

Efficacy of Two Vaccination Strategies against Infectious Bronchitis in Laying Hens
 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Layer Chickens
2.3. IBV Challenge Strain and Vaccines
2.4. Experimental Design
2.5. Scoring of Clinical Signs and Measuring of Egg Parameters
2.6. Monitoring of Serum Anti-IBV Antibodies
2.7. Flow Cytometry Technique
2.8. RNA Extraction and IBV Genome Load Quantification
2.9. Histopathology
2.10. Data Analysis
3. Results
3.1. Clinical Signs
3.2. Egg Production and Egg Quality Parameters
3.3. Antibody-Mediated Immune Response
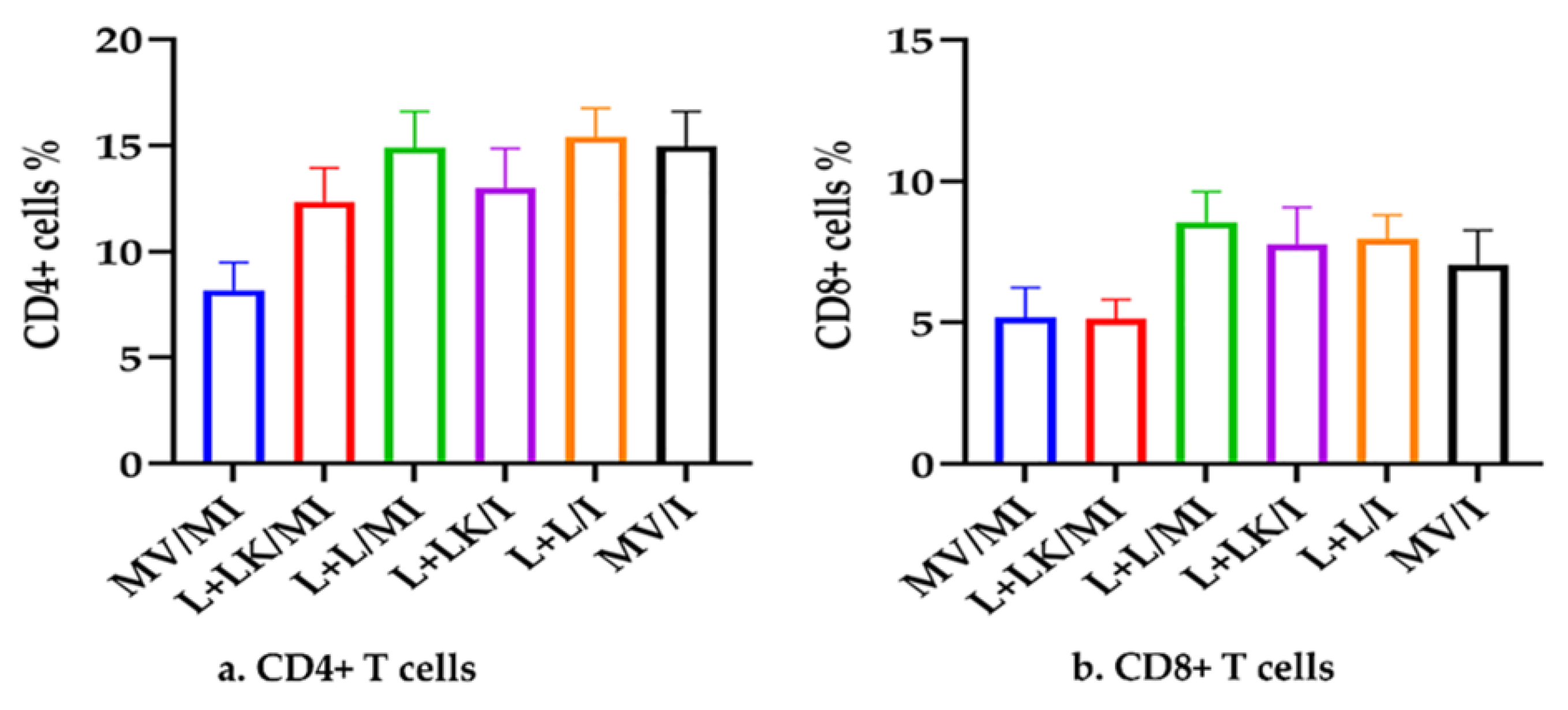
3.4. Peripheral Blood T-Cell Subsets (CD4 + and CD8 + T Cells)
3.5. IBV Genome Loads
3.5.1. IBV Genome Loads in Oropharyngeal and Cloacal Swabs
3.5.2. IBV Genome Loads in Tissues
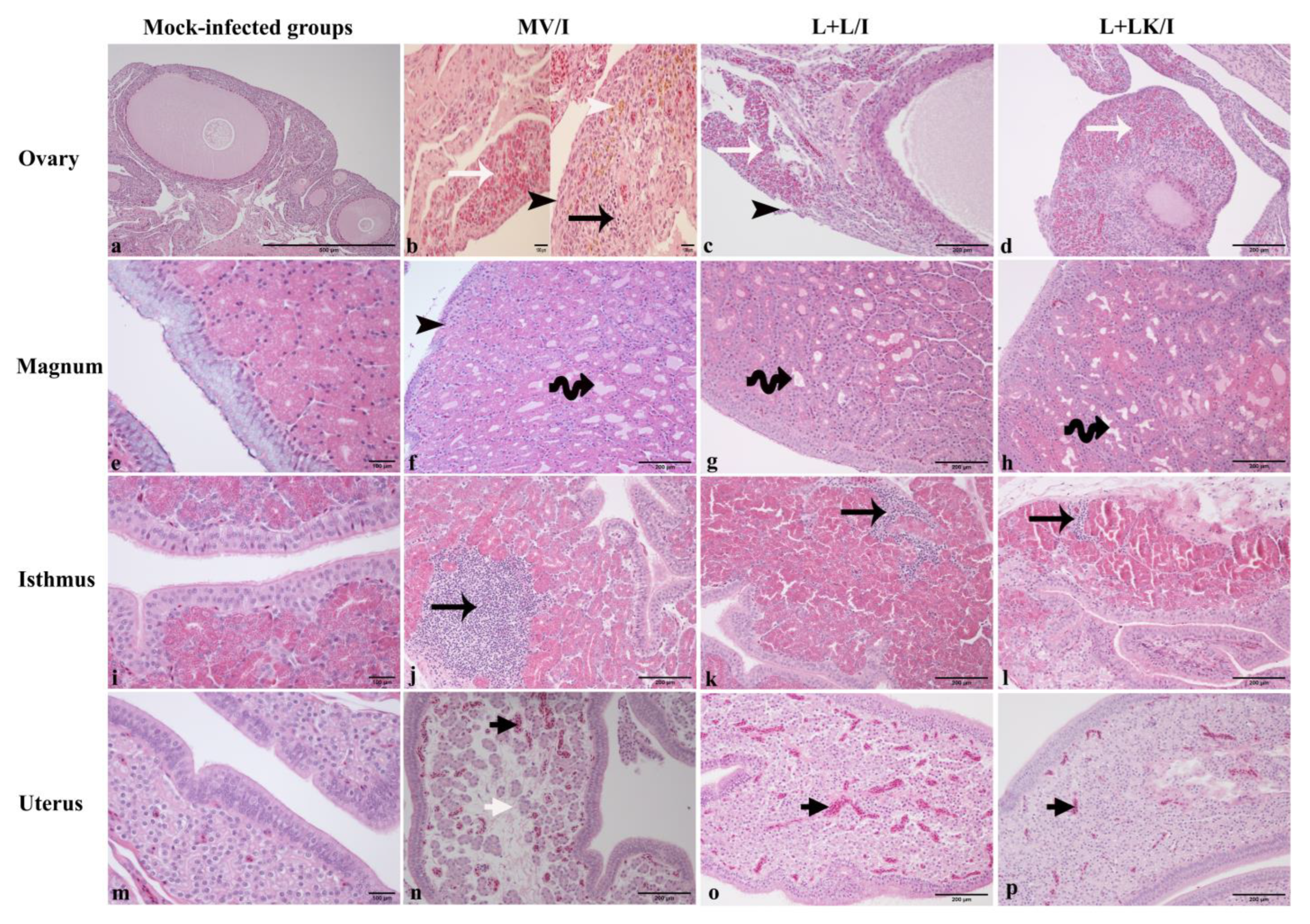
3.6. Histopathological Findings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cavanagh, D. Coronaviruses in poultry and other birds. Avian Pathol. 2005, 34, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef] [PubMed]
- França, M.; Woolcock, P.R.; Yu, M.; Jackwood, M.W.; Shivaprasad, H.L. Nephritis associated with infectious bronchitis virus Cal99 variant in game chickens. Avian Dis. 2011, 55, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Boroomand, Z.; Asasi, K.; Mohammadi, A. Pathogenesis and tissue distribution of avian infectious bronchitis virus isolate IRFIBV32 (793/B serotype) in experimentally infected broiler chickens. Sci. World J. 2012, 2012, 402537. [Google Scholar] [CrossRef]
- Liu, S.; Chen, J.; Han, Z.; Zhang, Q.; Shao, Y.; Kong, X.; Tong, G. Infectious bronchitis virus: S1 gene characteristics of vaccines used in China and efficacy of vaccination against heterologous strains from China. Avian Pathol. 2006, 35, 394–399. [Google Scholar] [CrossRef]
- Ambali, A.G.; Jones, R.C. Early pathogenesis in chicks of infection with an enterotropic strain of infectious bronchitis virus. Avian Dis. 1990, 34, 809–817. [Google Scholar] [CrossRef]
- MacDonald, J.W.; McMartin, D.A. Observations on the effects of the H52 and H120 vaccine strains of infectious bronchitis virus in the domestic fowl. Avian Pathol. 1976, 5, 157–173. [Google Scholar] [CrossRef]
- Sevoian, M.; Levine, P. Effects of Infectious Bronchitis on the Reproductive Tracts, Egg Production, and Egg Quality of Laying Chickens. Avian Dis. 1957, 1, 136. [Google Scholar] [CrossRef]
