
Identification of Closed Linear Epitopes in S1-RBD and S2-HR1/2 of SARS-CoV-2 Spike Protein Able to Induce Neutralizing Abs

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Peptide Synthesis
2.1.1. Reagents
2.1.2. Peptide Synthesis
2.2. Preparation of CPE-Conjugated Antigens
2.2.1. Conjugation of Linear Peptide on Maleimide OVA or KLH
2.2.2. SDS-PAGE Analysis of CPE-OVA
2.3. Immunization via Subcutaneous (SC) and Sublingual (SL) Routes
2.3.1. SC Immunization
2.3.2. SL Administration
2.4. ELISA for Mouse Sera, Human Sera, and Other Purified Rabbit Abs
2.4.1. Detection of IgG Abs-Specific to Individual Epitopes and Proteins
2.4.2. Epitope-Mapping of Human Sera and Purchased Rabbit Polyclonal Abs Using Individual CPEAgs
2.5. Binding Assays of RBD and ACE2
2.6. Neutralization of Pseudo-Type Lentivirus Infection
2.7. Statistical Analyses
3. Results
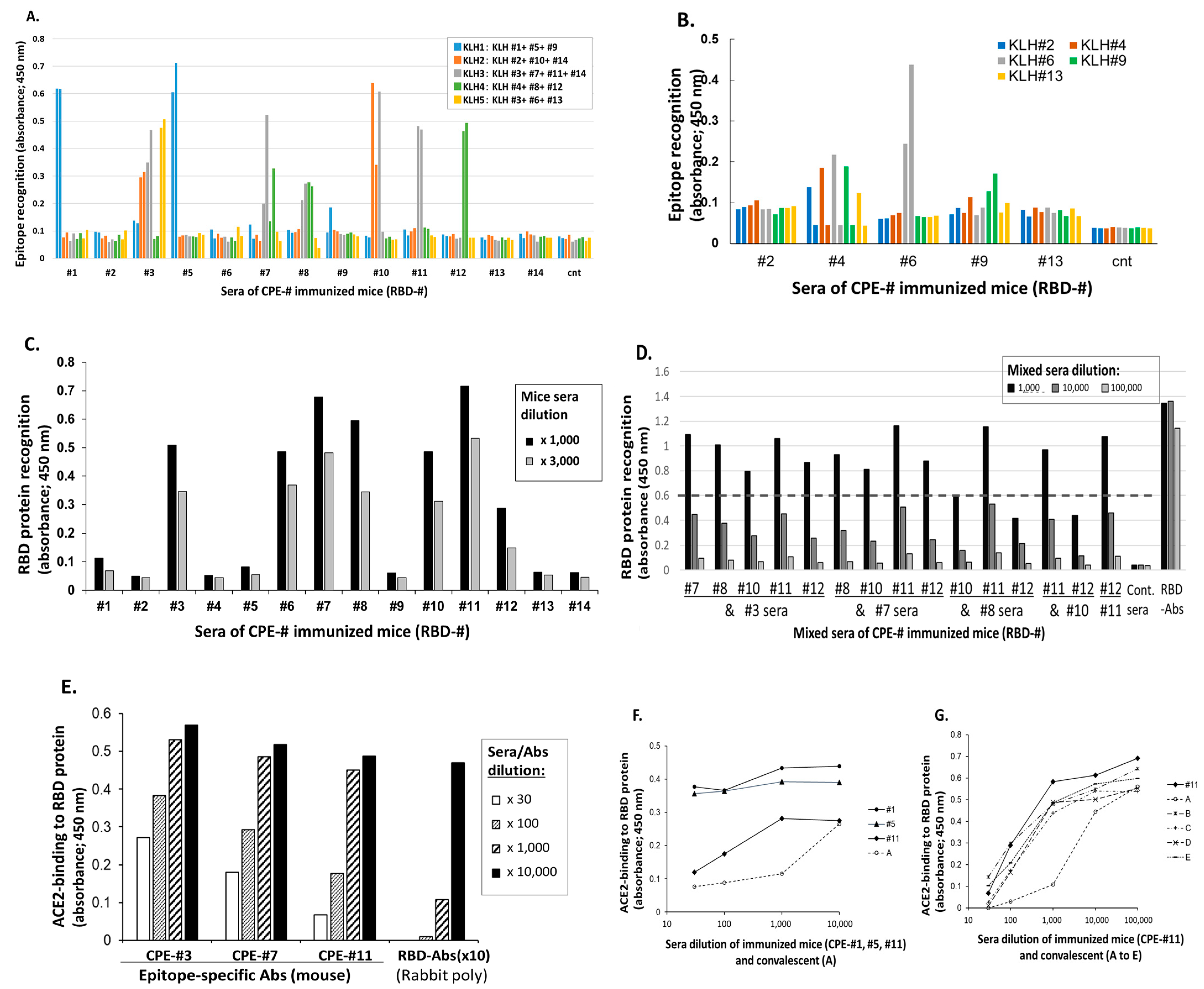
