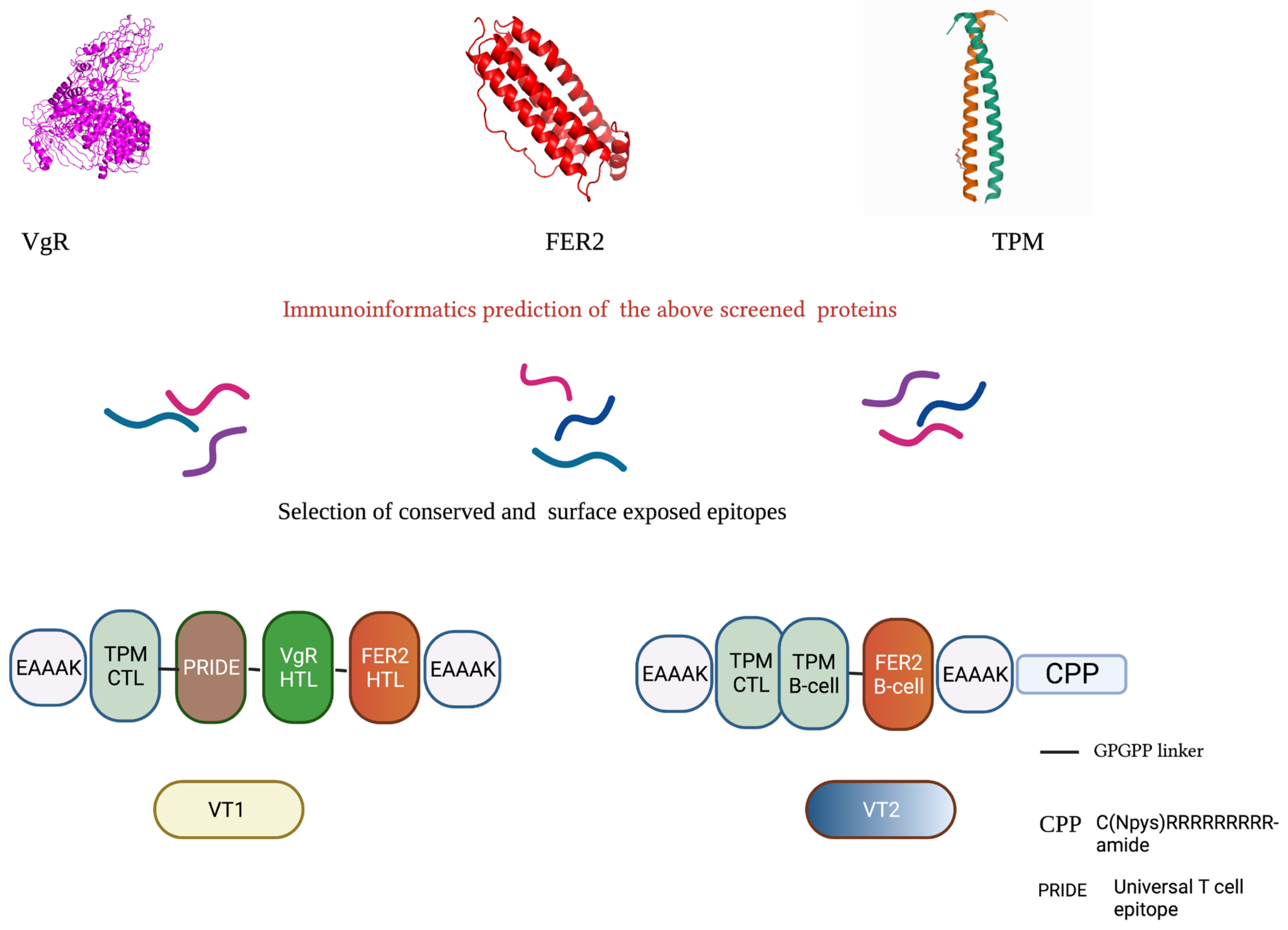
2.3. In Silico Designing of Multi-Epitopic Peptide Constructs
2.3.1. Prediction of Nature and Localization
2.3.2. Prediction of BandT-Cell Epitopes and Construction of Multi-Epitope Peptide Sequence
2.3.3. Evaluation of Antigenic and Physicochemical Properties of the Constructs
2.3.4. Prediction of Secondary and Tertiary Structure of the Constructs
2.3.5. Refinement and Validation of the Tertiary Structure
2.3.6. Molecular Docking of the Constructs with Bovine TLR-2, TLR-4, and TLR-9
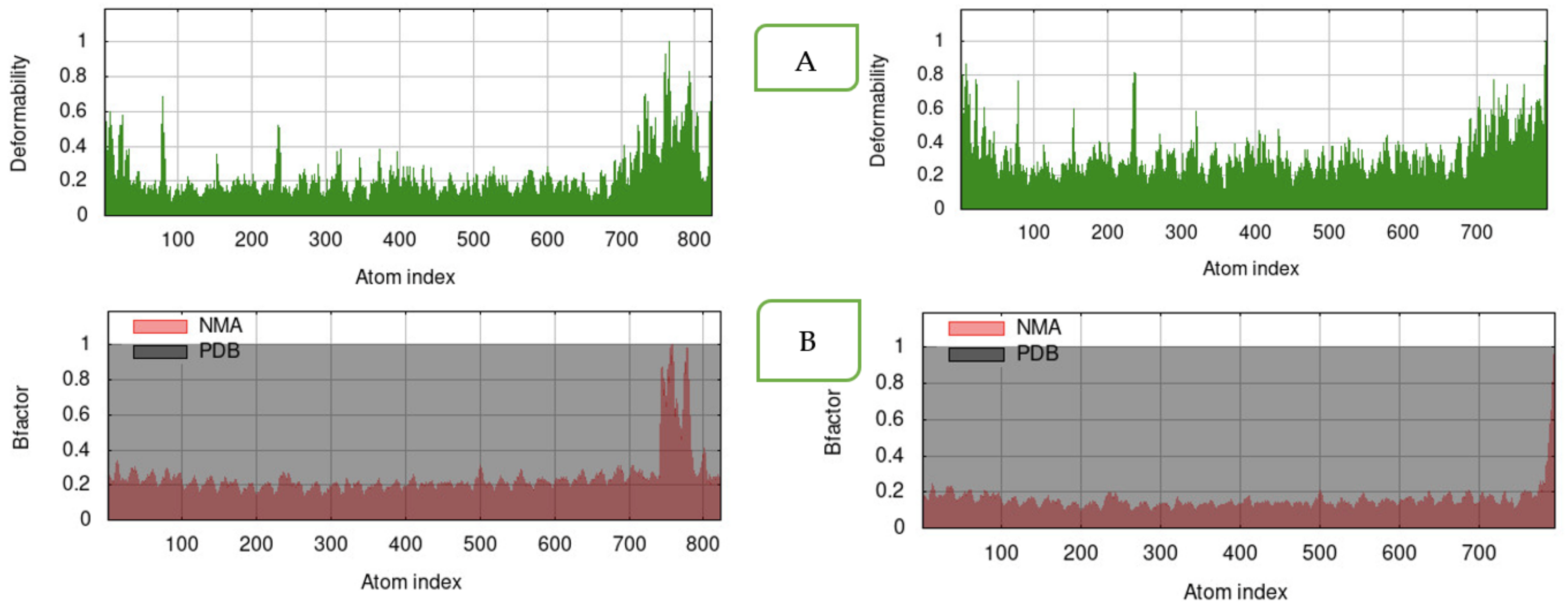
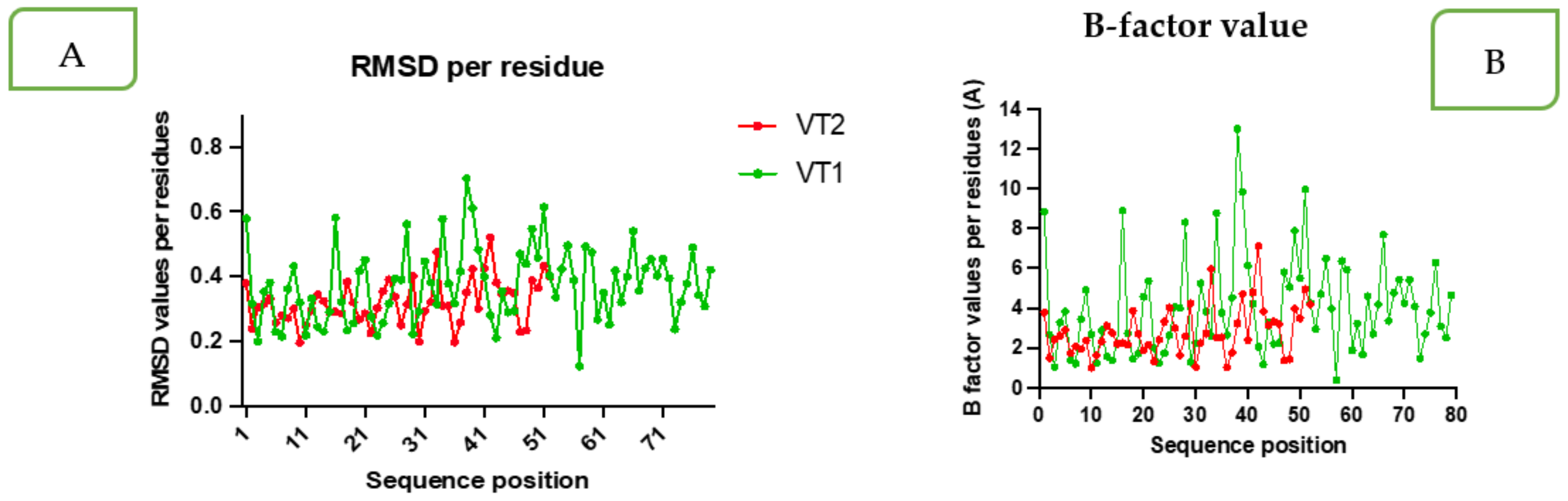
2.3.7. Molecular Dynamics Simulation (MDS)
2.4. Synthesis, Formulation and Storage of MEPs
2.5. Standardization of Immunization Doses
2.6. Standardization of ELISA for Monitoring of IgG and IgM Response
2.7. In Vivo Immunization and Challenge Study
2.8. Immune Response Monitoring
2.9. Expression Profile of Cytokines (IL-2, IL-4, IL-5, and INF-γ) by Q-PCR
2.9.1. Optimization of Q-PCR
2.9.2. Relative Gene Expression Profile
2.9.3. Quantification of Cytokines (IL-2, IL-4, and INF-γ) by ELISA
2.10. Statistical Analysis
3. Results
3.1. Nature and Sub-Cellular Localization of the Selected Proteins
3.2. Immunodominant Epitope Prediction and Their Characteristics
3.3. Prediction of Antigenic and Physiochemical Properties of the Constructs
3.4. Predicted Secondary and Tertiary Structure
3.5. Refinement and Validation of the Tertiary Constructs
3.6. Molecular Docking and Molecular Dynamics Simulation
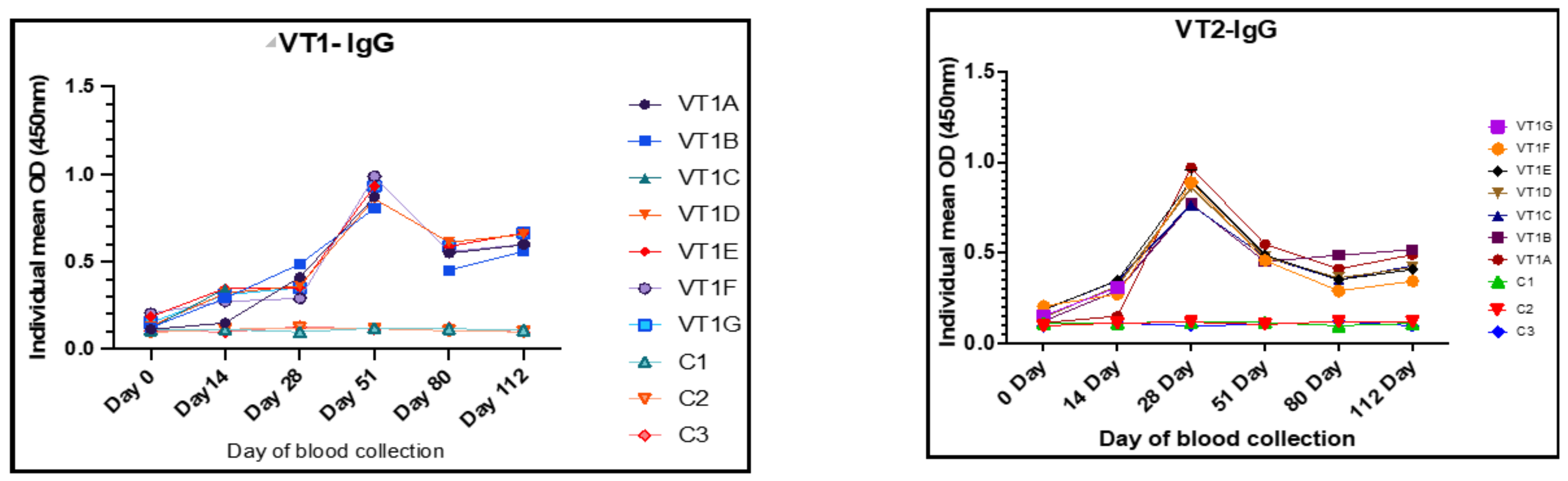
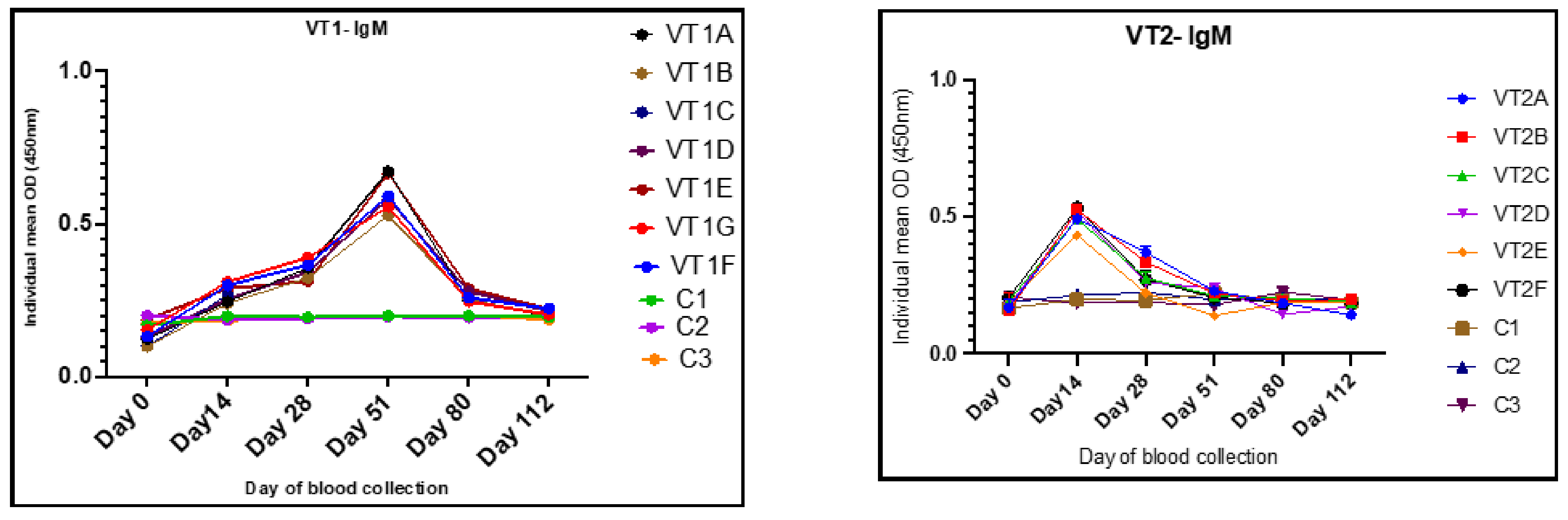
3.7. Dose Standardization on the Basis of IgG and IgM Responses
3.8. In Vivo Efficacy against Challenge Infestation
- Against larvae
- Against adults
3.9. Humoral Immune Responses to VT1 and VT2
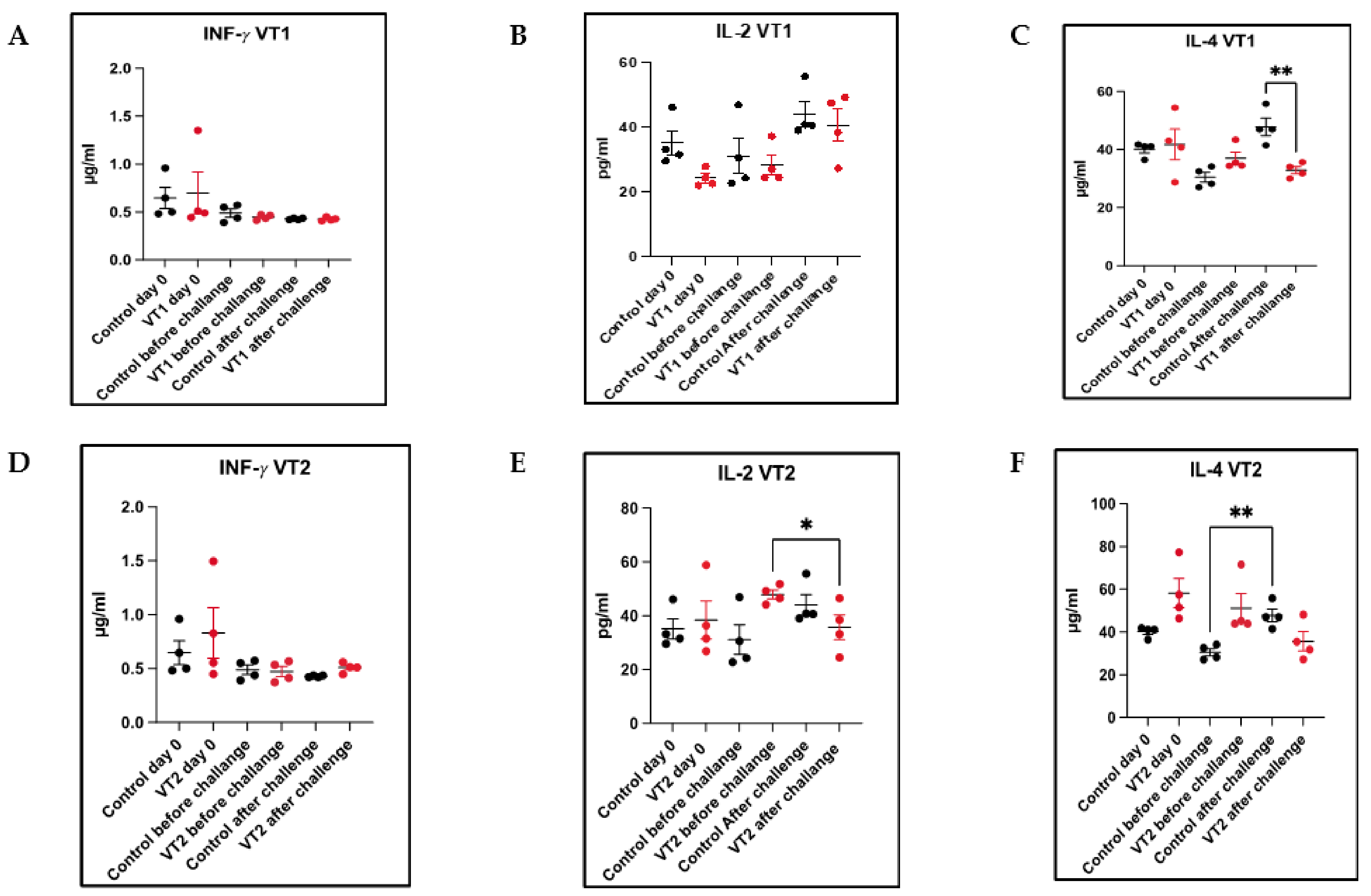
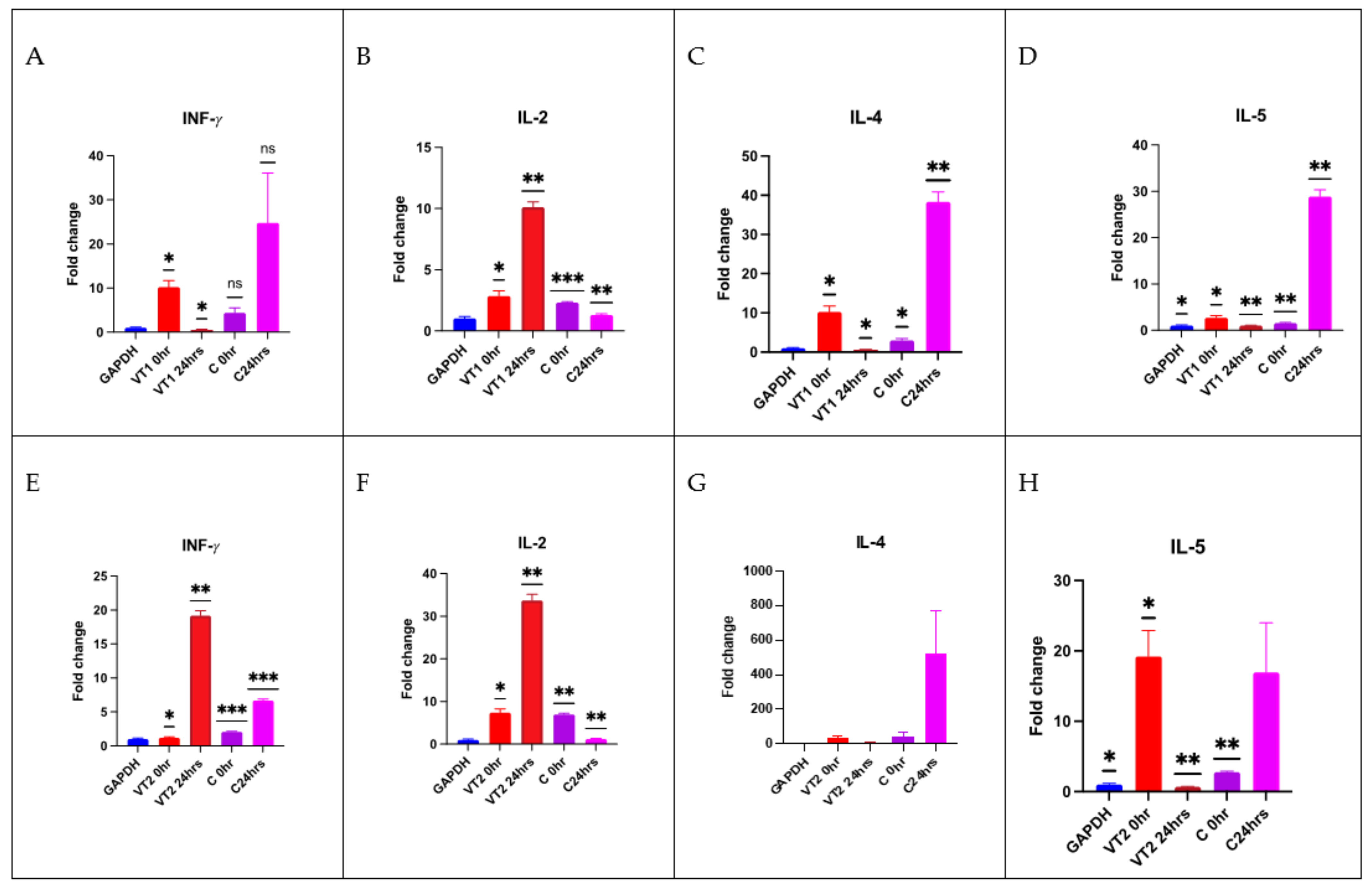
3.10. Cellular Immune Responses to VT1 and VT2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Otranto, D.; Brianti, E.; Latrofa, M.S.; Annoscia, G.; Weigl, S.; Lia, R.P.; Gaglio, G.; Napoli, E.; Giannetto, S.; Papadopoulos, E.; et al. On a Cercopithifilaria sp. transmitted by Rhipicephalus sanguineus: A neglected, but widespread Filarioid of dogs. Parasites Vectors 2012, 5, 1. [Google Scholar] [CrossRef]
- Wall, R.; Shearer, D. Veterinary Ectoparasites: Biology, Pathology and Control; Wiley: Hoboken, NJ, USA, 2001; ISBN 9780632056187. [Google Scholar]
- Lew-Tabor, A.E.; Rodriguez Valle, M. A review of reverse vaccinology approaches for the development of vaccines against ticks and tick borne diseases. Ticks Tick Borne Dis. 2016, 7, 573–585. [Google Scholar] [CrossRef]
- Singh, K.; Kumar, S.; Sharma, A.K.; Jacob, S.S.; RamVerma, M.; Singh, N.K.; Shakya, M.; Sankar, M.; Ghosh, S. Economic impact of predominant ticks and tick-borne diseases on Indian dairy production systems. Exp. Parasitol. 2022, 243, 108408. [Google Scholar] [CrossRef]
- Ghosh, S.; Azhahianambi, P.; Yadav, M.P. Upcoming and future strategies of tick control: Areview. J. Vector Borne Dis. 2007, 44, 79–89. [Google Scholar]