- Raj, G.D.; Jones, R.C. Infectious bronchitis virus: Immunopathogenesis of infection in the chicken. Avian Pathol. 1997, 26, 677–706. [Google Scholar] [CrossRef]
- Bande, F.; Arshad, S.S.; Omar, A.R.; Hair-Bejo, M.; Mahmuda, A.; Nair, V. Global distributions and strain diversity of avian infectious bronchitis virus: A review. Anim. Health Res. Rev. 2017, 18, 70–83. [Google Scholar] [CrossRef] [Green Version]
- Moreno, A.; Franzo, G.; Massi, P.; Tosi, G.; Blanco, A.; Antilles, N.; Biarnes, M.; Majó, N.; Nofrarías, M.; Dolz, R.; et al. A novel variant of the infectious bronchitis virus resulting from recombination events in Italy and Spain. Avian Pathol. 2017, 46, 28–35. [Google Scholar] [CrossRef] [PubMed]
- de Wit, J.J.; de Jong, M.C.; Pijpers, A.; Verheijden, J.H. Transmission of infectious bronchitis virus within vaccinated and unvaccinated groups of chickens. Avian Pathol. 1998, 27, 464–471. [Google Scholar] [CrossRef]
- Cook, J.K.A. Coronaviridae. In Poultry Diseases, 6th ed.; Pattison, P.F.M.M., Alexander, J.M.B.D.J., Eds.; Saunders Elsevier: Philadelphia, PA, USA, 2008; pp. 340–349. [Google Scholar]
- Jordan, B. Vaccination against infectious bronchitis virus: A continuous challenge. Vet. Microbiol. 2017, 206, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Roh, H.J.; Hilt, D.A.; Williams, S.M.; Jackwooda, M.W. Evaluation of infectious bronchitis virus Arkansas-type vaccine failure in commercial broilers. Avian Dis. 2013, 57, 248–259. [Google Scholar] [CrossRef] [PubMed]
- De Wit, J.J.; Swart, W.A.; Fabri, T.H. Efficacy of infectious bronchitis virus vaccinations in the field: Association between the alpha-IBV IgM response, protection and vaccine application parameters. Avian Pathol. 2010, 39, 123–131. [Google Scholar] [CrossRef]
- Chhabra, R.; Forrester, A.; Lemiere, S.; Awad, F.; Chantrey, J.; Ganapathy, K. Mucosal, Cellular, and Humoral Immune Responses Induced by Different Live Infectious Bronchitis Virus Vaccination Regimes and Protection Conferred against Infectious Bronchitis Virus Q1 Strain. Clin. Vaccine Immunol. 2015, 22, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Okino, C.H.; Alessi, A.C.; Montassier Mde, F.; Rosa, A.J.; Wang, X.; Montassier, H.J. Humoral and cell-mediated immune responses to different doses of attenuated vaccine against avian infectious bronchitis virus. Viral Immunol. 2013, 26, 259–267. [Google Scholar] [CrossRef]