3.1. Induction of S1 RBD Epitope-Specificantisera Inhibiting the Binding of RBD to hACE2
3.1.1. Induction of S1-RBD Epitope-Specific IgG Abs in Mice
3.1.2. RBD Protein Recognition by CPE-Specific Abs
3.1.3. Ability of CPE-Specific Antisera to Inhibit Binding to hACE2
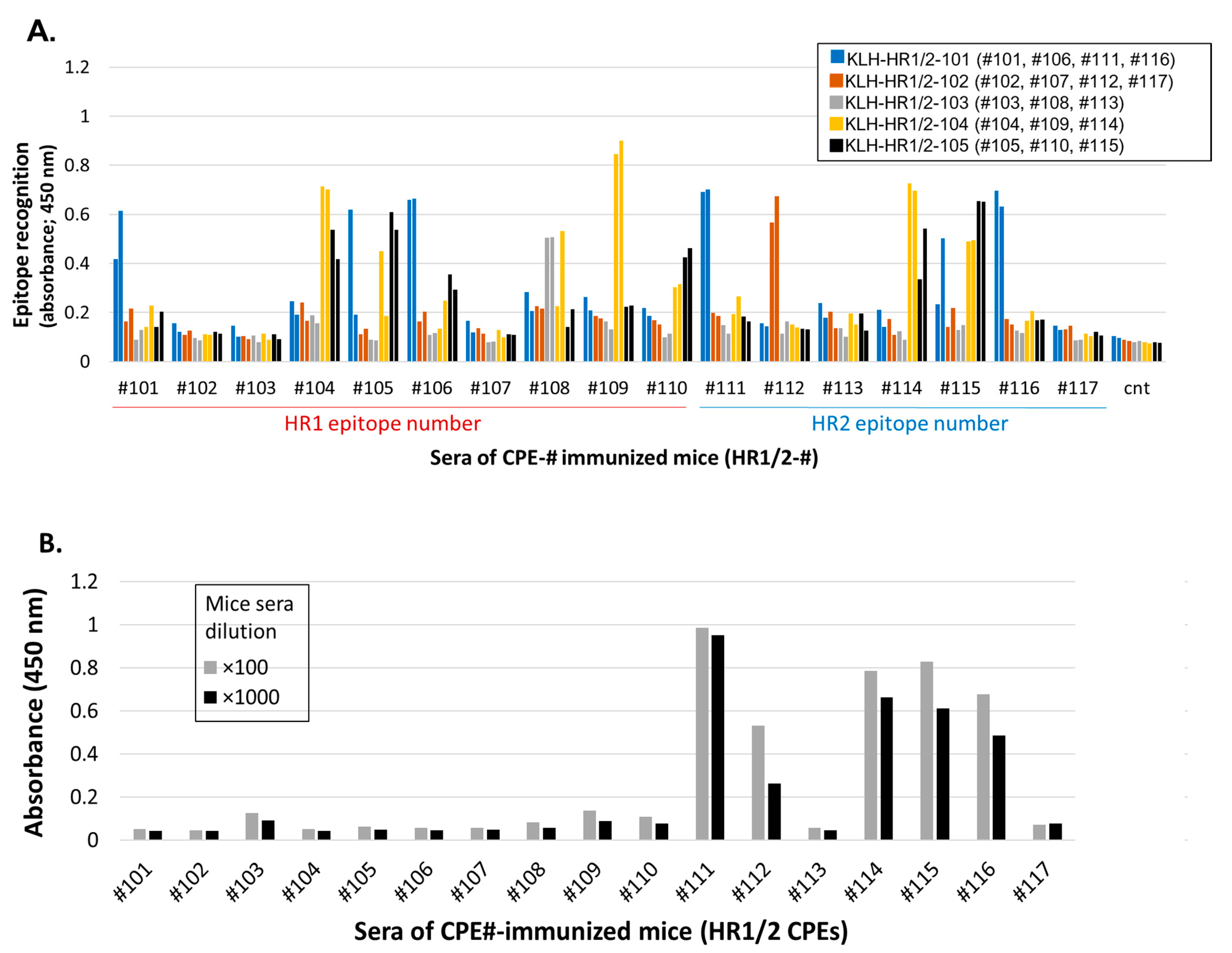
3.2. Induction of S2-HR1/2 Epitope-Specific Antisera Able to Recognize S2 Protein
3.2.1. Induction of S2-HR1/2 Epitope-Specific IgG Abs in Mice Using CPE Antigens
3.2.2. S2 Protein Recognition by CPE-Specific Antisera
3.2.3. Identification of Common Epitopes in S2-HR1/2 Using CPE-Ags
3.3. Antisera Recognizing Defined CPEs Inhibit Pseudo-Virus Infection in a Manner Mediated hACE2
3.3.1. Pseudo-Virus Infection of CRFK Cells via SARS-CoV-2 Spike Molecule
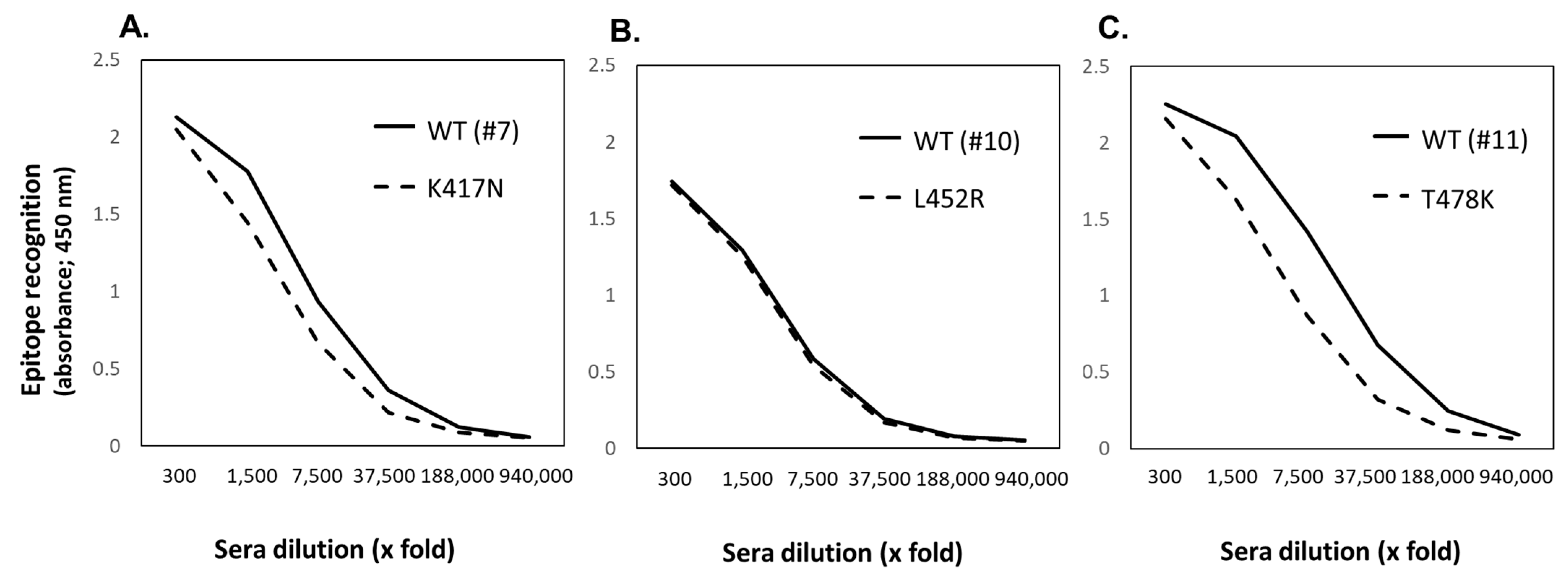
3.3.2. Selection of Ab-Epitopes Inhibiting the Process of SARS-CoV-2 Infection

{kind=link}
{kind=link}
