
Oral Immunization with Recombinant Saccharomyces cerevisiae Expressing Viral Capsid Protein 2 of Infectious Bursal Disease Virus Induces Unique Specific Antibodies and Protective Immunity
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Plasmids
2.2. Construction of Recombinant Yeast Strain Expressing VP2
2.3. Western Blotting Analysis
2.4. Quantitative Real-Time (qRT)-PCR
2.5. Immunofluorescence Assay (IFA)
2.6. Animal Immunization
2.7. Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. Statistical Analysis
3. Results
3.1. Construction of the Recombinant S. cerevisiae Strain ST1814G/Aga2-VP2
3.2. The Fermentation Kinetics of Recombinant ST1814G/Aga2-VP2 Strain
3.3. Health Status of the Mice Post Oral Immunization
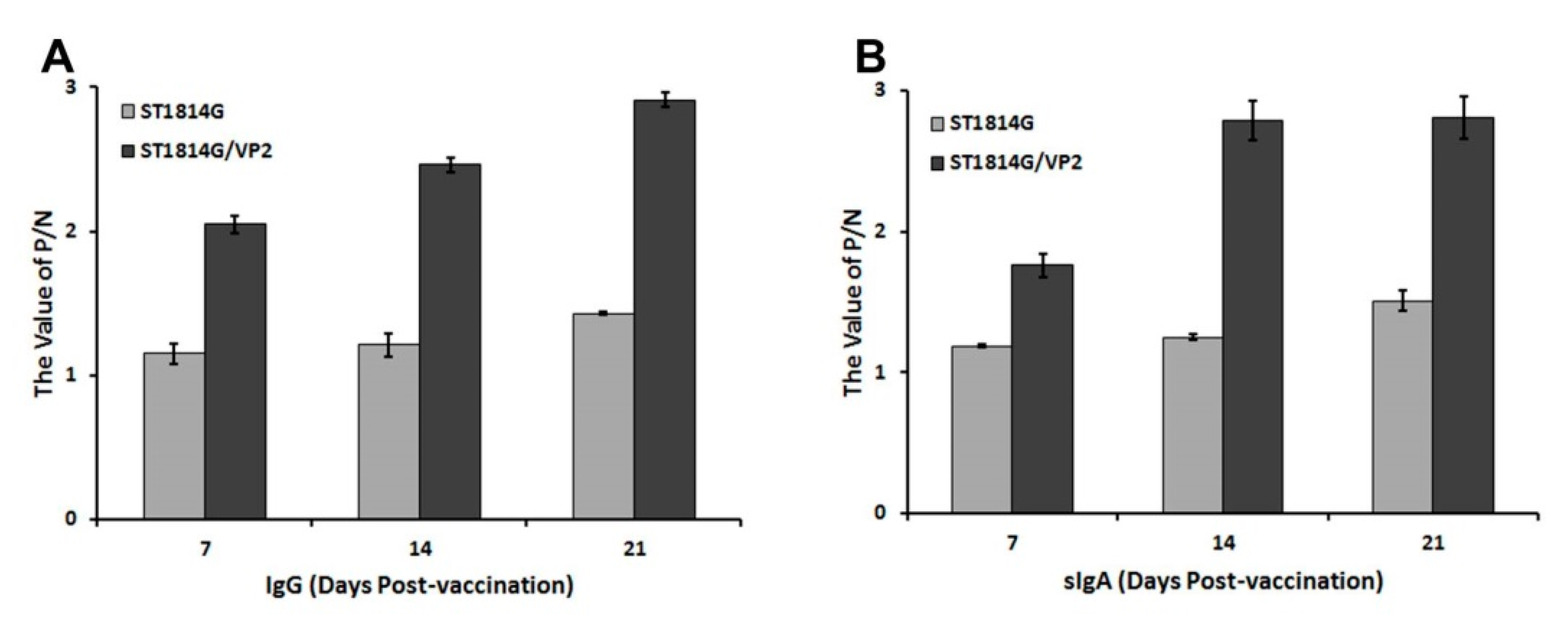
3.4. Antibody Levels of IgG and sIgA Post Oral Immunization
3.5. The Expression Patterns of Cytokines in Spleen and Thymus of the Immunized Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Deorao, C.V.; Rajasekhar, R.; Ravishankar, C.; Nandhakumar, D.; Sumod, K.; Palekkodan, H.; John, K.; Chaithra, G. Genetic variability in VP1 gene of infectious bursal disease virus from the field outbreaks of Kerala, India. Trop. Anim. Health Prod. 2021, 53, 407. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.L.; Wang, X.M.; Gao, Y.L.; Fu, C.Y. Direct evidence of reassortment and mutant spectrum analysis of a very virulent infectious bursal disease virus. Avian Dis. 2007, 51, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Palka, A.P.G.; Assunção de Matos, T.R.; de Souza, C.; Eugênio, D.S.; Krieger, M.A.; Fragoso, S.P.; Pavoni, D.P. Assessing the antigenicity of different VP3 regions of infectious bursal disease virus in chickens from South Brazil. BMC Vet. Res. 2021, 17, 259. [Google Scholar] [CrossRef] [PubMed]
- Ingrao, F.; Rauw, F.; Lambrecht, B.; van den Berg, T. Infectious Bursal Disease: A complex host-pathogen interaction. Dev. Comp. Immunol. 2013, 41, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Müller, H.; Schnitzler, D.; Bernstein, F.; Becht, H.; Cornelissen, D.; Lütticken, D.H. Infectious bursal disease of poultry: Antigenic structure of the virus and control. Vet. Microbiol. 1992, 33, 175–183. [Google Scholar] [CrossRef]
- Ferrero, D.; Garriga, D.; Navarro, A.; Rodríguez, J.F.; Verdaguer, N. Infectious Bursal Disease Virus VP3 Upregulates VP1-Mediated RNA-Dependent RNA Replication. J. Virol. 2015, 89, 11165–11168. [Google Scholar] [CrossRef]
- Ganguly, B.; Rastogi, S.K. Structural and functional modeling of viral protein 5 of Infectious Bursal Disease Virus. Virus Res. 2018, 247, 55–60. [Google Scholar] [CrossRef]
- Qiao, Q.; Song, M.; Song, C.; Zhang, Y.; Wang, X.; Huang, Q.; Wang, B.; Yang, P.; Zhao, S.; Li, Y.; et al. Single-Dose Vaccination of Recombinant Chimeric Newcastle Disease Virus (NDV) LaSota Vaccine Strain Expressing Infectious Bursal Disease Virus (IBDV) VP2 Gene Provides Full Protection against Genotype VII NDV and IBDV Challenge. Vaccines 2021, 9, 1483. [Google Scholar] [CrossRef]