- Bande, F.; Arshad, S.S.; Bejo, M.H.; Moeini, H.; Omar, A.R. Progress and challenges toward the development of vaccines against avian infectious bronchitis. J. Immunol. Res. 2015, 2015, 424860. [Google Scholar] [CrossRef]
- De Wit, J.J.; Boelm, G.J.; van Gerwe, T.J.; Swart, W.A. The required sample size in vaccination-challenge experiments with infectious bronchitis virus, a meta-analysis. Avian Pathol. 2013, 42, 9–16. [Google Scholar] [CrossRef]
- Awad, F.; Forrester, A.; Baylis, M.; Lemiere, S.; Ganapathy, K.; Hussien, H.A.; Capua, I. Protection conferred by live infectious bronchitis vaccine viruses against variant Middle East IS/885/00-like and IS/1494/06-like isolates in commercial broiler chicks. Vet. Rec. Open 2015, 2, e000111. [Google Scholar] [CrossRef] [Green Version]
- Bru, T.; Vila, R.; Cabana, M.; Geerligs, H.J. Protection of chickens vaccinated with combinations of commercial live infectious bronchitis vaccines containing Massachusetts, Dutch and QX-like serotypes against challenge with virulent infectious bronchitis viruses 793B and IS/1494/06 Israel variant 2. Avian Pathol. 2017, 46, 52–58. [Google Scholar] [CrossRef]
- De Wit, J.J.; Cook, J.K. Factors influencing the outcome of infectious bronchitis vaccination and challenge experiments. Avian Pathol. 2014, 43, 485–497. [Google Scholar] [CrossRef]
- Box, P.G.; Beresford, A.V.; Roberts, B. Protection of laying hens against infectious bronchitis with inactivated emulsion vaccines. Vet. Rec. 1980, 106, 264–268. [Google Scholar] [CrossRef]
- Box, P.G.; Ellis, K.R. Infectious bronchitis in laying hens: Interference with response to emulsion vaccine by attenuated live vaccine. Avian Pathol. 1985, 14, 9–22. [Google Scholar] [CrossRef]
- Box, P.G.; Holmes, H.C.; Finney, P.M.; Froymann, R. Infectious bronchitis in laying hens: The relationship between haemagglutination inhibition antibody levels and resistance to experimental challenge. Avian Pathol. 1988, 17, 349–361. [Google Scholar] [CrossRef]
- Muneer, M.A.; Newman, J.A.; Halvorson, D.A.; Sivanandan, V.; Coon, C.N. Effects of avian infectious bronchitis virus (Arkansas strain) on vaccinated laying chickens. Avian Dis. 1987, 31, 820–828. [Google Scholar] [CrossRef]