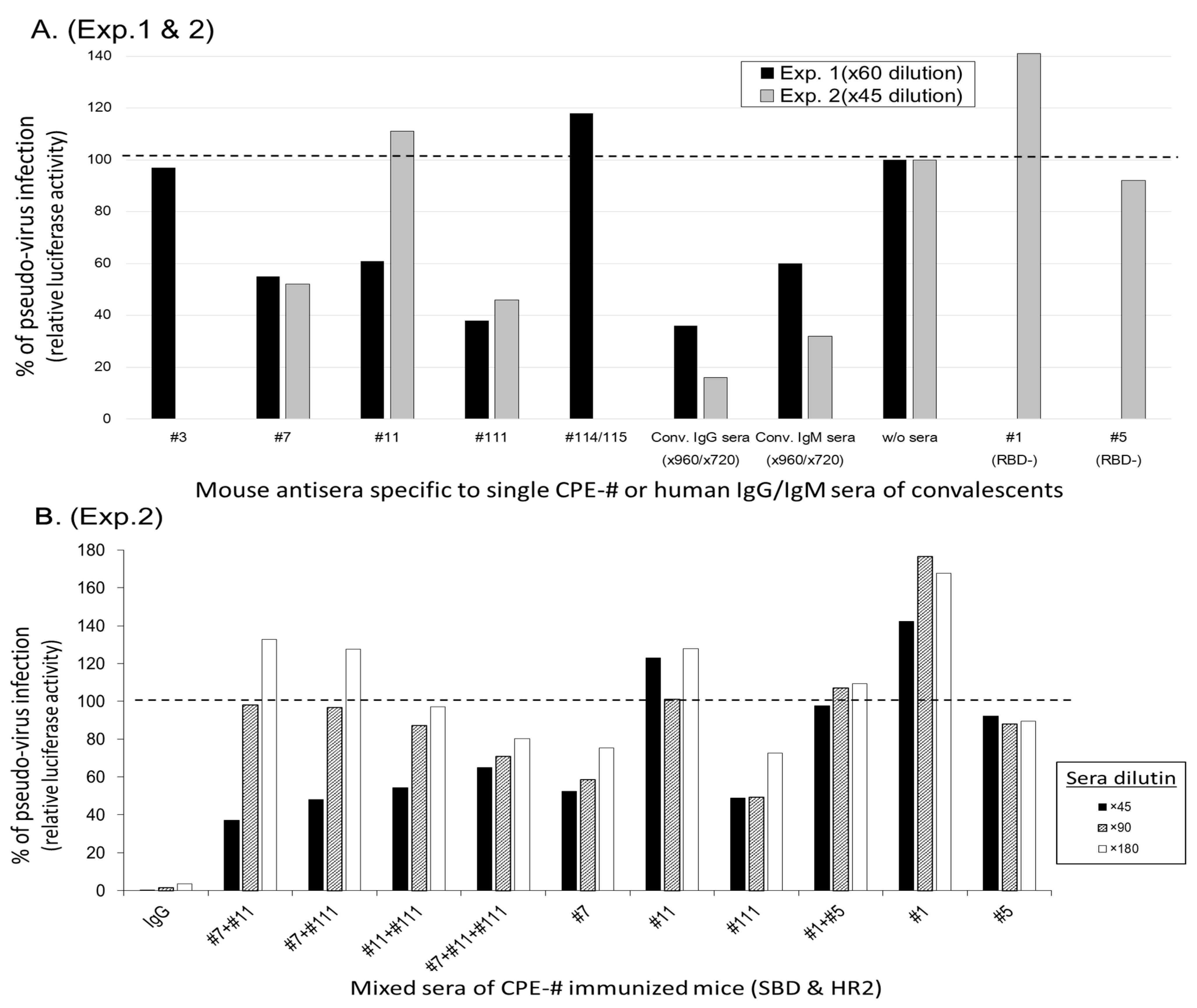{kind=link}
{kind=link}
{kind=link}
| Stepwise Selection of Infection-Responsible Epitopes of SARS-CoV-2 Spike * | ||||||
|---|---|---|---|---|---|---|
| Epitope # | Amino Acid Number | Amino Acid Sequence of SARS-CoV-2 Spike | Generation of Epitope-Abs (1) | Protein Recognition of Epitope-Abs (2) | Epitope Characterization | Pseudovirus Infection (5) |
| RBD | Inhibition of ACE2-binding by epitope-Abs (3) | |||||
| 1 | 316-336 | SNFRVQPTESIVRFPNITNLC | + | − | ||
| 2 | 336-355 | CPFGEVFNATRFASVYAWNR | − | − | ||
| 3 | 343-361 | NATRFASVYAWNRKRISNC | + | + | + | − |
| 4 | 361-379 | CVADYSVLYNSASFSTFKC | − | − | ||
| 5 | 380-392 | YGVSPTKLNDLCF | + | − | ||
| 6 | 391-414 | CFTNVYADSFVIRGDEVRQIAPGQ | + | + | − | |
| 7 | 411-432 | APGQTGKIADYNYKLPDDFTGC | + | + | + | + |
| 8 | 406-427 | EVRQIAPGQTGKIADYNYKLPD | + | + | − | |
| 9 | 431-448 | GCVIAWNSNNLDSKVGGN | − | − | ||