- Kumar, B.; Manjunathachar, H.V.; Ghosh, S. A review on Hyalomma species infestations on human and animals and progress on management strategies. Heliyon 2020, 6, e05675. [Google Scholar] [CrossRef]
- Ergönül, Ö. Crimean-Congo haemorrhagic fever. Lancet. Infect. Dis. 2006, 6, 203–214. [Google Scholar] [CrossRef]
- Ergonul, O.; Zeller, H.; Celikbas, A.; Dokuzoguz, B. The lack of Crimean-Congo hemorrhagic fever virus antibodies in healthcare workers in an endemic region. Int. J. Infect. Dis. 2007, 11, 48–51. [Google Scholar] [CrossRef]
- Yadav, P.D.; Patil, D.Y.; Shete, A.M.; Kokate, P.; Goyal, P.; Jadhav, S.; Sinha, S.; Zawar, D.; Sharma, S.K.; Kapil, A.; et al. Nosocomial infection of CCHF among health care workers in Rajasthan, India. BMC Infect. Dis. 2016, 16, 624. [Google Scholar] [CrossRef]
- Graf, J.F.; Gogolewski, R.; Leach-Bing, N.; Sabatini, G.A.; Molento, M.B.; Bordin, E.L.; Arantes, G.J. Tick control: An industry point of view. Parasitology 2004, 129, S427–S442. [Google Scholar] [CrossRef]
- Rodriguez-Vivas, R.I.; Hodgkinson, J.E.; Rosado-Aguilar, J.A.; Villegas-Perez, S.L.; Trees, A.J. The prevalence of pyrethroid pesistance phenotype and genotype in Rhipicephalus (Boophilus) microplus in Yucatan, Mexico. Vet. Parasitol. 2012, 184, 221–229. [Google Scholar] [CrossRef]
- Willadsen, P.; Jongejan, F. Immunology of the tick–host interaction and the control of ticks and tick-borne diseases. Parasitol. Today 1999, 15, 258–262. [Google Scholar] [CrossRef]
- De la Fuente, J.; Rodríguez, M.; Montero, C.; Redondo, M.; García-García, J.C.; Méndez, L.; Serrano, E.; Valdés, M.; Enríquez, A.; Canales, M.; et al. Vaccination against ticks (Boophilus spp.): The experience with the Bm86-based vaccine GavacTM. Genet. Anal. 1999, 15, 143–148. [Google Scholar] [CrossRef]
- De la Fuente, J.; Almazán, C.; Canales, M.; Pérez de la Lastra, J.M.; Kocan, K.M.; Willadsen, P. A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Anim. Health Res. Rev. 2007, 8, 23–28. [Google Scholar] [CrossRef]
- Ghosh, S.; Tiwari, S.S.; Srivastava, S.; Sharma, A.K.; Kumar, S.; Ray, D.D.; Rawat, A.K.S. Acaricidal properties of Ricinus communis leaf extracts against organophosphate and pyrethroids resistant Rhipicephalus (Boophilus) microplus. Vet. Parasitol. 2013, 192, 259–267. [Google Scholar] [CrossRef]
- Kemp, D.H.; Pearson, R.D.; Gough, J.M.; Willadsen, P. Vaccination against Boophilusmicroplus: Localization of antigens on tick gut cells and their interaction with the host immune system. Exp. Appl. Acarol. 1989, 7, 43–58. [Google Scholar] [CrossRef]
- De la Fuente, J.; Rodríguez, M.; Redondo, M.; Montero, C.; García-García, J.C.; Méndez, L.; Serrano, E.; Valdés, M.; Enriquez, A.; Canales, M.; et al. Field studies and cost-effectiveness analysis of vaccination with Gavac? against the cattle tick Boophilus microplus. Vaccine 1998, 16, 366–373. [Google Scholar] [CrossRef]
- De Vos, S.; Zeinstra, L.; Taoufik, A.; Willadsen, P.; Jongejan, F. Evidence for the utility from Boophilus microplus in vaccination against other tick species. Exp. Appl. Acarol. 2001, 25, 245–261. [Google Scholar] [CrossRef]