- Zafar, M.; Shah, M.A.; Shehzad, A.; Tariq, A.; Habib, M.; Muddassar, M.; Shah, M.S.; Iqbal, M.; Hemmatzadeh, F.; Rahman, M. Characterization of the highly immunogenic VP2 protrusion domain as a diagnostic antigen for members of Birnaviridae family. Appl. Microbiol. Biotechnol. 2020, 104, 3391–3402. [Google Scholar] [CrossRef]
- Saugar, I.; Luque, D.; Oña, A.; Rodríguez, J.F.; Carrascosa, J.L.; Trus, B.L.; Castón, J.R. Structural polymorphism of the major capsid protein of a double-stranded RNA virus: An amphipathic alpha helix as a molecular switch. Structure 2005, 13, 1007–1017. [Google Scholar] [CrossRef]
- Eterradossi, N.; Toquin, D.; Rivallan, G.; Guittet, M. Modified activity of a VP2-located neutralizing epitope on various vaccine, pathogenic and hypervirulent strains of infectious bursal disease virus. Arch. Virol. 1997, 142, 255–270. [Google Scholar] [CrossRef]
- Shaw, I.; Davison, T.F. Protection from IBDV-induced bursal damage by a recombinant fowlpox vaccine, fpIBD1, is dependent on the titre of challenge virus and chicken genotype. Vaccine 2000, 18, 3230–3241. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; An, Q.; Song, S.; Gao, D.; Ping, W. Construction of Recombinant Baculoviruses Expressing Infectious Bursal Disease Virus Main Protective Antigen and Their Immune Effects on Chickens. PLoS ONE 2015, 10, e0132993. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Shan, S.; Wang, S.; Zhang, H.; Ma, L.; Hu, L.; Huang, H.; Wei, K.; Zhu, R. Fused IgY Fc and Polysaccharide Adjuvant Enhanced the Immune Effect of the Recombinant VP2 and VP5 Subunits-A Prospect for Improvement of Infectious Bursal Disease Virus Subunit Vaccine. Front. Microbiol. 2017, 8, 2258. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.K.; Sung, C.H.; Hsieh, P.F.; Hsiao, P.F.; Wu, B.Y.; Chou, C.C. Identification and characterization of infectious bursal disease virus subviral particles by capillary zone electrophoresis: Potential application for vaccine production and quality control. Poult. Sci. 2019, 98, 1658–1663. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, S.N.; Prince, P.R.; Madhumathi, J.; Roy, P.; Narayanan, R.B.; Antony, U. Protective immune responses of recombinant VP2 subunit antigen of infectious bursal disease virus in chickens. Vet. Immunol. Immunopathol. 2012, 148, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Longet, S.; Lundahl, M.L.E.; Lavelle, E.C. Targeted Strategies for Mucosal Vaccination. Bioconjug. Chem. 2018, 29, 613–623. [Google Scholar] [CrossRef]
- Wu, H.; Singh, N.K.; Locy, R.D.; Scissum-Gunn, K.; Giambrone, J.J. Immunization of chickens with VP2 protein of infectious bursal disease virus expressed in Arabidopsis thaliana. Avian Dis. 2004, 48, 663–668. [Google Scholar] [CrossRef]