| 10 | 446-467 | GGNYNYLYRLFRKSNLKPFERD | + | + | − | |
| 11 | 459-480 | SNLKPFERDISTEIYQAGSTPC | + | + | + | + |
| 12 | 479-499 | PCNGVEGFNCYFPLQSYGFQP | + | + | − | |
| 13 | 500-515 | TNGVGYQPYRVVVLSF | − | − | ||
| 14 | 504-525 | GYQPYRVVVLSFELLHAPATVC | − | − | ||
| S2-HR1 | Recognition by convalescent sera (4) | |||||
| 101 | 901-920 | QMAYRFNGIGVTQNVLYENQ | + | − | − | |
| 102 | 911-929 | VTQNVLYENQKLIANQFNS | − | − | − | |
| 103 | 920-940 | QKLIANQFNSAIGKIQDSLSS | − | − | − | |
| 104 | 929-950 | SAIGKIQDSLSSTASALGKLQD | + | − | +/− | |
| 105 | 943-964 | SALGKLQDVVNQNAQALNTLVK | + | − | +/− | |
| 106 | 950-971 | DVVNQNAQALNTLVKQLSSNFG | + | − | − | |
| 107 | 963-985 | VKQLSSNFGAISSVLNDILSRLD | + | − | − | |
| 108 | 976-995 | VLNDILSRLDKVEAEVQIDR | + | − | − | |
| 109 | 988-1006 | EAEVQIDRLITGRLQSLQT | + | − | − | |
| 110 | 999-1019 | GRLQSLQTYVTQQLIRAAEIR | + | − | − | |
