
HBV Vaccines: Advances and Development

 ,
,
Abstract
:1. Introduction
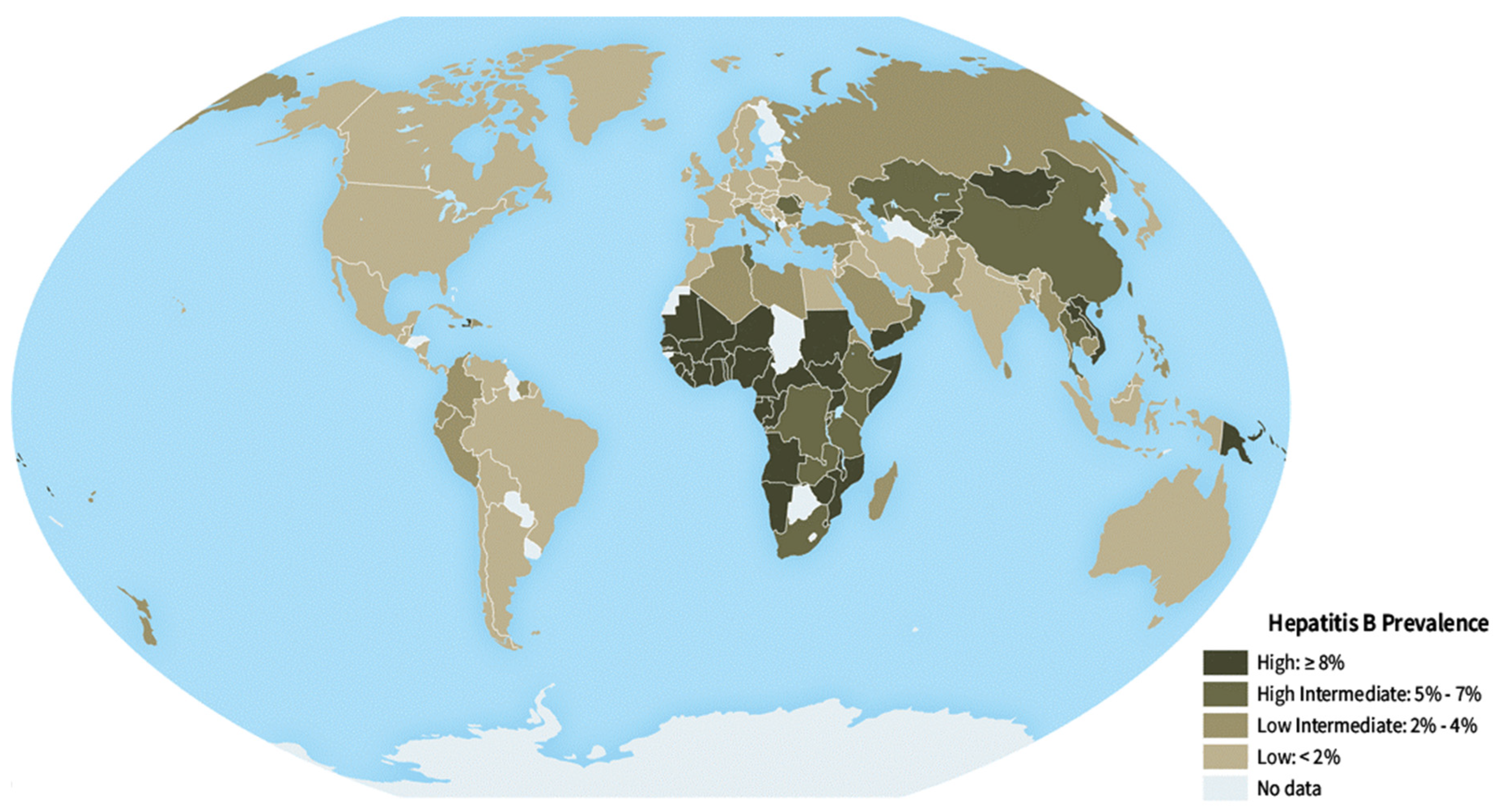
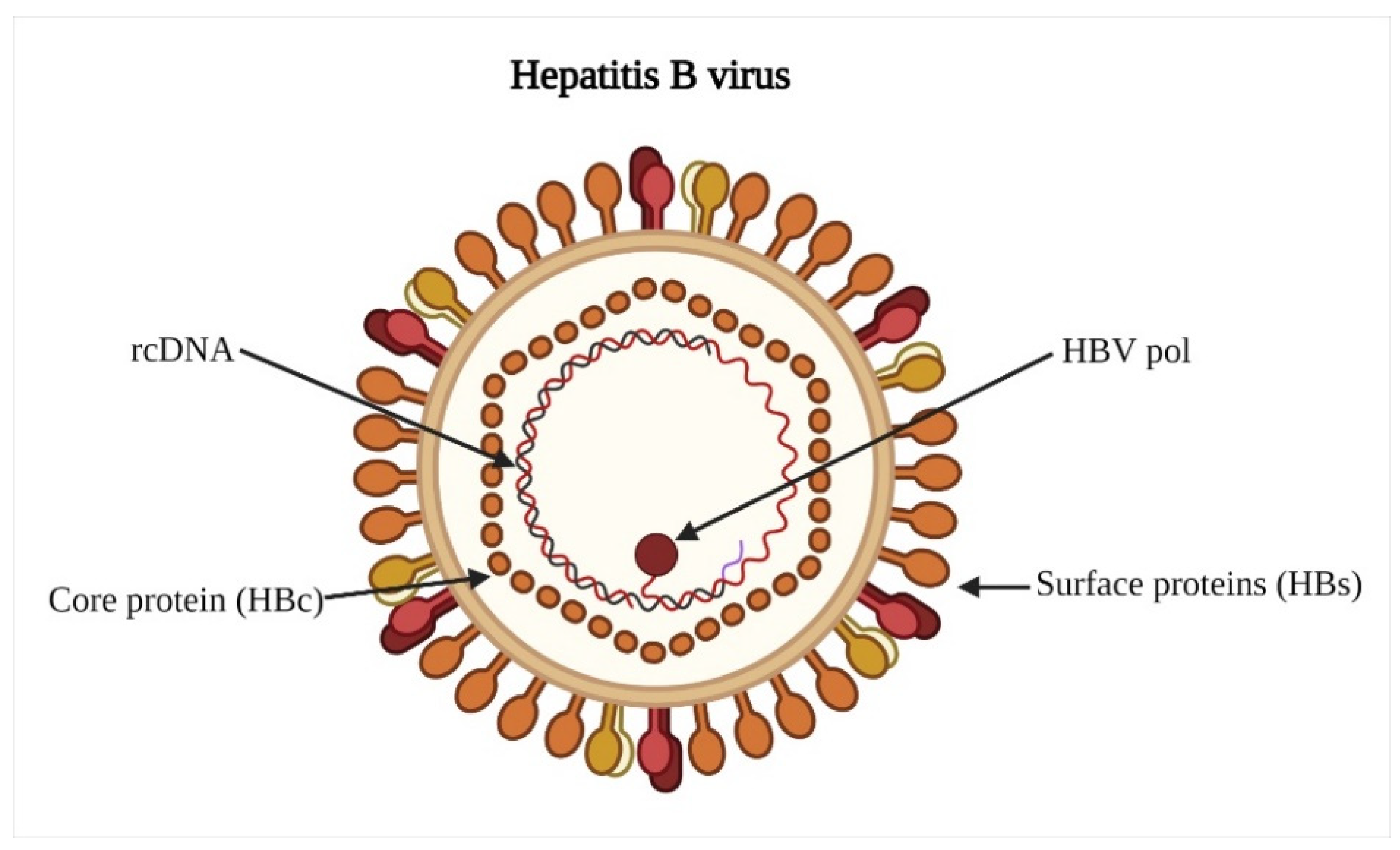
2. Hepatitis B Virus
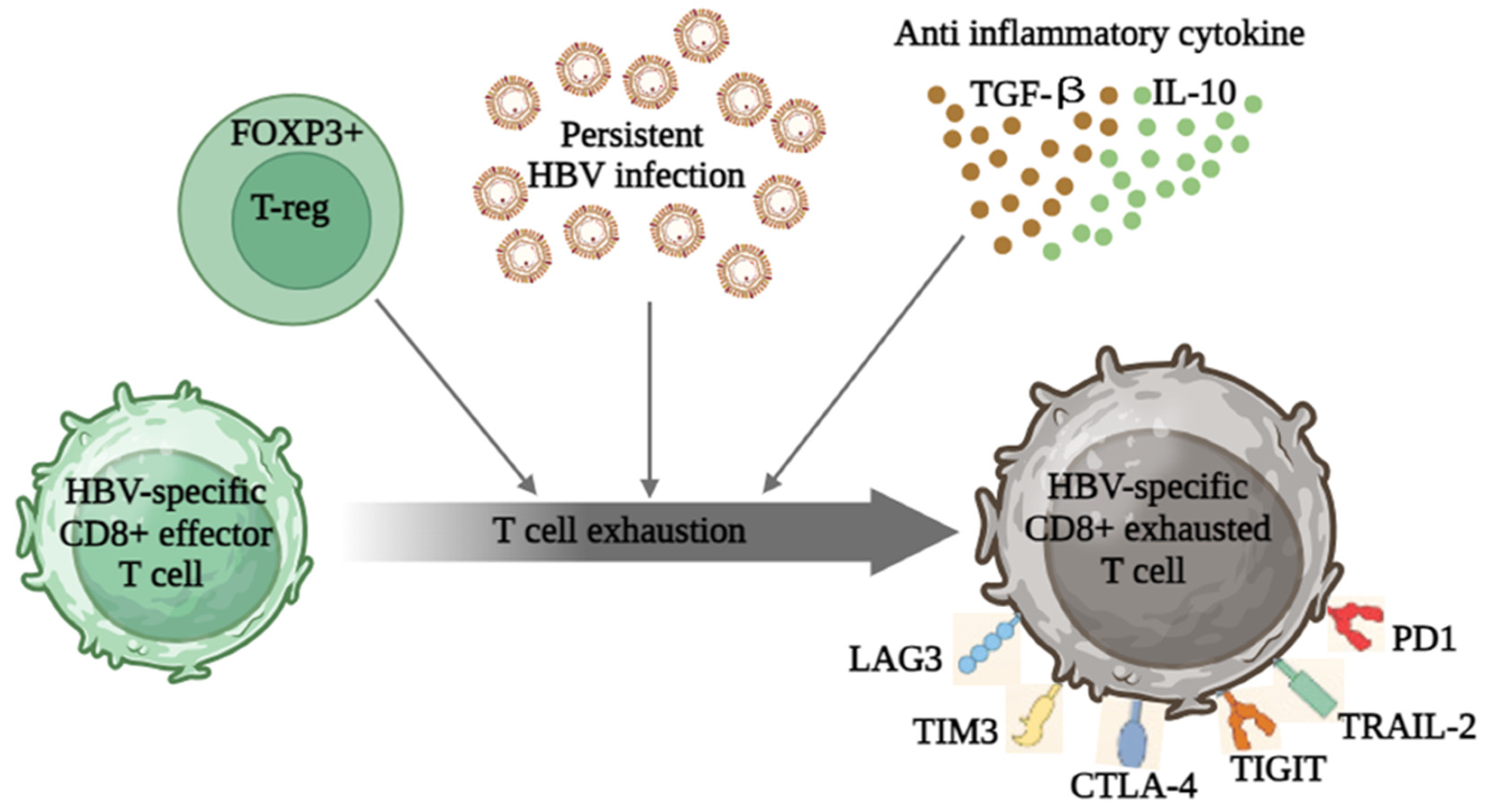
3. HBV-Induced Immune Dysregulation
4. Hepatitis B Vaccines
4.1. Needle-Free Hepatitis B Vaccine
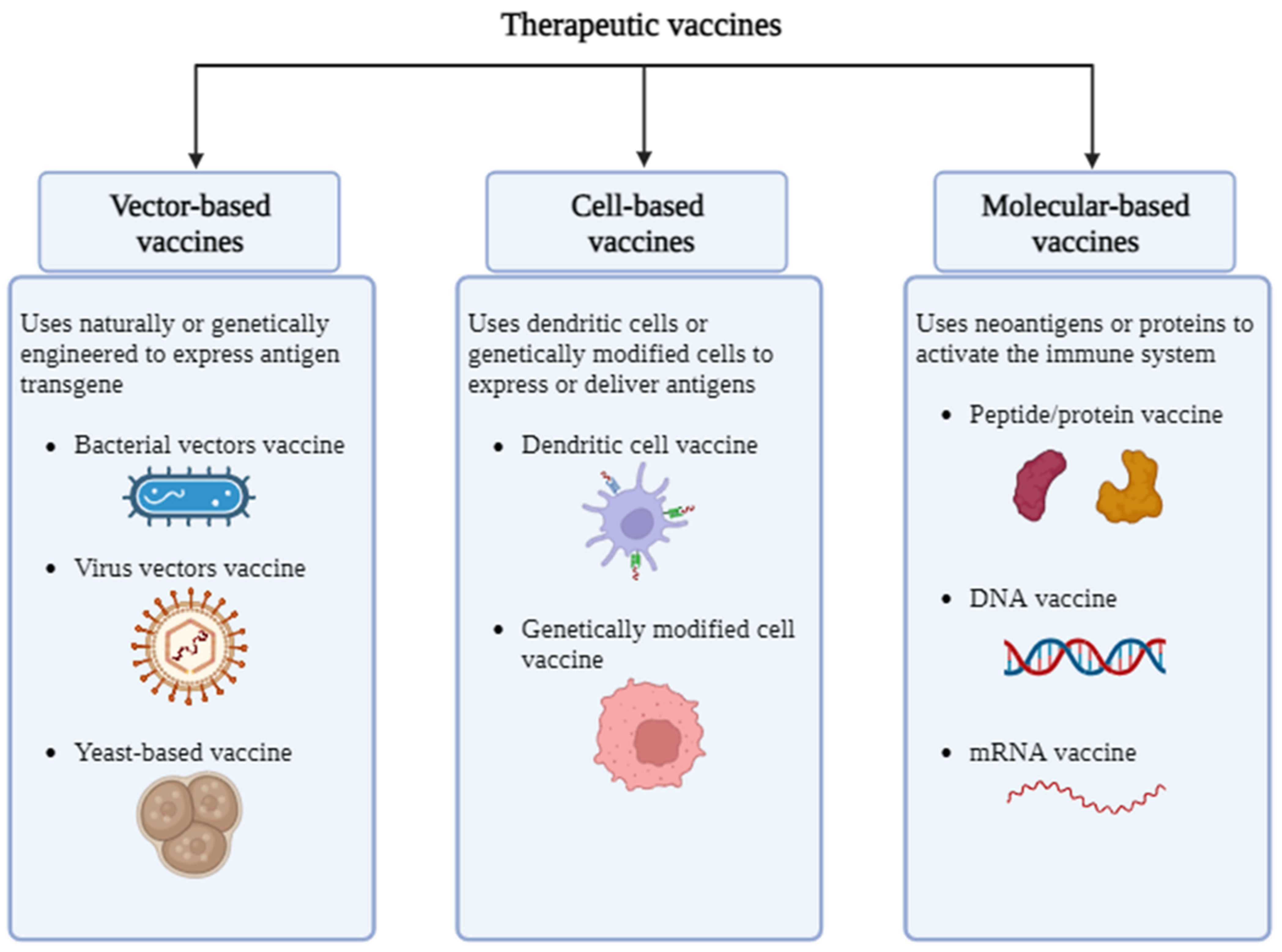
4.2. Therapeutic Vaccine
4.3. Adenovirus Vaccines
4.4. mRNA Vaccines
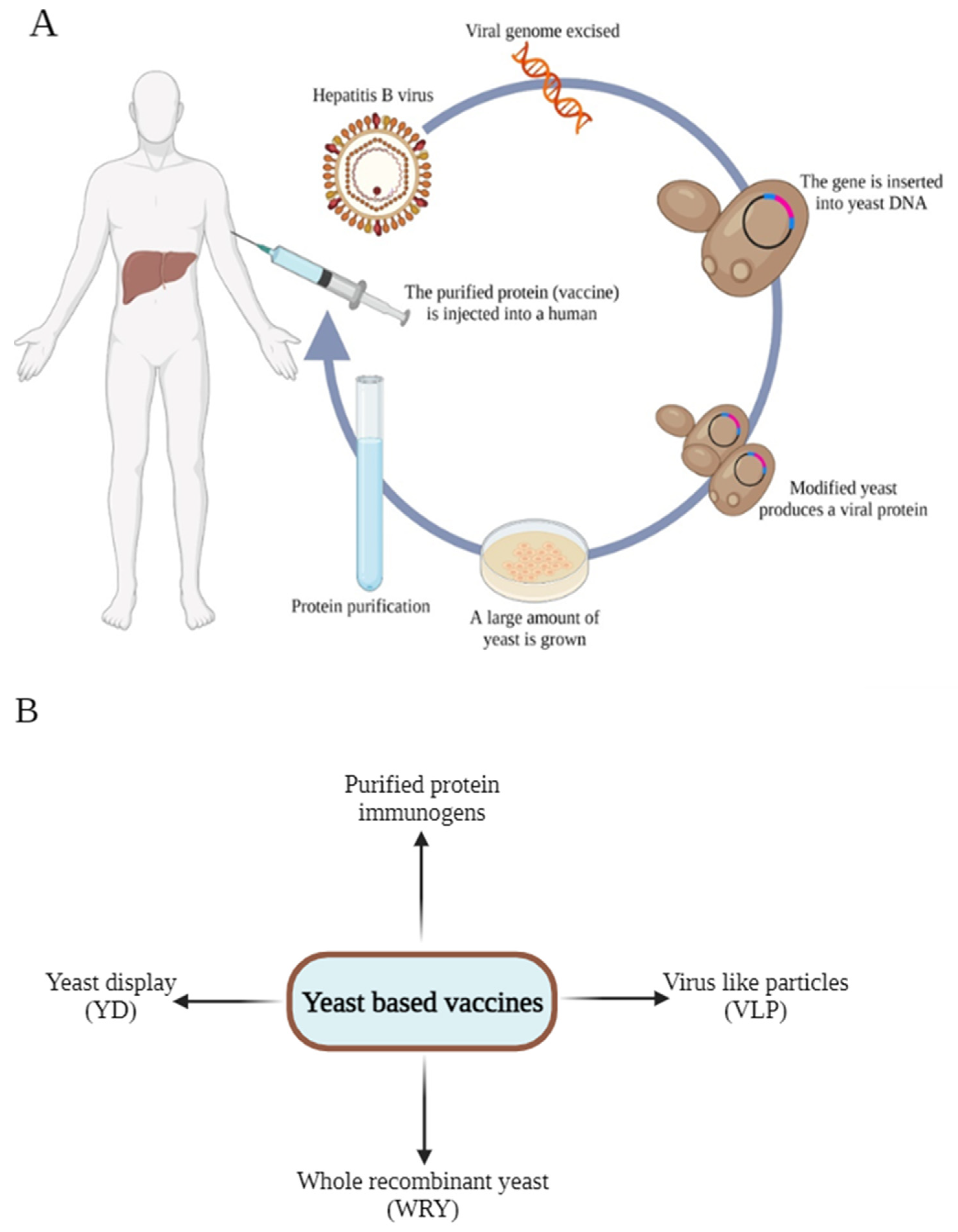
4.5. Yeast-Derived Vaccine

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast Species | Antigen/Immunogen | Expression Strategy | References |
|---|---|---|---|
| S. cerevisiae | HBsAg | Virus like particle | [93] |
| Hepatitis B surface antigen | Purified protein | [94] | |
| Hepatitis B surface antigen (HBsAg) | Virus like particle | [95] | |
| Hepatitis B virus (HBV) X, surface(S), and Core antigens (X-S-Core) | Whole recombinant yeast | [96] | |
| Surface protein GS-4774 | Whole recombinant yeast | [97] | |
| HBsAg | Virus like particle | [98] | |
| Hepatitis B core protein | Purified protein | [99] | |
| Hepatitis B surface antigen | Purified protein | [100,101] | |
| P. pastoris | HBsAg and HEnAg fusion protein | Virus like particle | [102] |
| HBsAg | Virus like particle | [103] | |
| Core protein (HBc) | Virus like particle | [104] | |
| Recombinant hepatitis B surface antigen | Purified protein | [89] | |
| HBsAg, HSP70 (1–370) | Whole recombinant yeast | [105] | |
| HBsAg | Virus like particle | [106] | |
| Hepatitis B surface antigen | Virus like particle | [107] | |
| Recombinant hepatitis B surface antigen | Virus like particle | [108] | |
| H. polymorpha | VrHB-IB | Purified protein | [109] |
4.6. DNA Vaccines
4.7. Recombinant Vaccines
4.8. Lipopeptide Epitope-Based Vaccine
4.9. Nanoparticle-Based Vaccines
4.10. HBV Vaccines in Clinical Trials
5. Concluding Remarks and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AdVs | Adenoviruses |
| APCs | Antigen Present Cells |
| cccDNA | Covalently Closed Circular DNA |
| CHO | Chinese Hamster Ovary |
| CpG | Cytosine Phosphoguanosine |
| CpG | Cytosine-Guanine Oligodeoxynucleotides |
| EV71 | Enterovirus 71 |
| FNC | Flash Nano Complexation |
| HBcAg | Hepatitis B Core Antigen |
| HBeAg | Hepatitis B E Antigen |
| HBsAg | Hepatitis B Surface Antigen |
| HBV | Hepatitis B Virus |
| HCC | Hepatocellular Carcinoma |
| HIV | Human Immunodeficiency Virus |
| LNs | Lymph Nodes |
| MAV | Mono-Antigenic Vaccine |
| MPL | Monophosphoryl Lipid |
| NE | Nanoemulsion |
| NTCP | Sodium Taurocholate Co-Transporting Polypeptide |
| ORFs | Open Reading Frames |
| rcDNA | Relaxed-Circular DNA |
| SGF | Simulated Gastric Fluid |
| SIF | Simulated Intestinal Fluid |
| VLPs | Virus-Like Particles |
References
- Hsu, Y.C.; Huang, D.Q.; Nguyen, M.H. Global Burden of Hepatitis B Virus: Current Status, Missed Opportunities and a Call for Action. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 524–537. [Google Scholar] [CrossRef]