- Kumar, B.; Azhahianambi, P.; Ray, D.D.; Chaudhuri, P.; de la Fuente, J.; Kumar, R.; Ghosh, S. Comparative efficacy of rHaa86 and rBm86 against Hyalomma anatolicum anatolicum and Rhipicephalus (Boophilus) microplus. Parasite Immunol. 2012, 34, 297–301. [Google Scholar] [CrossRef]
- Willadsen, P.; Smith, D.; Cobon, G.; McKenna, R.V. Comparative vaccination of cattle against Boophilus microplus with recombinant antigen Bm86 alone or in combination with recombinant Bm91. Parasite Immunol. 1996, 18, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Azhahianambi, P.; Ray, D.D.; Chaudhuri, P.; Gupta, R.; Ghosh, S. Vaccine efficacy of Bm86 ortholog of Hyalomma aatolicum anatolicum, rHaa86 expressed in prokaryotic expression system. J. Parasitol. Res. 2009, 2009, 165812. [Google Scholar] [CrossRef]
- Canales, M.; Almazán, C.; Naranjo, V.; Jongejan, F.; de la Fuente, J. Vaccination with recombinant Boophilusannulatus Bm86 ortholog protein, Ba86, protects cattle against B. annulatus and B. microplus infestations. BMC Biotechnol. 2009, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Purcell, A.W.; McCluskey, J.; Rossjohn, J. More than one reason to rethink the use of peptides in vaccine design. Nat. Rev. Drug Discov. 2007, 6, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Malonis, R.J.; Lai, J.R.; Vergnolle, O. Peptide-based vaccines: Current progress and future challenges. Chem. Rev. 2020, 120, 3210–3229. [Google Scholar] [CrossRef]
- Patarroyo, J.H.; Portela, R.W.; de Castro, R.O.; Couto Pimentel, J.; Guzman, F.; Patarroyo, M.E.; Vargas, M.I.; Prates, A.A.; Dias Mendes, M.A. Immunization of cattle with synthetic peptides derived from the Boophilus microplus gut protein (Bm86). Vet. Immunol. Immunopathol. 2002, 88, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Mallon, A.; Fernández, E.; Encinosa, P.E.; Bello, Y.; Méndez-Pérez, L.; Ruiz, L.C.; Pérez, D.; González, M.; Garay, H.; Reyes, O.; et al. A novel tick antigen shows high vaccine efficacy against the dog tick, Rhipicephalus ssanguineus. Vaccine 2012, 30, 1782–1789. [Google Scholar] [CrossRef]
- Rodríguez-Mallon, A.; Encinosa, P.E.; Méndez-Pérez, L.; Bello, Y.; Rodríguez Fernández, R.; Garay, H.; Cabrales, A.; Méndez, L.; Borroto, C.; Estrada, M.P. High efficacy of a 20 amino acid peptide of the acidic ribosomal protein P0 against the cattle tick, Rhipicephalus microplus. Ticks Tick Borne Dis. 2015, 6, 530–537. [Google Scholar] [CrossRef]
- Rodríguez Mallón, A.; Javier González, L.; Encinosa Guzmán, P.E.; Bechara, G.H.; Sanches, G.S.; Pousa, S.; Cabrera, G.; Cabrales, A.; Garay, H.; Mejías, R.; et al. Functional and mass spectrometric evaluation of an anti-tick antigen based on the P0 peptide conjugated to Bm86 protein. Pathogens 2020, 9, 513. [Google Scholar] [CrossRef]
- Kumar, B.; Manjunathachar, H.V.; Nagar, G.; Ravikumar, G.; de la Fuente, J.; Saravanan, B.C.; Ghosh, S. Functional characterization of candidate antigens of Hyalomma anatolicum and evaluation of Its cross-protective efficacy against Rhipicephalus microplus. Vaccine 2017, 35, 5682–5692. [Google Scholar] [CrossRef]