| S2-HR2 | ||||||
| 111 | 1144-1161 | ELDSFKEELDKYFKNHTS | + | + | + | + |
| 112 | 1154-1168 | KYFKNHTSPDVDLGD | + | + | − | |
| 113 | 1163-1181 | DVDLGDISGINASVVNIQK | − | − | − | |
| 114 | 1174-1193 | ASVVNIQKEIDRLNEVAKNL | + | + | + | − |
| 115 | 1184-1202 | DRLNEVAKNLNESLIDLQE | + | + | + | − |
| 116 | 1197-1213 | LIDLQELGKYEQYIKWP | + | + | − | |
| 117 | 1205-1223 | KYEQYIKWPWYIWLGFIAG | − | − | − | |
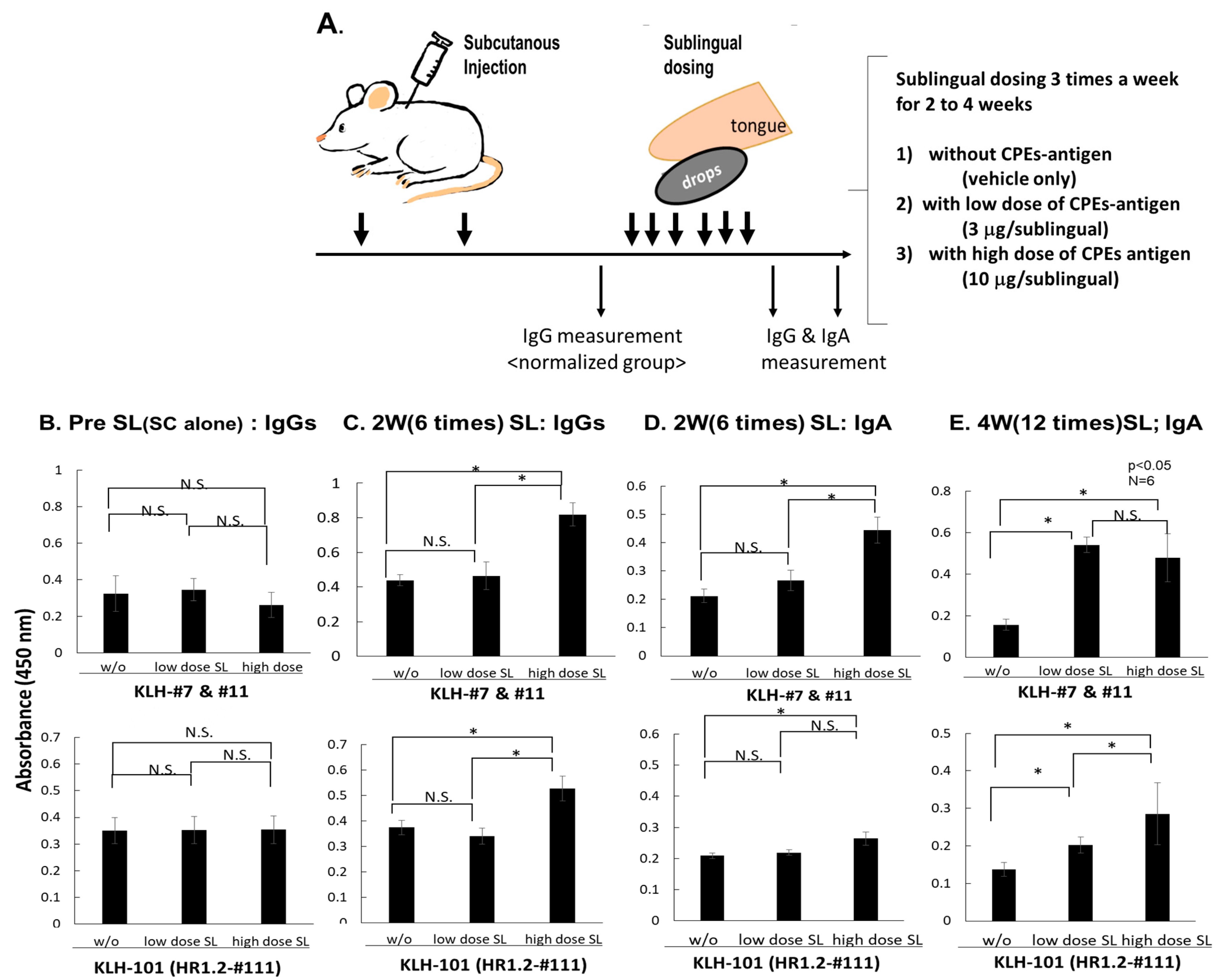
3.4. SL Administration of Defined CPEs-Ag Induces IgA Abs Specific to nAb-Epitopes
3.4.1. Experimental Setting of SL Administration to Assess Its Boosting Ability
3.4.2. Enhanced Production of IgG and IgA Specific to Neutralizing Epitopes by SL Administration
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Tracking SARS-CoV-2 Variants. Available online: www.who.int/activities/tracking-SARS-CoV-2-variants/ (accessed on 29 November 2022).