- Rizzo, G.E.M.; Cabibbo, G.; Craxì, A. Hepatitis B Virus-Associated Hepatocellular Carcinoma. Viruses 2022, 14, 986. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, F.; Xu, R.; Awan, M.U.N.; Jia, T.; Zhang, T.; Shi, W.; Liu, M.; Han, Q.; Zhu, Q.; Zhang, Q.; et al. Transcriptomics Based Identification of S100A3 as the Key Anti-Hepatitis B Virus Factor of 16F16. Biomed. Pharmacother. 2023, 163, 114904. [Google Scholar] [CrossRef]
- Abara, W.E.; Qaseem, A.; Schillie, S.; McMahon, B.J.; Harris, A.M. Hepatitis B Vaccination, Screening, and Linkage to Care: Best Practice Advice from the American College of Physicians and the Centers for Disease Control and Prevention. Ann. Intern. Med. 2017, 167, 794–804. [Google Scholar] [CrossRef]
- Kakisaka, K.; Sakai, A.; Yoshida, Y.; Miyasaka, A.; Takahashi, F.; Sumazaki, R.; Takikawa, Y. Hepatitis B Surface Antibody Titers at One and Two Years after Hepatitis B Virus Vaccination in Healthy Young Japanese Adults. Intern. Med. 2019, 58, 2349–2355. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; Liang, T.J. Experimental Models of Hepatitis B and C—New Insights and Progress. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 362–374. [Google Scholar] [CrossRef]
- Knolle, P.A.; Huang, L.R.; Kosinska, A.; Wohlleber, D.; Protzer, U. Improving Therapeutic Vaccination against Hepatitis B-Insights from Preclinical Models of Immune Therapy against Persistent Hepatitis B Virus Infection. Vaccines 2021, 9, 1333. [Google Scholar] [CrossRef]
- CDC. CDC Yellow Book 2020: Health Information for International Travel; Brunette, G.W., Nemhauser, J.B., Eds.; Oxford University Press: Oxford, UK, 2019. [Google Scholar] [CrossRef]
- Pattyn, J.; Hendrickx, G.; Vorsters, A.; Van Damme, P. Hepatitis B Vaccines. J. Infect. Dis. 2021, 224, 343–351. [Google Scholar] [CrossRef]
- Chang, M.H.; Chen, D.S. Prevention of Hepatitis B. Cold Spring Harb. Perspect. Med. 2015, 5, a021493. [Google Scholar] [CrossRef]
- Wang, H.; Men, P.; Xiao, Y.; Gao, P.; Lv, M.; Yuan, Q.; Chen, W.; Bai, S.; Wu, J. Hepatitis B Infection in the General Population of China: A Systematic Review and Meta-Analysis. BMC Infect. Dis. 2019, 19, 811. [Google Scholar] [CrossRef]
- Hepatitis B. Available online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-b (accessed on 15 February 2023).
- Hudu, S.A.; Harmal, N.S.; Saeed, M.I.; Alshrari, A.S.; Malik, Y.A.; Niazlin, M.T.; Hassan, R.; Sekawi, Z. Naturally Occurring Hepatitis B Virus Surface Antigen Mutant Variants in Malaysian Blood Donors and Vaccinees. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1349–1359. [Google Scholar] [CrossRef]
- Tsukuda, S.; Watashi, K. Hepatitis B Virus Biology and Life Cycle. Antiviral Res. 2020, 182, 104925. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Long, Y.; Fan, X.; Liu, H.; Li, C.; Chen, L.; Wang, Z. Proteomic Analysis of Differentially Expressed Proteins in Hepatitis B Virus-Related Hepatocellular Carcinoma Tissues. J. Exp. Clin. Cancer Res. 2009, 28, 122. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Iwamoto, M.; Yun, J.H.; Uchikubo-Kamo, T.; Son, D.; Jin, Z.; Yoshida, H.; Ohki, M.; Ishimoto, N.; Mizutani, K.; et al. Structural Insights into the HBV Receptor and Bile Acid Transporter NTCP. Nature 2022, 606, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Lampertico, P.; Agarwal, K.; Berg, T.; Buti, M.; Janssen, H.L.A.; Papatheodoridis, G.; Zoulim, F.; Tacke, F. EASL 2017 Clinical Practice Guidelines on the Management of Hepatitis B Virus Infection. J. Hepatol. 2017, 67, 370–398. [Google Scholar] [CrossRef] [PubMed]