- Wu, J.; Yu, L.; Li, L.; Hu, J.; Zhou, J.; Zhou, X. Oral immunization with transgenic rice seeds expressing VP2 protein of infectious bursal disease virus induces protective immune responses in chickens. Plant Biotechnol. J. 2007, 5, 570–578. [Google Scholar] [CrossRef]
- Kumar, R.; Kumar, P. Yeast-based vaccines: New perspective in vaccine development and application. FEMS Yeast Res. 2019, 19, foz007. [Google Scholar] [CrossRef]
- Patterson, R.; Eley, T.; Browne, C.; Martineau, H.M.; Werling, D. Oral application of freeze-dried yeast particles expressing the PCV2b Cap protein on their surface induce protection to subsequent PCV2b challenge in vivo. Vaccine 2015, 33, 6199–6205. [Google Scholar] [CrossRef]
- Bal, J.; Jung, H.Y.; Nguyen, L.N.; Park, J.; Jang, Y.S.; Kim, D.H. Evaluation of cell-surface displayed synthetic consensus dengue EDIII cells as a potent oral vaccine candidate. Microb. Cell Fact. 2018, 17, 146. [Google Scholar] [CrossRef]
- Upadhyaya, B.; Manjunath, R. Baker’s yeast expressing the Japanese encephalitis virus envelope protein on its cell surface: Induction of an antigen-specific but non-neutralizing antibody response. Yeast 2009, 26, 383–397. [Google Scholar] [CrossRef]
- Neutra, M.R.; Kozlowski, P.A. Mucosal vaccines: The promise and the challenge. Nat. Rev. Immunol. 2006, 6, 148–158. [Google Scholar] [CrossRef]
- Guo, Y.; Dong, J.; Zhou, T.; Auxillos, J.; Li, T.; Zhang, W.; Wang, L.; Shen, Y.; Luo, Y.; Zheng, Y.; et al. YeastFab: The design and construction of standard biological parts for metabolic engineering in Saccharomyces cerevisiae. Nucleic Acids Res. 2015, 43, e88. [Google Scholar] [CrossRef]
- Ganesh, K.; Raghavan, R. Hydropericardium hepatitis syndrome of broiler poultry: Current status of research. Res. Vet. Sci. 2000, 68, 201–206. [Google Scholar] [CrossRef]
- Shi, P.; Guo, Y.; Su, Y.; Zhu, M.; Fu, Y.; Chi, H.; Wu, J.; Huang, J. SUMOylation of DDX39A Alters Binding and Export of Antiviral Transcripts to Control Innate Immunity. J. Immunol. 2020, 205, 168–180. [Google Scholar] [CrossRef]
- Cao, H.; Hua, D.; Zhang, H.; Zhang, H.; Liu, N.; Feng, Z.; Li, H.; Zhao, B.; Zhang, L.; Guo, Y.; et al. Oral immunization of recombinant Saccharomyces cerevisiae expressing fiber-2 of fowl adenovirus serotype 4 induces protective immunity against homologous infection. Vet. Microbiol. 2022, 271, 109490. [Google Scholar] [CrossRef]
- Spackman, E.; Stephens, C.B.; Pantin-Jackwood, M.J. The Effect of Infectious Bursal Disease Virus-Induced Immunosuppression on Vaccination Against Highly Pathogenic Avian Influenza Virus. Avian Dis. 2018, 62, 36–44. [Google Scholar] [CrossRef]
- Parker, D.; de Wit, S.; Houghton, H.; Prandini, F. Assessment of impact of a novel infectious bursal disease (IBD) vaccination programme in breeders on IBD humoral antibody levels through the laying period. Vet. Rec. Open 2014, 1, e000016. [Google Scholar] [CrossRef]