- De Wit, J.J.S.; Malo, A.; Cook, J.K.A. Induction of IBV strain-specific neutralizing antibodies and broad spectrum protection in layer pullets primed with IBV Massachusetts (Mass) and 793B vaccines prior to injection of inactivated vaccine containing Mass antigen. Avian Pathol. 2019, 48, 135–147. [Google Scholar] [CrossRef]
- De Wit, J.J.; Nieuwenhuisen-van Wilgen, J.; Hoogkamer, A.; van de Sande, H.; Zuidam, G.J.; Fabri, T.H. Induction of cystic oviducts and protection against early challenge with infectious bronchitis virus serotype D388 (genotype QX) by maternally derived antibodies and by early vaccination. Avian Pathol. 2011, 40, 463–471. [Google Scholar] [CrossRef]
- Terregino, C.; Toffan, A.; Beato, M.S.; De Nardi, R.; Vascellari, M.; Meini, A.; Ortali, G.; Mancin, M.; Capua, I. Pathogenicity of a QX strain of infectious bronchitis virus in specific pathogen free and commercial broiler chickens, and evaluation of protection induced by a vaccination programme based on the Ma5 and 4/91 serotypes. Avian Pathol. 2008, 37, 487–493. [Google Scholar] [CrossRef]
- Hassan, M.S.H.; Buharideen, S.M.; Ali, A.; Najimudeen, S.M.; Goldsmith, D.; Coffin, C.S.; Cork, S.C.; van der Meer, F.; Abdul-Careem, M.F. Efficacy of Commercial Infectious Bronchitis Vaccines against Canadian Delmarva (DMV/1639) Infectious Bronchitis Virus Infection in Layers. Vaccines 2022, 10, 1194. [Google Scholar] [CrossRef]
- Pulendran, B.; Ahmed, R. Immunological mechanisms of vaccination. Nat. Immunol. 2011, 12, 509–517. [Google Scholar] [CrossRef]
- Hassan, M.S.H.; Ojkic, D.; Coffin, C.S.; Cork, S.C.; van der Meer, F.; Abdul-Careem, M.F. Delmarva (DMV/1639) Infectious Bronchitis Virus (IBV) Variants Isolated in Eastern Canada Show Evidence of Recombination. Viruses 2019, 11, 1054. [Google Scholar] [CrossRef]
- Martin, E.A.; Brash, M.L.; Hoyland, S.K.; Coventry, J.M.; Sandrock, C.; Guerin, M.T.; Ojkic, D. Genotyping of infectious bronchitis viruses identified in Canada between 2000 and 2013. Avian Pathol. 2014, 43, 264–268. [Google Scholar] [CrossRef]
- Amarasinghe, A.; Popowich, S.; De Silva Senapathi, U.; Abdul-Cader, M.S.; Marshall, F.; van der Meer, F.; Cork, S.C.; Gomis, S.; Abdul-Careem, M.F. Shell-Less Egg Syndrome (SES) Widespread in Western Canadian Layer Operations Is Linked to a Massachusetts (Mass) Type Infectious Bronchitis Virus (IBV) Isolate. Viruses 2018, 10, 437. [Google Scholar] [CrossRef]