- Berg, T.; Simon, K.G.; Mauss, S.; Schott, E.; Heyne, R.; Klass, D.M.; Eisenbach, C.; Welzel, T.M.; Zachoval, R.; Felten, G.; et al. Long-Term Response after Stopping Tenofovir Disoproxil Fumarate in Non-Cirrhotic HBeAg-Negative Patients—FINITE Study. J. Hepatol. 2017, 67, 918–924. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Ploss, A. Mechanism of Hepatitis B Virus CccDNA Formation. Viruses 2021, 13, 1463. [Google Scholar] [CrossRef]
- Khanam, A.; Chua, J.V.; Kottilil, S. Immunopathology of Chronic Hepatitis B Infection: Role of Innate and Adaptive Immune Response in Disease Progression. Int. J. Mol. Sci. 2021, 22, 5497. [Google Scholar] [CrossRef]
- Dembek, C.; Protzer, U.; Roggendorf, M. Overcoming Immune Tolerance in Chronic Hepatitis B by Therapeutic Vaccination. Curr. Opin. Virol. 2018, 30, 58–67. [Google Scholar] [CrossRef]
- Huang, H.; Rückborn, M.; Le-Trilling, V.T.K.; Zhu, D.; Yang, S.; Zhou, W.; Yang, X.; Feng, X.; Lu, Y.; Lu, M.; et al. Prophylactic and Therapeutic HBV Vaccination by an HBs-Expressing Cytomegalovirus Vector Lacking an Interferon Antagonist in Mice. Eur. J. Immunol. 2021, 51, 393–407. [Google Scholar] [CrossRef]
- Ma, Z.; Cao, Q.; Xiong, Y.; Zhang, E.; Lu, M. Interaction between Hepatitis B Virus and Toll-Like Receptors: Current Status and Potential Therapeutic Use for Chronic Hepatitis B. Vaccines 2018, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Pallett, L.J.; Gill, U.S.; Quaglia, A.; Sinclair, L.V.; Jover-Cobos, M.; Schurich, A.; Singh, K.P.; Thomas, N.; Das, A.; Chen, A.; et al. Metabolic Regulation of Hepatitis B Immunopathology by Myeloid-Derived Suppressor Cells. Nat. Med. 2015, 21, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Li, T.Y.; Yang, Y.; Zhou, G.; Tu, Z.K. Immune Suppression in Chronic Hepatitis B Infection Associated Liver Disease: A Review. World J. Gastroenterol. 2019, 25, 3527–3537. [Google Scholar] [CrossRef] [PubMed]
- Hoogeveen, R.C.; Boonstra, A. Checkpoint Inhibitors and Therapeutic Vaccines for the Treatment of Chronic HBV Infection. Front. Immunol. 2020, 11, 401. [Google Scholar] [CrossRef] [PubMed]
- McAleer, W.J.; Buynak, E.B.; Maigetter, R.Z.; Wampler, D.E.; Miller, W.J.; Hilleman, M.R. Human Hepatitis B Vaccine from Recombinant Yeast. Nature 1984, 307, 178–180. [Google Scholar] [CrossRef] [PubMed]
- Emini, E.A.; Ellis, R.W.; Miller, W.J.; McAleer, W.J.; Scolnick, E.M.; Gerety, R.J. Production and Immunological Analysis of Recombinant Hepatitis B Vaccine. J. Infect. 1986, 13 (Suppl. A), 3–9. [Google Scholar] [CrossRef]
- Bucci, M. First Recombinant DNA Vaccine for HBV. Nat. Res. 2020, 13, 3–9. [Google Scholar]
- Hepatitis B Foundation: History of Hepatitis B Vaccine. Available online: https://www.hepb.org/prevention-and-diagnosis/vaccination/history-of-hepatitis-b-vaccine/ (accessed on 15 February 2023).