- Banda, A.; Villegas, P.; Purvis, L.B.; Perozo, F. Protection conferred by coarse spray vaccination against challenge with infectious bursal disease virus in commercial broilers. Avian Dis. 2008, 52, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, D.J. Multivalent virus-like-particle vaccine protects against classic and variant infectious bursal disease viruses. Avian Dis. 2013, 57, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Saugar, I.; Irigoyen, N.; Luque, D.; Carrascosa, J.L.; Rodríguez, J.F.; Castón, J.R. Electrostatic interactions between capsid and scaffolding proteins mediate the structural polymorphism of a double-stranded RNA virus. J. Biol. Chem. 2010, 285, 3643–3650. [Google Scholar] [CrossRef] [PubMed]
- Rage, E.; Drissi Touzani, C.; Marusic, C.; Lico, C.; Göbel, T.; Bortolami, A.; Bonfante, F.; Salzano, A.M.; Scaloni, A.; Fellahi, S.; et al. Functional characterization of a plant-produced infectious bursal disease virus antigen fused to the constant region of avian IgY immunoglobulins. Appl. Microbiol. Biotechnol. 2019, 103, 7491–7504. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Liu, Y.; Wang, A.; Zhang, G.; Yang, G.; Chen, Y.; Ji, P.; Liu, C.; Song, Y.; Su, Y.; et al. High level soluble expression and one-step purification of IBDV VP2 protein in Escherichia coli. Biotechnol. Lett. 2016, 38, 901–908. [Google Scholar] [CrossRef]
- Austriaco, N. Yeast oral vaccines against infectious diseases. Front. Microbiol. 2023, 14, 1150412. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yao, L.; Guo, Y.; Li, X.; Ma, L.; Sun, R.; Han, X.; Liu, J.; Huang, J. Oral SARS-CoV-2 Spike Protein Recombinant Yeast Candidate Prompts Specific Antibody and Gut Microbiota Reconstruction in Mice. Front. Microbiol. 2022, 13, 792532. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Z.; Zhang, H.; Guo, Y.; Zhang, X.; Zhang, L.; Yang, L.; Li, S.; Li, C.; Cui, D.; et al. Recombinant hemagglutinin displaying on yeast reshapes congenital lymphocyte subsets to prompt optimized systemic immune protection against avian influenza infection. Front. Microbiol. 2023, 14, 1153922. [Google Scholar] [CrossRef]
- Brown, G.D.; Gordon, S. Fungal beta-glucans and mammalian immunity. Immunity 2003, 19, 311–315. [Google Scholar] [CrossRef]
- Wang, L.; Yang, M.; Luo, S.; Yang, G.A.-O.; Lu, X.; Lu, J.A.-O.; Chen, J.A.-O.X. Oral Vaccination of Recombinant Saccharomyces cerevisiae Expressing ORF132 Induces Protective Immunity against Cyprinid Herpesvirus-2. Vaccines 2023, 11, 186. [Google Scholar] [CrossRef]
- Macioła, A.K.; Pietrzak, M.A.; Kosson, P.; Czarnocki-Cieciura, M.; Śmietanka, K.; Minta, Z.; Kopera, E. The Length of N-Glycans of Recombinant H5N1 Hemagglutinin Influences the Oligomerization and Immunogenicity of Vaccine Antigen. Front. Immunol. 2017, 8, 444. [Google Scholar] [CrossRef]