- Qu, P.; Faraone, J.; Evans, J.P.; Zou, X.; Zheng, Y.M.; Carlin, C.; Bednash, J.S.; Lozanski, G.; Mallampalli, R.K.; Saif, L.J.; et al. Neutralization of the SARS-CoV-2 Omicron BA.4/5 and BA.2.12.1 Subvariants. N. Engl. J. Med. 2022, 386, 2526–2528. [Google Scholar] [CrossRef] [PubMed]
- Anichini, G.; Terrosi, C.; Savellini, G.G.; Gandolfo, C.; Barbagli, F.; Carta, G.A.; Fabrizi, S.; Miceli, G.B.; Cusi, M.G. Antibody Response against Circulating Omicron Variants 8 Months after the Third Dose of mRNA Vaccine. Vaccines 2022, 10, 1512. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Yisimayi, A.; Jian, F.; Song, W.; Xiao, T.; Wang, L.; Du, S.; Wang, J.; Li, Q.; Chen, X. BA.2.12.1, BA.4 and BA.5 escape antibodies elicited by Omicron infection. Nature 2022, 608, 593–602. [Google Scholar] [CrossRef]
- Rössler, A.; Riepler, L.; Bante, D.; von Laer, D.; Kimpel, J. SARS-CoV-2 Omicron Variant Neutralization in Serum from Vaccinated and Convalescent Persons. N. Engl. J. Med. 2022, 386, 698–700. [Google Scholar] [CrossRef]
- Li, L.; Liao, H.; Meng, Y.; Li, Y.; Han, P.; Liu, K.; Wang, Q.; Li, D.; Zhang, Y.; Wang, L.; et al. Structural basis of human ACE2 higher binding affinity to currently circulating Omicron SARS-CoV-2 sub-variants BA.2 and BA.1.1. Cell 2022, 185, 1–9. [Google Scholar] [CrossRef]
- Xiang, F.; Wang, X.; He, X.; Peng, Z.; Yang, B.; Zhang, J.; Zhou, Q.; Ye, H.; Ma, Y.; Li, H.; et al. Antibody detection and dynamic characteristics in patients with COVID-19. Clin. Infect. Dis. 2020, 71, 1930–1934. [Google Scholar] [CrossRef]
- Okba, N.; Müller, M.A.; Li, W.; Wang, C.; Geurtsvan Kessel, C.H.; Corman, V.M.; Lamers, M.M.; Sikkema, R.S.; de Bruin, E.; Chandler, F.D.; et al. Severe Acute Respiratory Syndrome Coronavirus 2−Specific Antibody Responses in Coronavirus Disease Patients. Emerg. Infect. Dis. 2020, 26, 1478. [Google Scholar] [CrossRef]
- Kleanthous, H.; Silverman, J.M.; Makar, K.W.; Yoon, I.-K.; Jackson, N.; Vaughn, D.W. Scientific rationale for developing potent RBD-based vaccines targeting COVID-19. npj Vaccines 2021, 6, 128. [Google Scholar] [CrossRef]
- Liu, Z.; Xu, W.; Xia, S.; Gu, C.; Wang, X.; Wang, Q.; Zhou, J.; Wu, Y.; Cai, X.; Qu, D.; et al. RBD-Fc-based COVID-19 vaccine candidate induces highly potent SARS-CoV-2 neutralizing antibody response. Signal Transduct. Target. Ther. 2020, 5, 282. [Google Scholar] [CrossRef]
- Jeong, S.K.; Heo, Y.K.; Jeong, J.H.; Ham, S.J.; Yum, J.S.; Ahn, B.C.; Song, C.S.; Chun, E.Y. COVID-19 Subunit Vaccine with a Combination of TLR1/2 and TLR3 agonists Induces Robust and Protective Immunity. Vaccines 2021, 9, 957. [Google Scholar] [CrossRef] [PubMed]
- Gale, E.C.; Powell, A.E.; Roth, G.A.; Meany, E.L.; Yan, J.; Ou, B.S.; Grosskopf, A.K.; Adamska, J.; Picece, V.C.T.M.; d’Aquino, A.I.; et al. Hydrogel-Based Slow Release of a Receptor-Binding Domain Subunit Vaccine Elicits Neutralizing Antibody Responses Against SARS-CoV-2. Adv. Mater. 2021, 33, e2104362. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Bogdan, P.; Nazarian, S. An in silico deep learning approach to multi-epitope vaccine design: A SARS-CoV-2 case study. Sci. Rep. 2021, 11, 3238. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Huang, J.; Li, T.; Hu, C.; Shen, M.; Mu, S.; Luo, F.; Song, S.; Hao, Y.; Wang, W.; et al. A Highly Conserved Peptide Vaccine Candidate Activates Both Humoral and Cellular Immunity Against SARS-CoV-2 Variant Strains. Front. Immunol. 2021, 10, 1931–1946. [Google Scholar] [CrossRef] [PubMed]
- Poh, C.M.; Carissimo, G.; Wang, B.; Amrun, S.N.; Lee, C.Y.; Chee, R.S.; Fong, S.W.; Yeo, N.K.; Lee, W.H.; Torres-Ruesta, A.; et al. Two linear epitopes on the SARS-CoV-2 spike protein that elicit neutralising antibodies in COVID-19 patients. Nat. Commun. 2020, 11, 2806. [Google Scholar] [CrossRef]
- Farrera-Soler, L.; Daguer, J.; Barluenga, S.; Vadas, O.; Cohen, P.; Pagano, S.; Yerly, S.; Kaiser, L.; Vuilleumier, N.; Winssinger, N. Identification of immunodominant linear epitopes from SARS-CoV-2 patient plasma. PLoS ONE 2020, 15, e0238089. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.; Wu, L.A.; Wang, Q.; Qi, J.; FuGao, G. Cell entry by SARS-CoV-2. Trends Biochem. Sci. 2021, 4, 848–860. [Google Scholar] [CrossRef]