- Edey, M.; Barraclough, K.; Johnson, D.W. Review Article: Hepatitis B and Dialysis. Nephrology 2010, 15, 137–145. [Google Scholar] [CrossRef]
- Purcell, R.H.; Gerin, J.L. Prospects for Second and Third Generation Hepatitis B Vaccines. Hepatology 1985, 5, 159–163. [Google Scholar] [CrossRef]
- Tong, N.K.C.; Beran, J.; Kee, S.A.; Miguel, J.L.; Sánchez, C.; Bayas, J.M.; Vilella, A.; De Juanes, J.R.; Arrazola, P.; Calbo-Torrecillas, F.; et al. Immunogenicity and Safety of an Adjuvanted Hepatitis B Vaccine in Pre-Hemodialysis and Hemodialysis Patients. Kidney Int. 2005, 68, 2298–2303. [Google Scholar] [CrossRef]
- Rendi-Wagner, P.; Shouval, D.; Genton, B.; Lurie, Y.; Rümke, H.; Boland, G.; Cerny, A.; Heim, M.; Bach, D.; Schroeder, M.; et al. Comparative Immunogenicity of a PreS/S Hepatitis B Vaccine in Non- and Low Responders to Conventional Vaccine. Vaccine 2006, 24, 2781–2789. [Google Scholar] [CrossRef] [PubMed]
- Vesikari, T.; Langley, J.M.; Segall, N.; Ward, B.J.; Cooper, C.; Poliquin, G.; Smith, B.; Gantt, S.; McElhaney, J.E.; Dionne, M.; et al. Immunogenicity and Safety of a Tri-Antigenic versus a Mono-Antigenic Hepatitis B Vaccine in Adults (PROTECT): A Randomised, Double-Blind, Phase 3 Trial. Lancet. Infect. Dis. 2021, 21, 1271–1281. [Google Scholar] [CrossRef]
- Romano’, L.; Zanetti, A.R. Hepatitis B Vaccination: A Historical Overview with a Focus on the Italian Achievements. Viruses 2022, 14, 1515. [Google Scholar] [CrossRef] [PubMed]
- Pollard, A.J.; Bijker, E.M. A Guide to Vaccinology: From Basic Principles to New Developments. Nat. Rev. Immunol. 2020, 21, 83–100. [Google Scholar] [CrossRef]
- Skibinski, D.A.G.; Baudner, B.C.; Singh, M.; O’hagan, D.T. Combination Vaccines. J. Glob. Infect. Dis. 2011, 3, 63. [Google Scholar] [CrossRef] [PubMed]
- Kramvis, A.; Mammas, I.N.; Spandidos, D.A. Exploring the Optimal Vaccination Strategy against Hepatitis B Virus in Childhood (Review). Biomed. Rep. 2023, 19, 48. [Google Scholar] [CrossRef]
- Mohanty, P.; Jena, P.; Patnaik, L. Vaccination against Hepatitis B: A Scoping Review. Asian Pac. J. Cancer Prev. 2020, 21, 3453–3459. [Google Scholar] [CrossRef]
- Tan, A.; Koh, S.; Bertoletti, A. Immune Response in Hepatitis B Virus Infection. Cold Spring Harb. Perspect. Med. 2015, 5, a021428. [Google Scholar] [CrossRef]
- Zhang, X.; Bai, L.; Zhang, S.; Zhou, X.; Li, Y.; Bai, J. Trx-1 Ameliorates Learning and Memory Deficits in MPTP-Induced Parkinson’s Disease Model in Mice. Free Radic. Biol. Med. 2018, 124, 380–387. [Google Scholar] [CrossRef]
- Lee, G.H.; Lim, S.G. CpG-Adjuvanted Hepatitis B Vaccine (HEPLISAV-B®) Update. Expert Rev. Vaccines 2021, 20, 487–495. [Google Scholar] [CrossRef]
- Horta, D.; Forné, M.; Agustí, A.; Raga, A.; Martín-Cardona, A.; Hernández-Soto, J.M.; Ruiz-Ramírez, P.; Esteve-Comas, M. Efficacy of Hepatitis B Virus Vaccines HBVaxpro40© and Fendrix© in Patients with Chronic Liver Disease in Clinical Practice. Vaccines 2022, 10, 1323. [Google Scholar] [CrossRef]
- Nevens, F.; Zuckerman, J.N.; Burroughs, A.K.; Jung, M.C.; Bayas, J.M.; Kallinowski, B.; Rivas, E.F.; Duvoux, C.; Neuhaus, P.; Saliba, F.; et al. Immunogenicity and Safety of an Experimental Adjuvanted Hepatitis B Candidate Vaccine in Liver Transplant Patients. Liver Transplant. 2006, 12, 1489–1495. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.; MacKie, D. Hepatitis B Surface Antigen-1018 ISS Adjuvant-Containing Vaccine: A Review of HEPLISAVTM Safety and Efficacy. Expert Rev. Vaccines 2011, 10, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, Y.; Sun, Y.; Cui, H.; Zhu, S.J.; Qiu, H.J. Mucosal Vaccines: Strategies and Challenges. Immunol. Lett. 2020, 217, 116–125. [Google Scholar] [CrossRef]
- El-Sayed, M.H.; Feld, J.J. Vaccination at the Forefront of the Fight against Hepatitis B and C. Nat. Rev. Gastroenterol. Hepatol. 2021, 19, 87–88. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.W.; Mestecky, J. Mucosal Vaccines: An Overview. Mucosal Immunol. Fourth Ed. 2015, 1–2, 1039–1046. [Google Scholar] [CrossRef]
- Vajdy, M.; O’Hagan, D.T. Microparticles for Intranasal Immunization. Adv. Drug Deliv. Rev. 2001, 51, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Saraf, S.; Mishra, D.; Asthana, A.; Jain, R.; Singh, S.; Jain, N.K. Lipid Microparticles for Mucosal Immunization against Hepatitis B. Vaccine 2006, 24, 45–56. [Google Scholar] [CrossRef]
- Khatri, K.; Goyal, A.K.; Gupta, P.N.; Mishra, N.; Mehta, A.; Vyas, S.P. Surface Modified Liposomes for Nasal Delivery of DNA Vaccine. Vaccine 2008, 26, 2225–2233. [Google Scholar] [CrossRef]
- Deacon, M.P.; McGurk, S.; Roberts, C.J.; Williams, P.M.; Tendler, S.J.B.; Davies, M.C.; Davis, S.S.; Harding, S.E. Atomic Force Microscopy of Gastric Mucin and Chitosan Mucoadhesive Systems. Biochem. J. 2000, 348, 557. [Google Scholar] [CrossRef]
- Singh, P.; Prabakaran, D.; Jain, S.; Mishra, V.; Jaganathan, K.S.; Vyas, S.P. Cholera Toxin B Subunit Conjugated Bile Salt Stabilized Vesicles (Bilosomes) for Oral Immunization. Int. J. Pharm. 2004, 278, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Mann, J.F.S.; Scales, H.E.; Shakir, E.; Alexander, J.; Carter, K.C.; Mullen, A.B.; Ferro, V.A. Oral Delivery of Tetanus Toxoid Using Vesicles Containing Bile Salts (Bilosomes) Induces Significant Systemic and Mucosal Immunity. Methods 2006, 38, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.; Khatri, K.; Gupta, P.N.; Goyal, A.K.; Mehta, A.; Vyas, S.P. Oral Immunization against Hepatitis B Using Bile Salt Stabilized Vesicles (Bilosomes). J. Pharm. Pharm. Sci. 2008, 11, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Hanning, S.; Falconer, J.; Locke, M.; Wen, J. Recent Advances in Non-Ionic Surfactant Vesicles (Niosomes): Fabrication, Characterization, Pharmaceutical and Cosmetic Applications. Eur. J. Pharm. Biopharm. 2019, 144, 18–39. [Google Scholar] [CrossRef]
- Makidon, P.E.; Bielinska, A.U.; Nigavekar, S.S.; Janczak, K.W.; Knowlton, J.; Scott, A.J.; Mank, N.; Cao, Z.; Rathinavelu, S.; Beer, M.R.; et al. Pre-Clinical Evaluation of a Novel Nanoemulsion-Based Hepatitis B Mucosal Vaccine. PLoS ONE 2008, 3, e2954. [Google Scholar] [CrossRef]
- Shan, P.; Wang, Z.; Li, J.; Wei, D.; Zhang, Z.; Hao, S.; Hou, Y.; Wang, Y.; Li, S.; Wang, X.; et al. A New Nano Adjuvant of PF3 Used for an Enhanced Hepatitis B Vaccine. Front. Bioeng. Biotechnol. 2022, 10, 903424. [Google Scholar] [CrossRef]
- Hudu, S.A.; Jimoh, A.O.; Ibrahim, K.G.; Alshrari, A.S. Hepatitis B Therapeutic Vaccine: A Patent Review. Pharmaceuticals 2022, 15, 1542. [Google Scholar] [CrossRef]
- Lobaina, Y.; Michel, M.L. Chronic Hepatitis B: Immunological Profile and Current Therapeutic Vaccines in Clinical Trials. Vaccine 2017, 35, 2308–2314. [Google Scholar] [CrossRef]
- Cargill, T.; Barnes, E. Therapeutic Vaccination for Treatment of Chronic Hepatitis B. Clin. Exp. Immunol. 2021, 205, 106. [Google Scholar] [CrossRef]
- Jansen, D.T.S.L.; Dou, Y.; de Wilde, J.W.; Woltman, A.M.; Buschow, S.I. Designing the Next-generation Therapeutic Vaccines to Cure Chronic Hepatitis B: Focus on Antigen Presentation, Vaccine Properties and Effect Measures. Clin. Transl. Immunol. 2021, 10, e1232. [Google Scholar] [CrossRef]
- Kosinska, A.D.; Bauer, T.; Protzer, U. Therapeutic Vaccination for Chronic Hepatitis B. Curr. Opin. Virol. 2017, 23, 75–81. [Google Scholar] [CrossRef]
- Buschow, S.I.; Jansen, D.T.S.L. CD4+ T Cells in Chronic Hepatitis B and T Cell-Directed Immunotherapy. Cells 2021, 10, 1114. [Google Scholar] [CrossRef] [PubMed]
- Kutscher, S.; Bauer, T.; Dembek, C.; Sprinzl, M.; Protzer, U. Design of Therapeutic Vaccines: Hepatitis B as an Example. Microb. Biotechnol. 2012, 5, 270. [Google Scholar] [CrossRef] [PubMed]
- Study Details | A Heterologous Protein Prime/MVA Boost Therapeutic Hepatitis B Vaccine Candidate | ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/study/NCT05727267?cond=HepatitisB&term=TherapeuticVaccine&page=4&rank=35 (accessed on 6 November 2023).