- Manjunathachar, H.V.; Kumar, B.; Saravanan, B.C.; Choudhary, S.; Mohanty, A.K.; Nagar, G.; Chigure, G.; Ravi Kumar, G.V.P.P.S.; Fuente, J.; Ghosh, S. Identification and characterization of vaccine candidates against Hyalomma anatolicum —Vector of Crimean-Congo haemorrhagic fever virus. Transbound. Emerg. Dis. 2019, 66, 422–434. [Google Scholar] [CrossRef]
- Xavier, M.A.; Tirloni, L.; Pinto, A.F.M.; Diedrich, J.K.; Yates, J.R.; Gonzales, S.; Farber, M.; da Silva Vaz, I.; Termignoni, C. Tick gené’s organ engagement in lipid metabolism revealed by a combined transcriptomic and proteomic approach. Ticks Tick Borne Dis. 2019, 10, 787–797. [Google Scholar] [CrossRef]
- Ghosh, S.; Azhahianambi, P. Laboratory rearing of Theileria annulata-free Hyalomma anatolicum anatolicum ticks. Exp. Appl. Acarol. 2007, 43, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [PubMed]
- Almagro Armenteros, J.J.; Sønderby, C.K.; Sønderby, S.K.; Nielsen, H.; Winther, O. DeepLoc: Prediction of protein subcellular localization using deep learning. Bioinformatics 2017, 33, 4049. [Google Scholar] [CrossRef] [PubMed]
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: Improving sequence-based B-cell epitope prediction using conformational epitopes. Nucleic Acids Res. 2017, 45, W24–W29. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Raghava, G.P.S. Prediction of continuous B-cell epitopes in an antigen using recurrent neural network. Proteins: Struct. Funct. Bioinform. 2006, 65, 40–48. [Google Scholar] [CrossRef]
- Reynisson, B.; Alvarez, B.; Paul, S.; Peters, B.; Nielsen, M. NetMHCpan-4.1 and NetMHCIIpan-4.0: Improved predictions of MHC antigen presentation by concurrent motif deconvolution and integration of MS MHC eluted ligand data. Nucleic Acids Res. 2020, 48, W449–W454. [Google Scholar] [CrossRef]
- Dhanda, S.K.; Vir, P.; Raghava, G.P. Designing of interferon-gamma inducing MHC class-II binders. Biol. Direct 2013, 8, 30. [Google Scholar] [CrossRef]
- Dimitrov, I.; Bangov, I.; Flower, D.R.; Doytchinova, I. AllerTOP v.2—A server for in silico prediction of allergens. J. Mol. Model 2014, 20, 2278. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Källberg, M.; Margaryan, G.; Wang, S.; Ma, J.; Xu, J. RaptorX server: Aresource for template-based protein structure modeling. In Methods in Molecular Biology; Humana Press: New York, NY, USA, 2014; pp. 17–27. [Google Scholar]
- Heo, L.; Park, H.; Seok, C. GalaxyRefine: Protein structure refinement driven by side-chain repacking. Nucleic Acids Res. 2013, 41, W384–W388. [Google Scholar] [CrossRef]
- Wiederstein, M.; Sippl, M.J. ProSA-Web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef]
- Davis, I.W.; Leaver-Fay, A.; Chen, V.B.; Block, J.N.; Kapral, G.J.; Wang, X.; Murray, L.W.; Arendall, W.B.; Snoeyink, J.; Richardson, J.S.; et al. MolProbity: All-atom contacts and structure validation for proteins and nucleic acids. Nucleic Acids Res. 2007, 35, W375–W383. [Google Scholar] [CrossRef]
- De Vries, S.J.; Bonvin, A.M.J.J. CPORT: Aconsensus interface predictor and its performance in prediction-driven docking with HADDOCK. PLoS ONE 2011, 6, e17695. [Google Scholar] [CrossRef]