- Corral-Lugo, A.; Lopez-Siles, M.; Lopez, D.; MaConnell, M.J.; Martin-Galiano, A.J. Identification and Analysis of Unstructured, Linear B-Cell Epitopes in SARS-CoV-2 Virion Proteins for Vaccine Development. Vaccines 2020, 8, 397. [Google Scholar] [CrossRef]
- Dai, L.; Gao, L.; Tao, L.; Hadinegoro, S.R.; Erkin, M.; Ying, Z.; He, P.; Girsang, R.; Hlth, M.; Vergara, H.; et al. Efficacy and Safety of the RBD-Dimer-based COVID-19 Vaccine ZG2001 in Adults. N. Engl. J. Med. 2022, 386, 2097–2111. [Google Scholar] [CrossRef]
- Wong, T.Y.; Lee, K.S.; Russ, B.P.; Horspool, A.M.; Kang, J.; Winters, M.T.; Wolf, M.A.; Rader, N.A.; Miller, O.A.; Shiflett, M. Intranasal administration of BReC-CoV-2 COVID-19 vaccine protects K18-hACE2 mice against lethal SARS-CoV-2 challenge. npj Vaccines 2022, 7, 36. [Google Scholar] [CrossRef]
- Sun, W.; He, L.; Zhang, H.; Tian, Z.; Bai, Z.; Sun, L.; Yang, L.; Jia, Z.; Bi, Y.; Luo, T.; et al. The self-assembled nanoparticle-based trimeric RBD mRNA vaccine elicits robust and durable protective immunity against SARS-CoV-2 in mice. Signal Transduct. Target. Ther. 2021, 6, 340. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Hu, Y.F.; Chen, L.L.; Yau, T.; Tong, Y.G.; Hu, J.C.; Cai, J.P.; Chan, K.H.; Dou, Y.; Deng, J.; et al. Mining of epitopes on spike protein of SARS-CoV-2 from COVID-19 patients. Cell Res. 2020, 30, 702–704. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lai, D.Y.; Zhang, H.N.; Jiang, H.W.; Tian, X.L.; Ma, M.L.; Tao, S.C.; Qi, H.; Meng, Q.-F.; Guo, S.-J.; et al. Linear epitopes of SARS-CoV-2 spike protein elicit neutralizing antibodies in COVID-19 patients. Cell. Mol. Immunol. 2020, 17, 1095–1097. [Google Scholar] [CrossRef]
- Lu, S.; Xie, X.-X.; Zhao, L.; Wang, B.; Zhu, J.; Yang, T.-R.; Ji, M.; Ly, C.-P.; Xue, J.; Dai, E.-H.; et al. The immunodominant and neutralization linear epitopes for SARS-CoV-2. Cell Rep. 2021, 34, 108666. [Google Scholar] [CrossRef] [PubMed]
- Musicò, A.; Frigerio, R.; Mussida, A.; Barzon, L.; Sinigaglia, A.; Riccetti, S.; Gobbi, F.; Piubelli, C. SARS-CoV-2 Epitope Mapping on Microarrays Highlights Strong Immune-Response to N Protein Region. Vaccines 2021, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Li, J.; Huang, Z.; Li, H.; Wang, Y.; Wang, X.; Kang, S.; Huang, X.; Wu, C.; Liu, T.; et al. Structural Basis of a Human Neutralizing Antibody Specific to the SARS-CoV-2 Spike Protein Receptor-Binding Domain. Microbiol. Spectr. 2021, 9, e0135221. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, G.; Liu, H.; Ding, P.; Liu, Y.; Tian, Y.; Wang, Y.; Wang, A. Epitope Profiling Reveals the Critical Antigenic Determinants in SARS-CoV-2 RBD-Based Antigen. Front. Immunol. 2021, 12, 707977. [Google Scholar] [CrossRef]
- Shrock, E.; Fujimura, E.; Kula, T.; Timms, R.T.; Lee, I.H.; Leng, Y.; Elledge, S.J.; Robinson, M.L.; Sie, B.M.; Li, M.Z.; et al. Viral epitope profiling of COVID-19 patients reveals cross-reactivity and correlates of severity. Science 2020, 370, eabd4250. [Google Scholar] [CrossRef]
- Zhao, Q.; Gao, Y.; Xiao, M.; Huangc, X.; Wu, X. Synthesis and immunological evaluation of synthetic peptide based anti-SARS-CoV-2 vaccine candidates. Chem. Commun. 2021, 57, 1474. [Google Scholar] [CrossRef]
- Shalash, A.O.; Hussein, W.M.; Skwarczynski, M.; Toth, I. Key Considerations for the Development of Safe and Effective SARS-CoV-2 Subunit Vaccine: A Peptide-Based Vaccine Alternative. Adv. Sci. 2021, 8, 2100985. [Google Scholar] [CrossRef]
- Sakamoto, S.; Kirinashizawa, M.; Mohara, Y.; Watanabe, Y. Generation and characterization of monoclonal antibodies against mature hepcidin and its application to neutralization and quantitative alteration assay. Biosci. Biotechnol. Biochem. 2021, 85, 340–350. [Google Scholar] [CrossRef]
- Sasaki, Y.; Sano, S.; Nakahara, M.; Murata, S.; Kometani, K.; Aiba, Y.; Sakamoto, S.; Watanabe, Y.; Tanaka, K.; Kurosaki, T.; et al. Defective immune responses in mice lacking LUBAC-mediated linear ubiquitination in B cells. EMBO J. 2013, 32, 2463–2476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pısıl, Y.; Shida, H.; Miura, T. A Neutralization Assay Based on Pseudo-Typed Lentivirus with SARS CoV-2 Spike Protein in ACE2-Expressing CRFK Cells. Pathogens 2021, 10, 153. [Google Scholar] [CrossRef]