- Hcj, E. VRON-0200, a Therapeutic HBV Vaccine with an Intrinsic Checkpoint Inhibitor, Elicits Broad CD8 + T Cell Responses and Sustained Antiviral Declines in Preclinical Studies. J. Hepatol. 2021, 75, S290–S291. [Google Scholar] [CrossRef]
- Maini, M.K.; Burton, A.R. Restoring, Releasing or Replacing Adaptive Immunity in Chronic Hepatitis B. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 662–675. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.chictr.org.cn/showproj.html?proj=198415 (accessed on 6 November 2023).
- Available online: https://www.chictr.org.cn/showproj.html?proj=183135 (accessed on 6 November 2023).
- China Clinical Trial Registry—World Health Organization International Clinical Trial Registry Platform Level 1 Registry. Available online: https://www.chictr.org.cn/showproj.html?proj=183135&u_atoken=7477eb86-915c-4bd8-85c4-29b1ca1e572c&u_asession=01ZfhbNQOfOJ_ecqQ0_QOWH4A8b2zUWiLpLD8kSK6Et8mg7uqnrPUmSDJLzRtMgc04ujM1u0oUGCbLm6v005ihKdsq8AL43dpOnCClYrgFm6o&u_asig=05erbUuFtwb2BbVr7aucPIlu4pKvNDk0O8zEXddpHvLdejnIZEc7kqGucWIxOx-Utgz0-mx52kpJX59Yhzj_HJ_BLV7j1i9wnuMuB7o4Fv63WDGoVL0Oo44bxgn5Dfu0_6v8U-8xQzApxtWeE0CpNVBLUEjSKMPVK9VeFo-qJ7-5ejV6XNVlaYeeHXjtIGZAUnksmHjM0JOodanL5-M1Qs1asNyZAtJD9jrxEnch2Yt2m4oyWgyTPs5zdCpIl4hjYgcP4J8Me-anrE00YmXHZl8uf4U8xf7t-0n_0Y_GFhTHXY94r_LXIIil3Y3aVPRGAe&u_aref=edjYh6p50KrglwfDmAhHhc1HlLA%3D (accessed on 6 November 2023).
- ISA104 (HBV)—ISA Therapeutics BV. Available online: https://www.isa-pharma.com/isa104-hbv/ (accessed on 6 November 2023).
- Draper, S.J.; Heeney, J.L. Viruses as Vaccine Vectors for Infectious Diseases and Cancer. Nat. Rev. Microbiol. 2010, 8, 62–73. [Google Scholar] [CrossRef]
- Mahmood, F.; Xu, R.; Awan, M.U.N.; Song, Y.; Han, Q.; Xia, X.; Zhang, J. PDIA3: Structure, Functions and Its Potential Role in Viral Infections. Biomed. Pharmacother. 2021, 143, 112110. [Google Scholar] [CrossRef]
- Henao-Restrepo, A.M.; Camacho, A.; Longini, I.M.; Watson, C.H.; Edmunds, W.J.; Egger, M.; Carroll, M.W.; Dean, N.E.; Diatta, I.; Doumbia, M.; et al. Efficacy and Effectiveness of an RVSV-Vectored Vaccine in Preventing Ebola Virus Disease: Final Results from the Guinea Ring Vaccination, Open-Label, Cluster-Randomised Trial (Ebola Ça Suffit!). Lancet 2017, 389, 505–518. [Google Scholar] [CrossRef]
- Farhad, T.; Neves, K.; Arbuthnot, P.; Maepa, M.B. Adenoviral Vectors: Potential as Anti-HBV Vaccines and Therapeutics. Genes 2022, 13, 1941. [Google Scholar] [CrossRef]
- Tatsis, N.; Ertl, H.C.J. Adenoviruses as Vaccine Vectors. Mol. Ther. 2004, 10, 616–629. [Google Scholar] [CrossRef]
- Coughlan, L.; Kremer, E.J.; Shayakhmetov, D.M. Adenovirus-Based Vaccines-a Platform for Pandemic Preparedness against Emerging Viral Pathogens. Mol. Ther. 2022, 30, 1822–1849. [Google Scholar] [CrossRef]
- Martin, P.; Dubois, C.; Jacquier, E.; Dion, S.; Mancini-Bourgine, M.; Godon, O.; Kratzer, R.; Lelu-Santolaria, K.; Evlachev, A.; Meritet, J.F.; et al. TG1050, an Immunotherapeutic to Treat Chronic Hepatitis B, Induces Robust T Cells and Exerts an Antiviral Effect in HBV-Persistent Mice. Gut 2015, 64, 1961–1971. [Google Scholar] [CrossRef] [PubMed]
- Chinnakannan, S.K.; Cargill, T.N.; Donnison, T.A.; Ansari, M.A.; Sebastian, S.; Lee, L.N.; Hutchings, C.; Klenerman, P.; Maini, M.K.; Evans, T.; et al. The Design and Development of a Multi-HBV Antigen Encoded in Chimpanzee Adenoviral and Modified Vaccinia Ankara Viral Vectors: A Novel Therapeutic Vaccine Strategy against HBV. Vaccines 2020, 8, 184. [Google Scholar] [CrossRef]
- Bloom, K.; Ely, A.; Mussolino, C.; Cathomen, T.; Arbuthnot, P. Inactivation of Hepatitis B Virus Replication in Cultured Cells and in Vivo with Engineered Transcription Activator-like Effector Nucleases. Mol. Ther. 2013, 21, 1889–1897. [Google Scholar] [CrossRef] [PubMed]
- Stone, D.; Long, K.R.; Loprieno, M.A.; De Silva Feelixge, H.S.; Kenkel, E.J.; Liley, R.M.; Rapp, S.; Roychoudhury, P.; Nguyen, T.; Stensland, L.; et al. CRISPR-Cas9 Gene Editing of Hepatitis B Virus in Chronically Infected Humanized Mice. Mol. Ther. Methods Clin. Dev. 2020, 20, 258–275. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.A. A Comparison of Plasmid DNA and MRNA as Vaccine Technologies. Vaccines 2019, 7, 37. [Google Scholar] [CrossRef]
- Lamb, C.; Arbuthnot, P. Development of an MRNA Vaccination Strategy for the Prevention and Treatment of HBV Infection. Ph.D. Thesis, University of the Witwatersrand, Johannesburg, South Africa, 2017. [Google Scholar]
- Zhang, C.; Maruggi, G.; Shan, H.; Li, J. Advances in MRNA Vaccines for Infectious Diseases. Front. Immunol. 2019, 10, 594. [Google Scholar] [CrossRef]
- Oǧuz, F.; Atmaca, H. MRNA as a Therapeutics: Understanding MRNA Vaccines. Adv. Pharm. Bull. 2022, 12, 274. [Google Scholar] [CrossRef]
- Gu, Y.Z.; Zhao, X.; Song, X. rong Ex Vivo Pulsed Dendritic Cell Vaccination against Cancer. Acta Pharmacol. Sin. 2020, 41, 959. [Google Scholar] [CrossRef]
- Malak, A.; Baronian, K.; Kunze, G. Blastobotrys (Arxula) Adeninivorans: A Promising Alternative Yeast for Biotechnology and Basic Research. Yeast 2016, 33, 535–547. [Google Scholar] [CrossRef]
- Huzair, F.; Sturdy, S. Biotechnology and the Transformation of Vaccine Innovation: The Case of the Hepatitis B Vaccines 1968–2000. Stud. Hist. Philos. Biol. Biomed. Sci. 2017, 64, 11. [Google Scholar] [CrossRef] [PubMed]
- Hardy, E.; Martínez, E.; Diago, D.; Díaz, R.; González, D.; Herrera, L. Large-Scale Production of Recombinant Hepatitis B Surface Antigen from Pichia Pastoris. J. Biotechnol. 2000, 77, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Wieland, S.F. The Chimpanzee Model for Hepatitis B Virus Infection. Cold Spring Harb. Perspect. Med. 2015, 5, a021469. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhou, X.; Zhou, Y.H. Hepatitis B Vaccine Development and Implementation. Hum. Vaccines Immunother. 2020, 16, 1533–1544. [Google Scholar] [CrossRef] [PubMed]
- Frey, S.; Dagan, R.; Ashur, Y.; Chen, X.Q.; Ibarra, J.; Kollaritsch, H.; Mazur, M.H.; Poland, G.A.; Reisinger, K.; Walter, E.; et al. Interference of Antibody Production to Hepatitis B Surface Antigen in a Combination Hepatitis A/Hepatitis B Vaccine. J. Infect. Dis. 1999, 180, 2018–2022. [Google Scholar] [CrossRef] [PubMed]
- Gaik, S.K.; Pujar, N.S.; Titchener-Hooker, N.J. Study of Detergent-Mediated Liberation of Hepatitis B Virus-like Particles from S. Cerevisiae Homogenate: Identifying a Framework for the Design of Future-Generation Lipoprotein Vaccine Processes. Biotechnol. Prog. 2008, 24, 623–631. [Google Scholar] [CrossRef]
- Jacobs, E.; Rutgers, T.; Voet, P.; Dewerchin, M.; Cabezon, T.; de Wilde, M. Simultaneous Synthesis and Assembly of Various Hepatitis B Surface Proteins in Saccharomyces Cerevisiae. Gene 1989, 80, 279–291. [Google Scholar] [CrossRef]