- Buharideen, S.M.; Hassan, M.S.H.; Najimudeen, S.M.; Niu, D.; Czub, M.; Gomis, S.; Abdul-Careem, M.F. Immune Responses in Laying Hens after an Infectious Bronchitis Vaccination of Pullets: A Comparison of Two Vaccination Strategies. Vaccines 2021, 9, 531. [Google Scholar] [CrossRef]
- He, L.; Martins, P.; Huguenin, J.; Van, T.N.; Manso, T.; Galindo, T.; Gregoire, F.; Catherinot, L.; Molina, F.; Espeut, J. Simple, sensitive and robust chicken specific sexing assays, compliant with large scale analysis. PLoS ONE 2019, 14, e0213033. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty per Cent Endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- De Silva Senapathi, U.; Abdul-Cader, M.S.; Amarasinghe, A.; van Marle, G.; Czub, M.; Gomis, S.; Abdul-Careem, M.F. The In Ovo Delivery of CpG Oligonucleotides Protects against Infectious Bronchitis with the Recruitment of Immune Cells into the Respiratory Tract of Chickens. Viruses 2018, 10, 635. [Google Scholar] [CrossRef]
- Anderson, K.E.; Tharrington, J.B.; Curtis, P.A. Shell Characteristics of Eggs from Historic Strains of Single Comb White Leghorn Chickens and the Relationships of Egg Shape to Shell Strength. Int. J. Poult. Sci. 2004, 3, 17–19. [Google Scholar] [CrossRef]
- Monira, K.N.; Salahuddin, M.; Miah, G. Effect of Breed and Holding Period on Egg Quality Characteristics of Chicken. Int. J. Poult. Sci. 2003, 2, 261–263. [Google Scholar] [CrossRef] [Green Version]
- Barboza-Solis, C.; Najimudeen, S.M.; Perez-Contreras, A.; Ali, A.; Joseph, T.; King, R.; Ravi, M.; Peters, D.; Fonseca, K.; Gagnon, C.A.; et al. Evaluation of Recombinant Herpesvirus of Turkey Laryngotracheitis (rHVT-LT) Vaccine against Genotype VI Canadian Wild-Type Infectious Laryngotracheitis Virus (ILTV) Infection. Vaccines 2021, 9, 1425. [Google Scholar] [CrossRef]
- Kameka, A.M.; Haddadi, S.; Kim, D.S.; Cork, S.C.; Abdul-Careem, M.F. Induction of innate immune response following infectious bronchitis corona virus infection in the respiratory tract of chickens. Virology 2014, 450–451, 114–121. [Google Scholar] [CrossRef]
- Benyeda, Z.; Szeredi, L.; Mató, T.; Süveges, T.; Balka, G.; Abonyi-Tóth, Z.; Rusvai, M.; Palya, V. Comparative histopathology and immunohistochemistry of QX-like, Massachusetts and 793/B serotypes of infectious bronchitis virus infection in chickens. J. Comp. Pathol. 2010, 143, 276–283. [Google Scholar] [CrossRef]