- Crowther, J.R. ELISA Guide Book; The Humana Press: Totowa, NJ, USA, 2000; Volume 149, ISBN 1-59259-049-7. [Google Scholar]
- Ghosh, S.; Singh, N.; Rawat, P. Comparative immunopotentiating roperties of saponin and incomplete freund’s adjuvant coupled to affinity purified larval antigen of Hyalomma anatolicum anatolicum. Indian J. Exp. Biol. 2002, 40, 1026–1031. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Willadsen, P. Antigen cocktails: Valid hypothesis or unsubstantiated hope? Trends Parasitol. 2008, 24, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Sajid, A.; Matias, J.; Arora, G.; Kurokawa, C.; DePonte, K.; Tang, X.; Lynn, G.; Wu, M.-J.; Pal, U.; Strank, N.O.; et al. mRNA vaccination induces tick resistance and prevents transmission of the Lyme disease agent. Sci. Transl. Med. 2021, 13, eabj9827. [Google Scholar] [CrossRef]
- Jelínková, L.; Jhun, H.; Eaton, A.; Petrovsky, N.; Zavala, F.; Chackerian, B. An Epitope-based malaria vaccine targeting the junctional region of circumsporozoite protein. NPJ Vaccines 2021, 6, 13. [Google Scholar] [CrossRef]
- Schoeler, G.B.; Wikel, S.K. Modulation of host immunity by haematophagous arthropods. Ann. Trop. Med. Parasitol. 2001, 95, 755–771. [Google Scholar] [CrossRef]
- Nuttall, P.; Labuda, M. Saliva-assisted transmission of tick-borne pathogens. In Ticks: Biology, Disease and Control; Bowman, A., Nuttall, P., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 205–219. [Google Scholar]
- Kazimírová, M.; Štibrániová, I. Tick salivary compounds: Their role in modulation of host defences and pathogen transmission. Front. Cell. Infect. Microbiol. 2013, 3, 43. [Google Scholar] [CrossRef]
- Chen, X.; Zaro, J.L.; Shen, W.C. Fusion protein linkers: Property, design and functionality. Adv. Drug Deliv. Rev. 2013, 65, 1357–1369. [Google Scholar] [CrossRef]
- Corradin, G.; Villard, V.; Kajava, A. Protein structure based strategies for antigen discovery and vaccine development against malaria and other pathogens. Endocr. Metab. Immune Disord. Drug Targets 2007, 7, 259–265. [Google Scholar] [CrossRef]
- Khatoon, N.; Pandey, R.K.; Prajapati, V.K. Exploring Leishmania secretory proteins to design B and T cell multi-epitope subunit vaccine using immunoinformatics approach. Sci. Rep. 2017, 7, 8285. [Google Scholar] [CrossRef]
- Atapour, A.; Vosough, P.; Jafari, S.; Sarab, G.A. A multi-epitope vaccine designed against blood-stage of malaria: An immunoinformatic and structural approach. Sci. Rep. 2022, 12, 11683. [Google Scholar] [CrossRef]
- Bell, J.K.; Mullen, G.E.D.; Leifer, C.A.; Mazzoni, A.; Davies, D.R.; Segal, D.M. Leucine-rich repeats and pathogen recognition in toll-like receptors. Trends Immunol. 2003, 24, 528–533. [Google Scholar] [CrossRef]
- Sá-Nunes, A.; Bafica, A.; Lucas, D.A.; Conrads, T.P.; Veenstra, T.D.; Andersen, J.F.; Mather, T.N.; Ribeiro, J.M.C.; Francischetti, I.M.B. Prostaglandin E2 Is a major inhibitor of dendritic cell maturation and function in Ixodes scapularis saliva. Cytokine 2007, 39, 37. [Google Scholar] [CrossRef]