- Hadiji-Abbes, N.; Martin, M.; Benzina, W.; Karray-Hakim, H.; Gergely, C.; Gargouri, A.; Mokdad-Gargouri, R. Extraction and Purification of Hepatitis B Virus-like M Particles from a Recombinant Saccharomyces Cerevisiae Strain Using Alumina Powder. J. Virol. Methods 2013, 187, 132–137. [Google Scholar] [CrossRef]
- King, T.H.; Kemmler, C.B.; Guo, Z.; Mann, D.; Lu, Y.; Coeshott, C.; Gehring, A.J.; Bertoletti, A.; Ho, Z.Z.; Delaney, W.; et al. A Whole Recombinant Yeast-Based Therapeutic Vaccine Elicits HBV X, S and Core Specific T Cells in Mice and Activates Human T Cells Recognizing Epitopes Linked to Viral Clearance. PLoS ONE 2014, 9, e0101904. [Google Scholar] [CrossRef]
- Gaggar, A.; Coeshott, C.; Apelian, D.; Rodell, T.; Armstrong, B.R.; Shen, G.; Subramanian, G.M.; McHutchison, J.G. Safety, Tolerability and Immunogenicity of GS-4774, a Hepatitis B Virus-Specific Therapeutic Vaccine, in Healthy Subjects: A Randomized Study. Vaccine 2014, 32, 4925–4931. [Google Scholar] [CrossRef]
- Pleckaityte, M.; Bremer, C.M.; Gedvilaite, A.; Kucinskaite-Kodze, I.; Glebe, D.; Zvirbliene, A. Construction of Polyomavirus-Derived Pseudotype Virus-like Particles Displaying a Functionally Active Neutralizing Antibody against Hepatitis B Virus Surface Antigen. BMC Biotechnol. 2015, 15, 85. [Google Scholar] [CrossRef]
- Zhang, C.; Ku, Z.; Liu, Q.; Wang, X.; Chen, T.; Ye, X.; Li, D.; Jin, X.; Huang, Z. High-Yield Production of Recombinant Virus-like Particles of Enterovirus 71 in Pichia Pastoris and Their Protective Efficacy against Oral Viral Challenge in Mice. Vaccine 2015, 33, 2335–2341. [Google Scholar] [CrossRef]
- Shirbaghaee, Z.; Bolhassani, A. Different Applications of Virus-like Particles in Biology and Medicine: Vaccination and Delivery Systems. Biopolymers 2016, 105, 113–132. [Google Scholar] [CrossRef]
- Samandari, T.; Fiore, A.E.; Negus, S.; Williams, J.L.; Kuhnert, W.; McMahon, B.J.; Bell, B.P. Differences in Response to a Hepatitis B Vaccine Booster Dose among Alaskan Children and Adolescents Vaccinated during Infancy. Pediatrics 2007, 120, e373–e381. [Google Scholar] [CrossRef]
- Bisht, H.; Chugh, D.A.; Swaminathan, S.; Khanna, N. Expression and Purification of Dengue Virus Type 2 Envelope Protein as a Fusion with Hepatitis B Surface Antigen in Pichia Pastoris. Protein Expr. Purif. 2001, 23, 84–96. [Google Scholar] [CrossRef]
- Zahid, M.; Lünsdorf, H.; Rinas, U. Assessing Stability and Assembly of the Hepatitis B Surface Antigen into Virus-like Particles during down-Stream Processing. Vaccine 2015, 33, 3739–3745. [Google Scholar] [CrossRef]
- Freivalds, J.; Dislers, A.; Ose, V.; Pumpens, P.; Tars, K.; Kazaks, A. Highly Efficient Production of Phosphorylated Hepatitis B Core Particles in Yeast Pichia Pastoris. Protein Expr. Purif. 2011, 75, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Bian, G.; Cheng, Y.; Wang, Z.; Hu, Y.; Zhang, X.; Wu, M.; Chen, Z.; Shi, B.; Sun, S.; Shen, Y.; et al. Whole Recombinant Hansenula Polymorpha Expressing Hepatitis B Virus Surface Antigen (Yeast-HBsAg) Induces Potent HBsAg-Specific Th1 and Th2 Immune Responses. Vaccine 2009, 28, 187–194. [Google Scholar] [CrossRef]
- Gurramkonda, C.; Adnan, A.; Gäbel, T.; Lünsdorf, H.; Ross, A.; Nemani, S.K.; Swaminathan, S.; Khanna, N.; Rinas, U. Simple High-Cell Density Fed-Batch Technique for High-Level Recombinant Protein Production with Pichia Pastoris: Application to Intracellular Production of Hepatitis B Surface Antigen. Microb. Cell Fact. 2009, 8, 13. [Google Scholar] [CrossRef]
- Lünsdorf, H.; Gurramkonda, C.; Adnan, A.; Khanna, N.; Rinas, U. Virus-like Particle Production with Yeast: Ultrastructural and Immunocytochemical Insights into Pichia Pastoris Producing High Levels of the Hepatitis B Surface Antigen. Microb. Cell Fact. 2011, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Lin, Q.; Sun, Y.; Lu, X.; Qiu, Y.; Li, Y.; Guo, X. Expression, Purification, and Characterization of Hepatitis B Virus Surface Antigens (HBsAg) in Yeast Pichia Pastoris. Appl. Biochem. Biotechnol. 2009, 158, 432–444. [Google Scholar] [CrossRef]
- Caetano, K.A.A.; Del-Rios, N.H.A.; Pinheiro, R.S.; Bergamaschi, F.P.R.; Dos Santos Carneiro, M.A.; Teles, S.A. Low Immunogenicity of Recombinant Hepatitis B Vaccine Derived from Hansenula Polymorpha in Adults Aged Over 40 Years. Am. J. Trop. Med. Hyg. 2017, 96, 118–121. [Google Scholar] [CrossRef]
- Inoue, T.; Tanaka, Y. Cross-Protection of Hepatitis B Vaccination among Different Genotypes. Vaccines 2020, 8, 456. [Google Scholar] [CrossRef] [PubMed]
- Wiedermann, U.; Garner-Spitzer, E.; Wagner, A. Primary Vaccine Failure to Routine Vaccines: Why and What to Do? Hum. Vaccin. Immunother. 2016, 12, 239. [Google Scholar] [CrossRef] [PubMed]
- Elbahrawy, A.; Atalla, H.; Alboraie, M.; Alwassief, A.; Madian, A.; El Fayoumie, M.; Tabll, A.A.; Aly, H.H. Recent Advances in Protective Vaccines against Hepatitis Viruses: A Narrative Review. Viruses 2023, 15, 214. [Google Scholar] [CrossRef] [PubMed]
- Cooreman, M.P.; Leroux-Roels, G.; Paulij, W.P. Vaccine- and Hepatitis B Immune Globulin-Induced Escape Mutations of Hepatitis B Virus Surface Antigen. J. Biomed. Sci. 2001, 8, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Lok, A.S.; Pan, C.Q.; Han, S.H.B.; Trinh, H.N.; Fessel, W.J.; Rodell, T.; Massetto, B.; Lin, L.; Gaggar, A.; Subramanian, G.M.; et al. Randomized Phase II Study of GS-4774 as a Therapeutic Vaccine in Virally Suppressed Patients with Chronic Hepatitis B. J. Hepatol. 2016, 65, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Boni, C.; Janssen, H.L.A.; Rossi, M.; Yoon, S.K.; Vecchi, A.; Barili, V.; Yoshida, E.M.; Trinh, H.; Rodell, T.C.; Laccabue, D.; et al. Combined GS-4774 and Tenofovir Therapy Can Improve HBV-Specific T-Cell Responses in Patients with Chronic Hepatitis. Gastroenterology 2019, 157, 227–241.e7. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Song, H.; Li, J.; Wang, Y.; Yan, X.; Zhao, B.; Zhang, X.; Wang, S.; Chen, L.; Qiu, B.; et al. Hansenula Polymorpha Expressed Heat Shock Protein Gp96 Exerts Potent T Cell Activation Activity as an Adjuvant. J. Biotechnol. 2011, 151, 343–349. [Google Scholar] [CrossRef]
- Kutzler, M.A.; Weiner, D.B. DNA Vaccines: Ready for Prime Time? Nat. Rev. Genet. 2008 910 2008, 9, 776–788. [Google Scholar] [CrossRef]
- Tzeng, T.T.; Chai, K.M.; Shen, K.Y.; Yu, C.Y.; Yang, S.J.; Huang, W.C.; Liao, H.C.; Chiu, F.F.; Dou, H.Y.; Liao, C.L.; et al. A DNA Vaccine Candidate Delivered by an Electroacupuncture Machine Provides Protective Immunity against SARS-CoV-2 Infection. npj Vaccines 2022, 7, 60. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Liang, H.; Chen, P.; Li, Y.; Li, Z.; Fan, S.; Wu, K.; Li, X.; Chen, W.; Qin, Y.; et al. Viral Vector Vaccine Development and Application during the COVID-19 Pandemic. Microorganisms 2022, 10, 1450. [Google Scholar] [CrossRef]
- Tsounis, E.P.; Mouzaki, A.; Triantos, C. Nucleic Acid Vaccines: A Taboo Broken and Prospect for a Hepatitis B Virus Cure. World J. Gastroenterol. 2021, 27, 7005–7013. [Google Scholar] [CrossRef]