- Jackwood, M.W.; Jordan, B.J.; Roh, H.J.; Hilt, D.A.; Williams, S.M. Evaluating Protection against Infectious Bronchitis Virus by Clinical Signs, Ciliostasis, Challenge Virus Detection, and Histopathology. Avian Dis. 2015, 59, 368–374. [Google Scholar] [CrossRef]
- Sjaak de Wit, J.J.; Ter Veen, C.; Koopman, H.C.R. Effect of IBV D1466 on egg production and egg quality and the effect of heterologous priming to increase the efficacy of an inactivated IBV vaccine. Avian Pathol. 2020, 49, 185–192. [Google Scholar] [CrossRef]
- Coffman, R.L.; Sher, A.; Seder, R.A. Vaccine adjuvants: Putting innate immunity to work. Immunity 2010, 33, 492–503. [Google Scholar] [CrossRef]
- Yan, S.; Zhao, J.; Xie, D.; Huang, X.; Cheng, J.; Guo, Y.; Liu, C.; Ma, Z.; Yang, H.; Zhang, G. Attenuation, safety, and efficacy of a QX-like infectious bronchitis virus serotype vaccine. Vaccine 2018, 36, 1880–1886. [Google Scholar] [CrossRef]
- Chousalkar, K.K.; Roberts, J.R. Effects of Australian strains of infectious bronchitis virus on internal and external quality of hen eggs. Anim. Prod. Sci. 2009, 49, 162–169. [Google Scholar] [CrossRef]
- Butler, E.J.; Curtis, M.J.; Pearson, A.W.; McDougall, J.S. Effect of infectious bronchitis on the structure and composition of egg albumen. J. Sci. Food Agric. 1972, 23, 359–369. [Google Scholar] [CrossRef]
- Cook, J.K.; Davison, T.F.; Huggins, M.B.; McLaughlan, P. Effect of in ovo bursectomy on the course of an infectious bronchitis virus infection in line C White Leghorn chickens. Arch. Virol. 1991, 118, 225–234. [Google Scholar] [CrossRef]
- Cook, J.K.; Huggins, M.B.; Ellis, M.M. Use of an infectious bronchitis virus and Escherichia coli model infection to assess the ability to vaccinate successfully against infectious bronchitis in the presence of maternally-derived immunity. Avian Pathol. 1991, 20, 619–626. [Google Scholar] [CrossRef]
- Ignjatovic, J.; McWaters, P.G. Monoclonal antibodies to three structural proteins of avian infectious bronchitis virus: Characterization of epitopes and antigenic differentiation of Australian strains. J. Gen. Virol. 1991, 72, 2915–2922. [Google Scholar] [CrossRef]
- Raggi, L.G.; Lee, G.G. Lack of Correlation between Infectivity, Serologic Response and Challenge Results in Immunization with an Avian Infectious Bronchitis Vaccine. J. Immunol. 1965, 94, 538–543. [Google Scholar] [CrossRef]
- Collisson, E.W.; Pei, J.; Dzielawa, J.; Seo, S.H. Cytotoxic T lymphocytes are critical in the control of infectious bronchitis virus in poultry. Dev. Comp. Immunol. 2000, 24, 187–200. [Google Scholar] [CrossRef]