- Oliveira, C.J.F.; Sá-Nunes, A.; Francischetti, I.M.B.; Carregaro, V.; Anatriello, E.; Silva, J.S.; de Miranda Santos, I.K.F.; Ribeiro, J.M.C.; Ferreira, B.R. Deconstructing tick saliva. J. Biol. Chem. 2011, 286, 10960–10969. [Google Scholar] [CrossRef]
- Rezaei, S.; Sefidbakht, Y.; Uskoković, V. Tracking the pipeline: Immunoinformatics and the COVID-19 vaccine design. Brief. Bioinform. 2021, 22, bbab241. [Google Scholar] [CrossRef]
- Pandey, R.K.; Bhatt, T.K.; Prajapati, V.K. Novel immunoinformatics approaches to design multi-epitope subunit vaccine for malaria by investigating anopheles salivary protein. Sci. Rep. 2018, 8, 1125. [Google Scholar] [CrossRef]
- Nevagi, R.J.; Toth, I.; Skwarczynski, M. Peptide-based vaccines. In Peptide Applications in Biomedicine, Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2018; pp. 327–358. [Google Scholar]
- Alexander, J.; del Guercio, M.-F.; Maewal, A.; Qiao, L.; Fikes, J.; Chesnut, R.W.; Paulson, J.; Bundle, D.R.; DeFrees, S.; Sette, A. Linear PADRE T helper epitope and carbohydrate B cell epitope conjugates induce specific high titer IgG antibody responses. J. Immunol. 2000, 164, 1625–1633. [Google Scholar] [CrossRef]
- Almazán, C.; Šimo, L.; Fourniol, L.; Rakotobe, S.; Borneres, J.; Cote, M.; Peltier, S.; Mayé, J.; Versillé, N.; Richardson, J.; et al. Multiple antigenic peptide-based vaccines targeting Ixodes ricinus neuropeptides induce a specific antibody response but do not impact tick infestation. Pathogens 2020, 9, 900. [Google Scholar] [CrossRef]
- Yang, J.; Luo, Y.; Shibu, M.A.; Toth, I.; Skwarczynskia, M. Cell-penetrating peptides: Efficient vectors for vaccine delivery. Curr. Drug Deliv. 2019, 16, 430–443. [Google Scholar] [CrossRef]
- Hajdusek, O.; Sojka, D.; Kopacek, P.; Buresova, V.; Franta, Z.; Sauman, I.; Winzerling, J.; Grubhoffer, L. Knockdown of proteins involved in iron metabolism limits tick reproduction and development. Proc. Natl. Acad. Sci. USA 2009, 106, 1033–1038. [Google Scholar] [CrossRef]
- Hajdusek, O.; Almazán, C.; Loosova, G.; Villar, M.; Canales, M.; Grubhoffer, L.; Kopacek, P.; de la Fuente, J. Characterization of Ferritin 2 for the control of tick infestations. Vaccine 2010, 28, 2993–2998. [Google Scholar] [CrossRef]
- Seixas, A.; Alzugaray, M.F.; Tirloni, L.; Parizi, L.F.; Pinto, A.F.M.; Githaka, N.W.; Konnai, S.; Ohashi, K.; Yates III, J.R.; Termignoni, C.; et al. Expression profile of Rhipicephalus microplus vitellogenin receptor during oogenesis. Ticks Tick Borne Dis. 2018, 9, 72–81. [Google Scholar] [CrossRef]
- Pedro, E.E.G.; Anayram, P.M.; Yamil, B.S.; Frank, L.L.B.; William, F.H.; Carlos, P.H.; Llilian, G.P.; Nemecio, G.; Mario, P.E.; Alina, R.-M. Comparison of montanide GEL 01 and oily montanideTM ISA 50 in presenting a peptide to the immune system of dogs. J. Vet. Med. Anim. Hlth. 2021, 13, 28–33. [Google Scholar] [CrossRef]
- Manning, J.E.; Oliveira, F.; Coutinho-Abreu, I.V.; Herbert, S.; Meneses, C.; Kamhawi, S.; Baus, H.A.; Han, A.; Czajkowski, L.; Rosas, L.A.; et al. Safety and immunogenicity of a mosquito saliva peptide-based vaccine: Arandomised, placebo-controlled, double-blind, phase 1 trial. Lancet 2020, 395, 1998–2007. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Schedule of Immunization | VT2 * # | VT1 * # | Control | ||