- Phillips, S.; Jagatia, R.; Chokshi, S. Novel Therapeutic Strategies for Chronic Hepatitis B. Virulence 2022, 13, 1111. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Jeong, S.W.; Jang, J.Y. Hepatitis B Virus Reactivation Associated with Therapeutic Interventions. Front. Med. 2022, 8, 2965. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z. DNA Vaccine. Adv. Genet. 2005, 54, 257. [Google Scholar] [CrossRef]
- Aleebrahim-Dehkordi, E.; Molavi, B.; Mokhtari, M.; Deravi, N.; Fathi, M.; Fazel, T.; Mohebalizadeh, M.; Koochaki, P.; Shobeiri, P.; Hasanpour-Dehkordi, A. T Helper Type (Th1/Th2) Responses to SARS-CoV-2 and Influenza A (H1N1) Virus: From Cytokines Produced to Immune Responses. Transpl. Immunol. 2022, 70, 101495. [Google Scholar] [CrossRef]
- Zhong, S.; Zhang, T.; Tang, L.; Li, Y. Cytokines and Chemokines in HBV Infection. Front. Mol. Biosci. 2021, 8, 1188. [Google Scholar] [CrossRef]
- Desombere, I.; Gijbels, Y.; Verwulgen, A.; Leroux-Roels, G. Characterization of the T Cell Recognition of Hepatitis B Surface Antigen (HBsAg) by Good and Poor Responders to Hepatitis B Vaccines. Clin. Exp. Immunol. 2000, 122, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Lang-Meli, J.; Neumann-Haefelin, C.; Thimme, R. Immunotherapy and Therapeutic Vaccines for Chronic HBV Infection. Curr. Opin. Virol. 2021, 51, 149–157. [Google Scholar] [CrossRef]
- Wang, X.Y.; Zhang, X.X.; Yao, X.; Jiang, J.H.; Xie, Y.H.; Yuan, Z.H.; Wen, Y.M. Serum HBeAg Sero-Conversion Correlated with Decrease of HBsAg and HBV DNA in Chronic Hepatitis B Patients Treated with a Therapeutic Vaccine. Vaccine 2010, 28, 8169–8174. [Google Scholar] [CrossRef]
- Xu, D.Z.; Wang, X.Y.; Shen, X.L.; Gong, G.Z.; Ren, H.; Guo, L.M.; Sun, A.M.; Xu, M.; Li, L.J.; Guo, X.H.; et al. Results of a Phase III Clinical Trial with an HBsAg-HBIG Immunogenic Complex Therapeutic Vaccine for Chronic Hepatitis B Patients: Experiences and Findings. J. Hepatol. 2013, 59, 450–456. [Google Scholar] [CrossRef]
- Ishikawa, T.; Kakumu, S. Combination Therapy with Lamivudine and HB Vaccine on Chronic Hepatitis B. Hepatol. Res. 2007, 37, S62–S66. [Google Scholar] [CrossRef]
- Le Hoa, P.T.; Huy, N.T.; Thu, L.T.; Nga, C.N.; Nakao, K.; Eguchi, K.; Chi, N.H.; Hoang, B.H.; Hirayama, K. Randomized Controlled Study Investigating Viral Suppression and Serological Response Following Pre-S1/Pre-S2/S Vaccine Therapy Combined with Lamivudine Treatment in HBeAg-Positive Patients with Chronic Hepatitis B. Antimicrob. Agents Chemother. 2009, 53, 5134–5140. [Google Scholar] [CrossRef]
- A Study to Evaluate the Efficacy and Safety of Therapeutic Hepatitis B Vaccine—Full Text View—ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04289987 (accessed on 16 February 2023).
- Lai, M.W.; Hsu, C.W.; Lin, C.L.; Chien, R.N.; Lin, W.R.; Chang, C.S.; Liang, K.H.; Yeh, C.T. Multiple Doses of Hepatitis B Recombinant Vaccine for Chronic Hepatitis B Patients with Low Surface Antigen Levels: A Pilot Study. Hepatol. Int. 2018, 12, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Bin Lee, Y.; Lee, J.H.; Kim, Y.J.; Yoon, J.H.; Lee, H.S. The Effect of Therapeutic Vaccination for the Treatment of Chronic Hepatitis B Virus Infection. J. Med. Virol. 2015, 87, 575–582. [Google Scholar] [CrossRef]
- Al Mahtab, M.; Akbar, S.M.F.; Aguilar, J.C.; Guillen, G.; Penton, E.; Tuero, A.; Yoshida, O.; Hiasa, Y.; Onji, M. Treatment of Chronic Hepatitis B Naïve Patients with a Therapeutic Vaccine Containing HBs and HBc Antigens (a Randomized, Open and Treatment Controlled Phase III Clinical Trial). PLoS ONE 2018, 13, e0201236. [Google Scholar] [CrossRef]
- Wei, L.; Zhao, T.; Zhang, J.; Mao, Q.; Gong, G.; Sun, Y.; Chen, Y.; Wang, M.; Tan, D.; Gong, Z.; et al. Efficacy and Safety of a Nanoparticle Therapeutic Vaccine in Patients with Chronic Hepatitis B: A Randomized Clinical Trial. Hepatology 2022, 75, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhou, X.; Bian, Y.; Wang, S.; Chai, Q.; Guo, Z.; Wang, Z.; Zhu, P.; Peng, H.; Yan, X.; et al. Dual-Targeting Nanoparticle Vaccine Elicits a Therapeutic Antibody Response against Chronic Hepatitis B. Nat. Nanotechnol. 2020, 15, 406–416. [Google Scholar] [CrossRef]
- Ke, X.; Howard, G.P.; Tang, H.; Cheng, B.; Saung, M.T.; Santos, J.L.; Mao, H.Q. Physical and Chemical Profiles of Nanoparticles for Lymphatic Targeting. Adv. Drug Deliv. Rev. 2019, 151–152, 72–93. [Google Scholar] [CrossRef]
- Qiao, D.; Liu, L.; Chen, Y.; Xue, C.; Gao, Q.; Mao, H.Q.; Leong, K.W.; Chen, Y. Potency of a Scalable Nanoparticulate Subunit Vaccine. Nano Lett. 2018, 18, 3007–3016. [Google Scholar] [CrossRef]
- Qiao, D.; Chen, Y.; Liu, L. Engineered Therapeutic Nanovaccine against Chronic Hepatitis B Virus Infection. Biomaterials 2021, 269, 120674. [Google Scholar] [CrossRef]
- Miao, J.; Gao, P.; Li, Q.; He, K.; Zhang, L.; Wang, J.; Huang, L. Advances in Nanoparticle Drug Delivery Systems for Anti-Hepatitis b Virus Therapy: A Narrative Review. Int. J. Mol. Sci. 2021, 22, 11227. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.M.; Simon, J.K.; Baker, J.R. Applications of Nanotechnology for Immunology. Nat. Rev. Immunol. 2013, 13, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Bruxvoort, K.; Sy, L.S.; Luo, Y.; Tseng, H.F. Real-World Evidence for Regulatory Decisions: Concomitant Administration of Zoster Vaccine Live and Pneumococcal Polysaccharide Vaccine. Am. J. Epidemiol. 2018, 187, 1856–1862. [Google Scholar] [CrossRef]
- Bridges, C.B.; Watson, T.L.; Nelson, N.P.; Chavez-Torres, M.; Fineis, P.; Ntiri-Reid, B.; Wake, E.; Leahy, J.M.; Kurian, A.K.; Hall, M.A.K.; et al. Challenges with Hepatitis B Vaccination of High Risk Adults—A Pilot Program. Vaccine 2019, 37, 5111–5120. [Google Scholar] [CrossRef] [PubMed]
- Trantham, L.; Kurosky, S.K.; Zhang, D.; Johnson, K.D. Adherence with and Completion of Recommended Hepatitis Vaccination Schedules among Adults in the United States. Vaccine 2018, 36, 5333–5339. [Google Scholar] [CrossRef]
- Bruxvoort, K.; Slezak, J.; Qian, L.; Sy, L.S.; Ackerson, B.; Reynolds, K.; Huang, R.; Solano, Z.; Towner, W.; Mercado, C.; et al. Association Between 2-Dose vs. 3-Dose Hepatitis B Vaccine and Acute Myocardial Infarction. JAMA 2022, 327, 1260–1268. [Google Scholar] [CrossRef]
- Jackson, S.; Lentino, J.; Kopp, J.; Murray, L.; Ellison, W.; Rhee, M.; Shockey, G.; Akella, L.; Erby, K.; Heyward, W.L.; et al. Immunogenicity of a Two-Dose Investigational Hepatitis B Vaccine, HBsAg-1018, Using a Toll-like Receptor 9 Agonist Adjuvant Compared with a Licensed Hepatitis B Vaccine in Adults. Vaccine 2018, 36, 668–674. [Google Scholar] [CrossRef]
- Li, L.; Barry, V.; Daffis, S.; Niu, C.; Huntzicker, E.; French, D.M.; Mikaelian, I.; Lanford, R.E.; Delaney, W.E.; Fletcher, S.P. Anti-HBV Response to Toll-like Receptor 7 Agonist GS-9620 Is Associated with Intrahepatic Aggregates of T Cells and B Cells. J. Hepatol. 2018, 68, 912–921. [Google Scholar] [CrossRef]