- Amrun, S.N.; Lee, C.Y.P.; Lee, B.; Fong, S.W.; Young, B.E.; Chee, R.S.L.; Ng, L.F.; Yeo, N.K.-W.; Torres-Ruesta, A.; Carissimo, G.; et al. Linear B-cell epitopes in the spike and nucleocapsid proteins as markers of SARS-CoV-2 exposure and disease severity. EBioMedicine 2020, 58, 102911. [Google Scholar] [CrossRef]
- Okuya, K.; Hattori, T.; Saito, T.; Takadate, Y.; Sasaki, M.; Furuyama, W.; Marzi, A.; Ohiro, Y.; Konno, S.; Hattori, T.; et al. Multiple Routes of Antibody-Dependent Enhancement of SARSCoV-2 Infection. Microbiol. Spectr. 2022, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wang, M.; Zhang, X.; Li, S.; Lu, Q.; Zeng, H.; Hou, H.; Li, H.; Zhang, M.; Jiang, F.; et al. Antibody-dependent cellular cytotoxicity response to SARS-CoV-2 in COVID-19 patients. Signal Transduct. Target. Ther. 2021, 6, 346. [Google Scholar] [CrossRef]
- Jiang, W.; Wang, J.; Jiao, S.; Gu, C.; Xu, W.; Chen, B.; Wang, R.; Chen, H.; Xie, Y.; Wang, A.; et al. Characterization of MW06, a human monoclonal antibody with cross-neutralization activity against both SARS-CoV-2 and SARS-CoV. MABS 2021, 13, e1953683. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Yan, R.; Liu, M.; Liu, Z.; Wang, Y.; Luan, D.; Wu, K.; Song, Z.; Sun, T.; Ma, Y.; et al. Antibody-dependent enhancement (ADE) of SARS-CoV-2 infection in recovered COVID-19 patients: Studies based on cellular and structural biology analysis. medRxiv 2020. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, Z.; Li, S.; Xu, W.; Zhang, Q.; Silva, I.T.; Li, C.; Wu, Y.; Jiang, Q.; Liu, Z.; et al. Enhancement versus neutralization by SARS-CoV-2 antibodies from a convalescent donor associates with distinct epitopes on the RBD. Cell Rep. 2021, 34, 108699. [Google Scholar] [CrossRef]
- Dacon, C.; Tucker, C.; Peng, L.; Lee, C.C.D.; Lin, T.H.; Yuan, M.; Cong, Y.; Wang, L.; Purser, L.; Williams, J.K.; et al. Broadly neutralizing antibodies target the coronavirus fusion peptide. Science 2022, 377, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, Y.; Liu, K.; Li, Y.; Lu, Q.; Wang, Q.; Zhang, Y.; Wang, L.; Liao, H.; Zheng, A.; et al. The molecular basis for SARS-CoV-2 binding to dog ACE2. Nat. Commun. 2021, 12, 4195. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Fierros, L.; García-Silva, I.; Rosales-Mendoza, S. Development of SARSCoV-2 vaccines: Should we focus on mucosal immunity? Expert Opin. Biol. Ther. 2020, 20, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Ejemel, M.; Li, Q.; Hou, S.; Schiller, Z.A.; Tree, J.A.; Wallace, A.; Amcheslavsky, A.; Yilmaz, N.K.; Buttigieg, K.R.; Elmore, M.J.; et al. A cross-reactive human IgA monoclonal antibody blocks SARS-CoV-2 spike-ACE2 interaction. Nat. Commun. 2020, 11, 4198. [Google Scholar] [CrossRef] [PubMed]
- Muecksch, F.; Wang, Z.; Cho, A.; Gaebler, C.; Tanfous, T.B.; DaSilva, J.; Bednarski, E.; Ramos, V.; Zong, S.; Johnson, B.; et al. Increased memory B cell potency and breadth after a SARS-CoV-2 mRNA boost. Nature 2022, 607, 128–134. [Google Scholar] [CrossRef] [PubMed]





| Participant\CPE Number | CPE # of Abs Positive * (Strong Positive) | ||
|---|---|---|---|
| #104/#105 | #111 | #114/#115 | |
| Convalescents recovered from moderate and severe COVID-19 (out of five convalescents) | 3/5 (0/5) | 4/5 (3/5) | 3/5 (2/5) |
| Participants vaccinated with Spike mRNA-LNPs (out of ten particiants) | 2/10 (0/10) | 8/10 (6/10) | 7/10 (4/10) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watanabe, Y.; Hosokawa, N.; Yoshida, M.; Miura, T.; Kawano, M. Identification of Closed Linear Epitopes in S1-RBD and S2-HR1/2 of SARS-CoV-2 Spike Protein Able to Induce Neutralizing Abs. Vaccines 2023, 11, 287. https://doi.org/10.3390/vaccines11020287
Watanabe Y, Hosokawa N, Yoshida M, Miura T, Kawano M. Identification of Closed Linear Epitopes in S1-RBD and S2-HR1/2 of SARS-CoV-2 Spike Protein Able to Induce Neutralizing Abs. Vaccines. 2023; 11(2):287. https://doi.org/10.3390/vaccines11020287
Chicago/Turabian StyleWatanabe, Yoshihiro, Natsuko Hosokawa, Misaki Yoshida, Tomoyuki Miura, and Mitsuhiro Kawano. 2023. "Identification of Closed Linear Epitopes in S1-RBD and S2-HR1/2 of SARS-CoV-2 Spike Protein Able to Induce Neutralizing Abs" Vaccines 11, no. 2: 287. https://doi.org/10.3390/vaccines11020287