|---|---|---|---|---|---|
| Group 1 | Group 2 | Group 1 | Group 2 | ||
| 0 day | 50 µg in 100 µL Mili-QW + 150 µL adjuvant | 25 µg in 100 µL Mili-QW + 150 µL adjuvant | 50 µg in 100 µL Mili-QW + 150 µL adjuvant | 25 µg in 100 µL Mili-QW + 150 µL adjuvant | 100 µL Mili-QW + 150 µL adjuvant |
| 14 day | 25 µg in 100 µL Mili-QW + 150 µL adjuvant | 12.5 µg in 100 µL Mili-QW + 150 µL adjuvant | 25 µg in 100 µL Mili-QW + 150 µL adjuvant | 12.5 µg in 100 µL Mili-QW + 150 µL adjuvant | Same dose |
| 28 day | Same as booster dose | Same as booster dose | Same as booster dose | Same as booster dose | Same dose |
| For larvae | DT (%) = 100 (1 − NTV/NTC) | Where DT% is the percentage rejection in the number of nymphs; NTV, the number of nymphs dropped from the immunized rabbits; and NTC, the number of nymphs dropped from thecontrol rabbits. |
| MO (%) = 100 (1 − MLI/MLC) | Where MO (%) is the percent reduction in moulting of engorged nymphs; MLI and MLC, the number of engorged nymphs moulted to adults from immunized and control rabbits, respectively. | |
| E (%) = 100 [1 − (CRT × CRM)] | Where E (%) is the percentage efficacy of antigen against larvae; CRT is reduction in the number of nymphs (NTV/NTC); CRM is reduction in number of engorged nymphs moulted to adults (MLI/MLC). | |
| For adults | DT% = 100(1 − NTV/NTC) | Where DT% is the percentage rejection in the number of females; NTV, the number of females dropped from the immunized animals; NTC, the number of females dropped from the control animals. |
| DR (%) = 100(1 − PMTV/PMTC) | Where DR (%) is the percentage reduction of mean weight of adult females; PMTV is the mean weight of adult females dropped from the immunized animals; PMTC is the mean weight of adult females dropped from the control animals. | |
| DO (%) = 100(1 − PATV/PATC) | Where DO (%) is the percentage reduction of mean weight of eggs; PATV is the mean weight of eggs of females which fed on vaccinated animals; PATC is the mean weight of eggs of females which fed on control animals. | |
| RI = Egg weight/engorge tick weight | Where RI is designated as reproductive index. | |
| RF (%) = 100(1 − RIV/RIC) | Where RF (%) is the percentage reduction in adult fertility; RIV is the mean RI of adult females dropped from the immunized animals; RIC is the mean RI of adult females dropped from the control animals. | |
| E (%) = 100 [1 − (CRT × CRO × CRI)] | Where E% is the percentage efficacy of antigens; CRO is reduction in egg laying capacity (PATV/PATC); CRT is the reduction in the number of adult females (NTV/NTC); CRI is the reduction in tick fertility. |
| Protein | CTL Epitopes | IFN-γ Epitope | HTL Epitopes | IFN-γEpitope | B-Cell Epitopes | IFN-γEpitope |
|---|---|---|---|---|---|---|
| VgR | GVHVYHPVL (522) | Yes | YGEPFLLYMLPNQIR (1301) | No | ||
| ALFEDWLYW (1514) | Yes | |||||
| ALLVLGYVL (1682) | No | LCVALLVLGYVLYRR (1680) | No | |||