- Wanzeck, K.; Boyd, K.L.; McCullers, J.A. Glycan shielding of the influenza virus hemagglutinin contributes to immunopathology in mice. Am. J. Respir. Crit. Care Med. 2011, 183, 767–773. [Google Scholar] [CrossRef]
- García, G.; Dogi, C.; de Moreno de LeBlanc, A.; Greco, C.; Cavaglieri, L. Gut-borne Saccharomyces cerevisiae, a promising candidate for the formulation of feed additives, modulates immune system and gut microbiota. Benef. Microbes 2016, 7, 659–668. [Google Scholar] [CrossRef]
- Shah, A.U.; Li, Y.; Ouyang, W.; Wang, Z.; Zuo, J.; Shi, S.; Yu, Q.; Lin, J.; Yang, Q. From nasal to basal: Single-cell sequencing of the bursa of Fabricius highlights the IBDV infection mechanism in chickens. Cell Biosci. 2021, 11, 212. [Google Scholar] [CrossRef]
- Wang, H.; Li, W.; Zheng, S.J. Advances on Innate Immune Evasion by Avian Immunosuppressive Viruses. Front. Immunol. 2022, 13, 901913. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Sato, S.; Ishii, K.J.; Coban, C.; Hemmi, H.; Yamamoto, M.; Terai, K.; Matsuda, M.; Inoue, J.; Uematsu, S.; et al. Interferon-alpha induction through Toll-like receptors involves a direct interaction of IRF7 with MyD88 and TRAF6. Nat. Immunol. 2004, 5, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Yanai, H.; Mizutani, T.; Negishi, H.; Shimada, N.; Suzuki, N.; Ohba, Y.; Takaoka, A.; Yeh, W.C.; Taniguchi, T. Role of a transductional-transcriptional processor complex involving MyD88 and IRF-7 in Toll-like receptor signaling. Proc. Natl. Acad. Sci. USA 2004, 101, 15416–15421. [Google Scholar] [CrossRef] [PubMed]
- Kaminuma, O.; Elly, C.; Tanaka, Y.; Mori, A.; Liu, Y.C.; Altman, A.; Miyatake, S. Vav-induced activation of the human IFN-gamma gene promoter is mediated by upregulation of AP-1 activity. FEBS Lett. 2002, 514, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Paul, W.E. CD4 T cells: Fates, functions, and faults. Blood 2008, 112, 1557–1569. [Google Scholar] [CrossRef] [PubMed]
- Ruterbusch, M.; Pruner, K.B.; Shehata, L.; Pepper, M. In Vivo CD4(+) T Cell Differentiation and Function: Revisiting the Th1/Th2 Paradigm. Annu. Rev. Immunol. 2020, 38, 705–725. [Google Scholar] [CrossRef]
- Mosmann, T.R.; Coffman, R.L. TH1 and TH2 cells: Different patterns of lymphokine secretion lead to different functional properties. Annu. Rev. Immunol. 1989, 7, 145–173. [Google Scholar] [CrossRef] [PubMed]
- Paul, W.E.; Seder, R.A. Lymphocyte responses and cytokines. Cell 1994, 76, 241–251. [Google Scholar] [CrossRef]
- Weaver, C.T.; Harrington, L.E.; Mangan, P.R.; Gavrieli, M.; Murphy, K.M. Th17: An effector CD4 T cell lineage with regulatory T cell ties. Immunity 2006, 24, 677–688. [Google Scholar] [CrossRef]
- Sakaguchi, S. Naturally arising CD4+ regulatory t cells for immunologic self-tolerance and negative control of immune responses. Annu. Rev. Immunol. 2004, 22, 531–562. [Google Scholar] [CrossRef]