- Pei, J.; Sekellick, M.J.; Marcus, P.I.; Choi, I.S.; Collisson, E.W. Chicken interferon type I inhibits infectious bronchitis virus replication and associated respiratory illness. J. Interferon Cytokine Res. 2001, 21, 1071–1077. [Google Scholar] [CrossRef]
- Ahmed, R.; Butler, L.D.; Bhatti, L. T4+ T helper cell function in vivo: Differential requirement for induction of antiviral cytotoxic T-cell and antibody responses. J. Virol. 1988, 62, 2102–2106. [Google Scholar] [CrossRef]
- Liu, G.; Wang, Q.; Liu, N.; Xiao, Y.; Tong, T.; Liu, S.; Wu, D. Infectious bronchitis virus nucleoprotein specific CTL response is generated prior to serum IgG. Vet. Immunol. Immunopathol. 2012, 148, 353–358. [Google Scholar] [CrossRef]
- Pei, J.; Briles, W.E.; Collisson, E.W. Memory T cells protect chicks from acute infectious bronchitis virus infection. Virology 2003, 306, 376–384. [Google Scholar] [CrossRef]
- Seo, S.H.; Collisson, E.W. Specific cytotoxic T lymphocytes are involved in in vivo clearance of infectious bronchitis virus. J. Virol. 1997, 71, 5173–5177. [Google Scholar] [CrossRef]
- Chousalkar, K.K.; Roberts, J.R.; Reece, R. Comparative histopathology of two serotypes of infectious bronchitis virus (T and n1/88) in laying hens and cockerels. Poult. Sci. 2007, 86, 50–58. [Google Scholar] [CrossRef]
- Pereira, N.A.; Alessi, A.C.; Montassier, H.J.; Pereira, R.J.G.; Taniwaki, S.A.; Botosso, V.F.; Rui, B.R.; Richtzenhain, L.J. Gonadal pathogenicity of an infectious bronchitis virus strain from the Massachusetts genotype. Braz. J. Microbiol. 2019, 50, 313–320. [Google Scholar] [CrossRef]
- Ghani, S.; Feuerer, M.; Doebis, C.; Lauer, U.; Loddenkemper, C.; Huehn, J.; Hamann, A.; Syrbe, U. T cells as pioneers: Antigen-specific T cells condition inflamed sites for high-rate antigen-non-specific effector cell recruitment. Immunology 2009, 128, e870–e880. [Google Scholar] [CrossRef]
- Rodenberg, J.; Sharma, J.M.; Belzer, S.W.; Nordgren, R.M.; Naqi, S. Flow cytometric analysis of B cell and T cell subpopulations in specific-pathogen-free chickens infected with infectious bursal disease virus. Avian Dis. 1994, 38, 16–21. [Google Scholar] [CrossRef]
- Klein, D. Quantification using real-time PCR technology: Applications and limitations. Trends Mol. Med. 2002, 8, 257–260. [Google Scholar] [CrossRef]