| FER2 | INLELHASL (42) | No | NLELHASLVYMQMAA (43) | No | DFLEQEFLAEQVKSID (111) | Yes |
| DDDPQMADFLEQEFL (141) | No | |||||
| DTGLGEFLLDQQLRT (176) | No | |||||
| LNAIPVSPQTNLFYS (33) | No | |||||
| TPM | RMDGLEGQL (140) | No | ELRVVGNNLKSLEVS (196) | No | LEEELRVVGNNLKSL (193) | No |
| KIVELEEEL (189) | Yes |
| IgG | IgM | |||||
|---|---|---|---|---|---|---|
| Dose | 50 µg | 100 µg | Control | 50 µg | 100 µg | Control |
| Day 0 | 0.11 ± 0.00 ** | 0.12 ± 0.01 | 0.12 ± 0.00 | 0.11 ± 0.00 | 0.13 ± 0.00 | 0.16 ± 0.03 |
| Day 14 | 0.151 ± 0.01 ** | 0.23 ± 0.014 | 0.13 ± 0.00 | 0.32 ± 0.00 | 0.29 ± 0.00 | 0.18 ± 0.02 |
| Day 28 | 0.412 ± 0.00 ** | 0.31 ± 0.01 | 0.13 ± 0.00 | 0.40 ± 0.00 | 0.40 ± 0.00 | 0.17 ± 0.00 |
| Day 42 | 0.87 ± 0.01 ** | 0.57 ± 0.01 | 0.11 ± 0.00 | 0.74± 0.01 | 0.57 ± 0.00 | 0.17 ± 0.00 |
| IgG | IgM | |||||
|---|---|---|---|---|---|---|
| Dose | 50 µg | 100 µg | Control | 50 µg | 100 µg | Control |
| Day 0 | 0.12 ± 0.01 * | 0.11 ± 0.00 | 0.11 ± 0.00 | 0.17 ±0.03 * | 0.12 ± 0.01 * | 0.17 ± 0.01 |
| Day 14 | 0.15 ± 0.01 * | 0.29 ± 0.00 | 0.11 ± 0.00 | 0.38 ± 0.03 * | 0.53 ± 0.02 * | 0.2 ± 0.00 |
| Day 28 | 0.47 ± 0.01 * | 0.97 ± 0.01 | 0.12 ± 0.01 | 0.27 ± 0.02 * | 0.34 ± 0.01 * | 0.19 ± 0.00 |
| Day 42 | 0.45 ± 0.01 * | 0.85 ± 0.01 | 0.12 ± 0.02 | 0.21 ±0.01 * | 0.25 ± 0.03 * | 0.2 ± 0.01 |
| Group | Mean wt (mg)/Nymph | No. of Nymphs (Mean ± SE) | DT% | MO% | E% |
|---|---|---|---|---|---|
| VT1 | 7.67 ± 0.0 | 72.75 ± 13.5 * | 67.9 | 79.3 | 93.3 |
| VT2 | 7.11 ± 0.0 | 41 ± 2.6 *** | 81.9 | 83.0 | 96.9 |
| Control | 8.13 ± 0.0 | 226.7 ± 12.4 *** |
| Group | No. of Females Dropped (Mean ± SE) | wt (mg)/Tick (Mean ± SE) | Egg Masses (Mean ± SE) | DT% | DR% | DO% | RF% | E% |
|---|---|---|---|---|---|---|---|---|
| VT1 | 8 ± 0.4 *** | 196.7± 6.8 *** | 57.0 ± 6.7 *** | 43.7 | 44.4 | 68.5 | 42.7 | 89.9 |
| VT2 | 8 ± 0.4 *** | 202.2± 4.7 *** | 82.2 ± 6.5 *** | 43.7 | 43.0 | 54.2 | 47.5 | 86.4 |
| Control | 14.2 ± 0.4 | 357.3 ± 4.3 | 181.4± 3.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nandi, A.; Manisha; Solanki, V.; Tiwari, V.; Sajjanar, B.; Sankar, M.; Saini, M.; Shrivastava, S.; Bhure, S.K.; Ghosh, S. Protective Efficacy of Multiple Epitope-Based Vaccine against Hyalomma anatolicum, Vector of Theileria annulata and Crimean–Congo Hemorrhagic Fever Virus. Vaccines 2023, 11, 881. https://doi.org/10.3390/vaccines11040881
Nandi A, Manisha, Solanki V, Tiwari V, Sajjanar B, Sankar M, Saini M, Shrivastava S, Bhure SK, Ghosh S. Protective Efficacy of Multiple Epitope-Based Vaccine against Hyalomma anatolicum, Vector of Theileria annulata and Crimean–Congo Hemorrhagic Fever Virus. Vaccines. 2023; 11(4):881. https://doi.org/10.3390/vaccines11040881
Chicago/Turabian StyleNandi, Abhijit, Manisha, Vandana Solanki, Vishvanath Tiwari, Basavaraj Sajjanar, Muthu Sankar, Mohini Saini, Sameer Shrivastava, S. K. Bhure, and Srikant Ghosh. 2023. "Protective Efficacy of Multiple Epitope-Based Vaccine against Hyalomma anatolicum, Vector of Theileria annulata and Crimean–Congo Hemorrhagic Fever Virus" Vaccines 11, no. 4: 881. https://doi.org/10.3390/vaccines11040881