- Boni, C.; Barili, V.; Acerbi, G.; Rossi, M.; Vecchi, A.; Laccabue, D.; Penna, A.; Missale, G.; Ferrari, C.; Fisicaro, P. HBV Immune-Therapy: From Molecular Mechanisms to Clinical Applications. Int. J. Mol. Sci. 2019, 20, 2754. [Google Scholar] [CrossRef]
- Aguilar, J.C.; Aguiar, J.A.; Akbar, S.M.F. Action Mechanisms and Scientific Rationale of Using Nasal Vaccine (HeberNasvac) for the Treatment of Chronic Hepatitis B. Vaccines 2022, 10, 2087. [Google Scholar] [CrossRef] [PubMed]
- Zoulim, F.; Fournier, C.; Habersetzer, F.; Sprinzl, M.; Pol, S.; Coffin, C.S.; Leroy, V.; Ma, M.; Wedemeyer, H.; Lohse, A.W.; et al. Safety and Immunogenicity of the Therapeutic Vaccine TG1050 in Chronic Hepatitis B Patients: A Phase 1b Placebo-Controlled Trial. Hum. Vaccin. Immunother. 2020, 16, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Lok, A.S.; Zoulim, F.; Dusheiko, G.; Ghany, M.G. Hepatitis B Cure: From Discovery to Regulatory Approval. J. Hepatol. 2017, 67, 847–861. [Google Scholar] [CrossRef] [PubMed]
- Gehring, A.J.; Protzer, U. Targeting Innate and Adaptive Immune Responses to Cure Chronic HBV Infection. Gastroenterology 2019, 156, 325–337. [Google Scholar] [CrossRef]
- Phase I Study of INO-1800 with or without INO-9112 + EP in Chronic Hepatitis B Subjects—Full Text View—ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/NCT02431312 (accessed on 16 February 2023).





| Vaccine Generation | Type | Year | References |
|---|---|---|---|
| First hepatitis B Vaccine | Heat-treated form of the virus. | 1971 | [33] |
| I generation Hepatitis B Vaccine | Plasma-derived hepatitis B vaccine | 1981 | [34] |
| II generation Recombinant Vaccine | HBV DNA vaccine, expressed in yeast | 1986 | [34,35] |
| III generation Recombinant Vaccine | pre-S/S vaccines expressed in mammalian cells | 1990 | [34,35] |
| IV generation Recombinant Vaccine | Recombinant HBV vaccines with adjuvant (AS04, CpG) | 2005 | [36] |
| Vaccine | Adjuvant | Dose | Age | Administration | Reference |
|---|---|---|---|---|---|
| FENDRIX | 3-O-desacyl-4′-monophosphoryl lipid A and aluminum phosphate. | Four doses. (There should be a gap of 1 month between the first and second, and between the second and third injections. The fourth injection is given 4 months after the third.) | 15 years onwards. | Intramuscularly | [44] |
| ENGERIX B | Aluminum hydroxide | Three shots over a 6-month period. | Used in both pediatrics, starting with infants at birth, and adults. | Intramuscularly | [45] |
| HBVAXPRO | Amorphous aluminum hydroxyphosphate sulfate | At least three doses. | From birth through to 15 years of age. | Intramuscularly | [44] |
| HEPLISAV | Cytosine phosphoguanosine (CpG) 1018 adjuvant (HepB-CpG) | Only two doses. | Adults aged 18 years and older. | Intramuscularly | [46] |
| Vaccine Type | Administered Regime | Adjuvant | Results Summary | Trial Phase | Trial Registration | Reference |
|---|---|---|---|---|---|---|
| HBsAg-HBIG | Breaking immune tolerance to HBV by modulating viral antigen processing and presentation | Alum | Serum HBV DNA decreased and normalization of liver function | III | NCT03575208 (https://classic.clinicaltrials.gov/ct2/history/NCT03575208. Accessed date: 15 February 2023) | [129] |
| GS-4774 | Yeast-derived vaccine, includes HBsAg, HBcAg and hepatitis B X | Yeast component has been shown to have adjuvant properties and to reduce frequency and inhibitory activity of T regulatory cells | Activate an HBV-specific T cell immune response to reduce the number of cells containing HBV | II | NCT01943799 (https://clinicaltrials.gov/study/NCT01943799. Accessed date: 16 February 2023) | [115] |
| GS-9620 | Orally active small molecule agonist of toll-like receptor 7 (TLR7) | TLR Agonists as Vaccine Adjuvants | Serum viral DNA and antigens were suppressed for an extended period of time | I | NCT01590654 (https://www.cdek.liu.edu/trial/NCT01590654. Accessed date: 16 February 2023) | [148] |
| Theravax (DV-601) | HBV surface antigen (HBsAg) and HBV core antigen (HcAg) | Saponin-based ISCOMATRIX adjuvant | The development of an HBV-specific interferon- γ T-cell response, an HBV-specific lymphoproliferative response, and a decrease in HBV DNA | Ib | NCT01023230 (https://clinicaltrials.gov/study/NCT01023230. Accessed date: 15 February 2023) | [149] |
| Nasvac | Hepatitis B surface antigen (HBsAg) and hepatitis B core antigen (HBcAg) | Without adjuvants | After a multi-TLR agonist action, activate several innate immune and signal transduction pathways | III | NCT01374308 (https://clinicaltrials.gov/study/NCT01374308. Accessed date: 16 February 2023) | [150] |
| INO-1800 | DNA vaccine encoding HBsAg and a consensus sequence of HBcAg | Without adjuvants | Determines a virus-specific T-cell immune response | I | NCT02431312 (https://clinicaltrials.gov/study/NCT02431312. Accessed date: 16 February 2023) | |
| TG-1050 | Adenovirus 5-based therapeutic vaccine expressing core, polymerase, and surface antigen HBV proteins | Without adjuvants | Capable of inducing HBV-specific cellular immune response and IFN- γ producing T-cells targeting 1 to 3 encoded antigens | 1/1b | NCT02428400 (https://clinicaltrials.gov/study/NCT02428400. Accessed date: 19 February 2023) | [151] |
| HB-110 | 2nd-generation therapeutic adenoviral-based DNA vaccine encoding S, L, core, polymerase protein | Human IL-12 as adjuvant | Sustained CD4+ memory T cell responses were produced by long-lasting viral suppression and clear T-cell responses, particularly those that were memory-related | 1/2a | NCT01641536 (https://clinicaltrials.gov/study/NCT01641536. Accessed date: 26 July 2023) | [120] |
| CVI-HBV-002 | Activate the patient’s immune system effectively to fight and control the virus infection | Highly immunogenic L-HBsAg and powerful adjuvant L-pampo | Immune response was induced under the immune tolerance status and HBsAg particles in the blood were decreased by the antibody induced | II | NCT04289987 (https://clinicaltrials.gov/study/NCT04289987. Accessed date: 16 February 2023) | [132] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmood, F.; Xu, R.; Awan, M.U.N.; Song, Y.; Han, Q.; Xia, X.; Wei, J.; Xu, J.; Peng, J.; Zhang, J. HBV Vaccines: Advances and Development. Vaccines 2023, 11, 1862. https://doi.org/10.3390/vaccines11121862
Mahmood F, Xu R, Awan MUN, Song Y, Han Q, Xia X, Wei J, Xu J, Peng J, Zhang J. HBV Vaccines: Advances and Development. Vaccines. 2023; 11(12):1862. https://doi.org/10.3390/vaccines11121862
Chicago/Turabian StyleMahmood, Faisal, Ruixian Xu, Maher Un Nisa Awan, Yuzhu Song, Qinqin Han, Xueshan Xia, Jia Wei, Jun Xu, Juan Peng, and Jinyang Zhang. 2023. "HBV Vaccines: Advances and Development" Vaccines 11, no. 12: 1862. https://doi.org/10.3390/vaccines11121862