- De Wit, J.J. Detection of infectious bronchitis virus. Avian Pathol. 2000, 29, 71–93. [Google Scholar] [CrossRef]
- Roh, H.J.; Hilt, D.A.; Jackwood, M.W. Detection of infectious bronchitis virus with the use of real-time quantitative reverse transcriptase-PCR and correlation with virus detection in embryonated eggs. Avian Dis. 2014, 58, 398–403. [Google Scholar] [CrossRef]
- Raj, G.D.; Jones, R.C. Effect of T-cell suppression by cyclosporin on primary and persistent infections of infectious bronchitis virus in chickens. Avian Pathol. 1997, 26, 257–276. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccination/Challenge | MV (n = 14) | L+LK (n = 18) | L+L (n = 18) | |||
|---|---|---|---|---|---|---|
| Live attenuated vaccine: 1-day-old (Mass) 2-, 5-, 9-, and 14-week-old (Mass + Conn) | No No | Yes Yes | Yes Yes | |||
| Inactivated IB vaccine (Mass) 14-week-old | No | Yes | No | |||
| At peak of lay | MV/MI (n = 5) | L+LK/MI (n = 8) | L+L/MI (n = 8) | MV/I (n = 9) | L+LK/I (n = 10) | L+L/I (n = 10) |
| Mass IBV challenge 26-week-old | No | No | No | Yes | Yes | Yes |
| Tissue | Microscopic Lesion |
|---|---|
| Ovary | Necrosis of ovarian covering epithelium Heterophilic and/or mononuclear cell infiltrations in the cortical stroma Congestion and edema in the cortical stroma |
| Magnum, isthmus, and uterus | Necrosis of lining epithelium and deciliation Mononuclear cell infiltrations in lamina propria Glandular dilatation Congestion and edema in the lamina propria |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, A.; Hassan, M.S.H.; Najimudeen, S.M.; Farooq, M.; Shany, S.; El-Safty, M.M.; Shalaby, A.A.; Abdul-Careem, M.F. Efficacy of Two Vaccination Strategies against Infectious Bronchitis in Laying Hens. Vaccines 2023, 11, 338. https://doi.org/10.3390/vaccines11020338
Ali A, Hassan MSH, Najimudeen SM, Farooq M, Shany S, El-Safty MM, Shalaby AA, Abdul-Careem MF. Efficacy of Two Vaccination Strategies against Infectious Bronchitis in Laying Hens. Vaccines. 2023; 11(2):338. https://doi.org/10.3390/vaccines11020338
Chicago/Turabian StyleAli, Ahmed, Mohamed S. H. Hassan, Shahnas M. Najimudeen, Muhammad Farooq, Salama Shany, Mounir Mohamed El-Safty, Adel A. Shalaby, and Mohamed Faizal Abdul-Careem. 2023. "Efficacy of Two Vaccination Strategies against Infectious Bronchitis in Laying Hens" Vaccines 11, no. 2: 338. https://doi.org/10.3390/vaccines11020338