- Krause, P.; Morris, V.; Greenbaum, J.A.; Park, Y.; Bjoerheden, U.; Mikulski, Z.; Muffley, T.; Shui, J.W.; Kim, G.; Cheroutre, H.; et al. IL-10-producing intestinal macrophages prevent excessive antibacterial innate immunity by limiting IL-23 synthesis. Nat. Commun. 2015, 6, 7055. [Google Scholar] [CrossRef] [PubMed]
- Mellor, A.L.; Munn, D.H. IDO expression by dendritic cells: Tolerance and tryptophan catabolism. Nat. Rev. Immunol. 2004, 4, 762–774. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Genbank Accession Number | Primer Sequence |
|---|---|---|
| β-actin-F | NM_007393.5 | AGAGGGAAATCGTGCGTGAC |
| β-actin-R | CAATAGTGATGACCTGGCCGT | |
| VP2-F1 | KT381974.1 | CTACACTATAACTGCAGCCGAT |
| VP2-R1 | CGCAGTCCCATCAAAGCCTA | |
| IL-2-F | NM_008366.3 | GTGCTCCTTGTCAACAGCG |
| IL-2-R | GGGGAGTTTCAGGTTCCTGTA | |
| IL-10-F | NM_010548.2 | GGTTGCCAAGCCTTATCGGA |
| IL-10-R | ACCTGCTCCACTGCCTTGCT | |
| IFN-α-F | NM_010503.2 | CTAGACTCTGTGCTTTCCTCGT |
| IFN-α-R | ATCGCATCCTAGAGAACAGGT | |
| IFN-γ-F | NM_008337.4 | ATGAACGCTACACACTGCATC |
| IFN-γ-R | CCATCCTTTTGCCAGTTCCTC | |
| MyD88-F | NM_010851.3 | GAGATGATCCGGCAACTAGAAC |
| MyD88-R | GTCCTTCTTCATCGCCTTGTAT | |
| TGF-β-F | NM_011577.2 | CTCCCGTGGCTTCTAGTGC |
| TGF-β-R | GCCTTAGTTTGGACAGGATCTG | |
| TLR3-F | NM_001357316.1 | AGTACAACAATATACAGCGTCT |
| TLR3-R | TGCTTAGTAAATGCTCGCTTC | |
| Gata3-F | NM_008091.3 | CTGGAGGAGGAACGCTAATG |
| Gata3-R | GATGACATGTGTCTGGAGAGAG | |
| Foxp3-F | NM_001199347.1 | GTGGCCTCAATGGACAAGA |
| Foxp3-R | AAGGGTGGCATAGGTGAAAG | |
| TBX21-F | NM_019507.2 | CACATCGTGGAGGTGAATGA |
| TBX21-R | CTTCTCACCTCTTCTATCCAACC | |
| RORC-F | NM_001293734.1 | CGGAGCAGACACACTTACATAC |
| RORC-R | CTTTGCCTCGTTCTGGACTATAC | |
| IDO-F | M69109.1 | CCTGCCTCCTATTCTGTCTTATG |
| IDO-R | ACATAGCTACCAGTCTGGAGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Hua, D.; Qu, Q.; Cao, H.; Feng, Z.; Liu, N.; Huang, J.; Zhang, L. Oral Immunization with Recombinant Saccharomyces cerevisiae Expressing Viral Capsid Protein 2 of Infectious Bursal Disease Virus Induces Unique Specific Antibodies and Protective Immunity. Vaccines 2023, 11, 1849. https://doi.org/10.3390/vaccines11121849
Li H, Hua D, Qu Q, Cao H, Feng Z, Liu N, Huang J, Zhang L. Oral Immunization with Recombinant Saccharomyces cerevisiae Expressing Viral Capsid Protein 2 of Infectious Bursal Disease Virus Induces Unique Specific Antibodies and Protective Immunity. Vaccines. 2023; 11(12):1849. https://doi.org/10.3390/vaccines11121849
Chicago/Turabian StyleLi, Huliang, Deping Hua, Qingxia Qu, Hongwei Cao, Zhehan Feng, Na Liu, Jinhai Huang, and Lei Zhang. 2023. "Oral Immunization with Recombinant Saccharomyces cerevisiae Expressing Viral Capsid Protein 2 of Infectious Bursal Disease Virus Induces Unique Specific Antibodies and Protective Immunity" Vaccines 11, no. 12: 1849. https://doi.org/10.3390/vaccines11121849