
Comparative Epidemiological Assessment of Monkeypox Infections on a Global and Continental Scale Using Logistic and Gompertz Mathematical Models
 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Mathematical Modeling
2.2.1. Logistic Model
2.2.2. Gompertz Model
2.3. Statistical Analysis
2.3.1. Normality Tests for the Variable Cases
2.3.2. Kruskal–Wallis Test for Monkeypox Cases Velocity
2.3.3. Post-Hoc Test: Pairwise Wilcoxon Test for Monkeypox Cases Velocity
2.3.4. Multiple Linear Regression Analysis for Monkeypox Cases Velocity
2.3.5. Comparison of Modeled Variable against Real Data
2.4. Estimation of the Basic Reproduction Number
3. Results
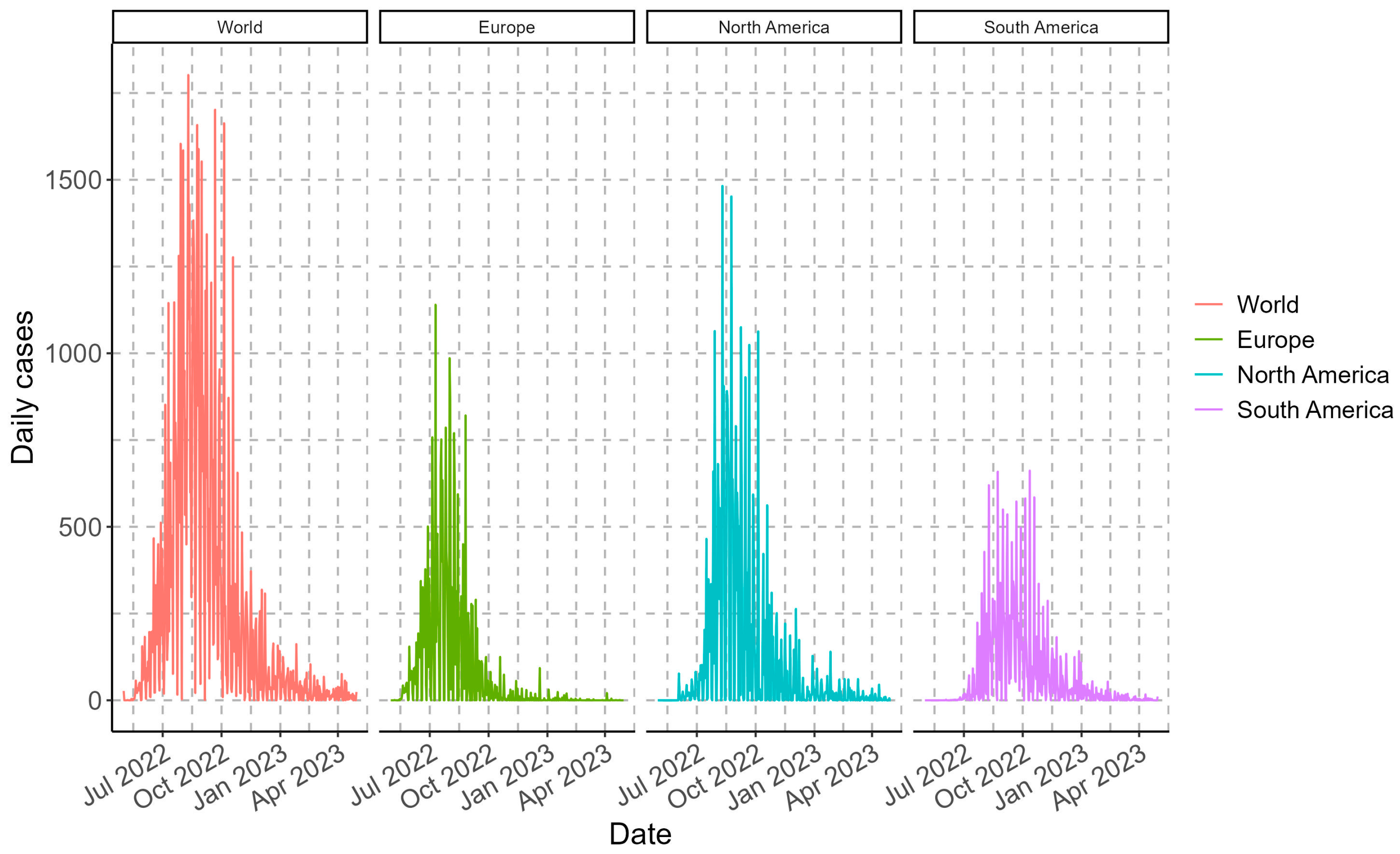
3.1. Epidemiological Panorama of Monkeypox
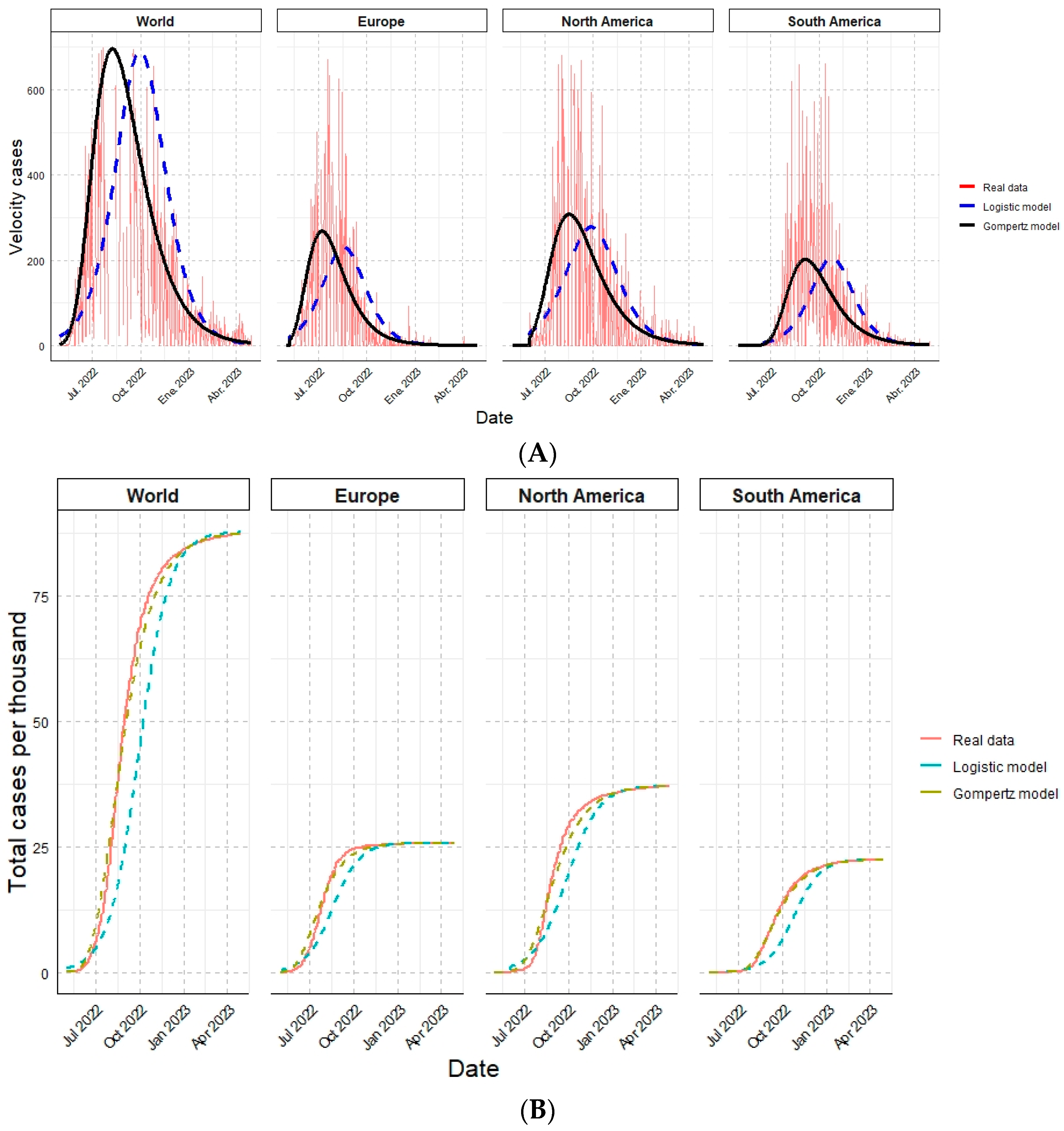
3.2. Mathematical Modeling
3.3. Statistical Analysis
3.3.1. Normality Tests for the Variable Cases
3.3.2. Kruskal–Wallis Test for Monkeypox Cases Velocity
3.3.3. Post-Hoc Test: Pairwise Wilcoxon Test for Monkeypox Cases Velocity
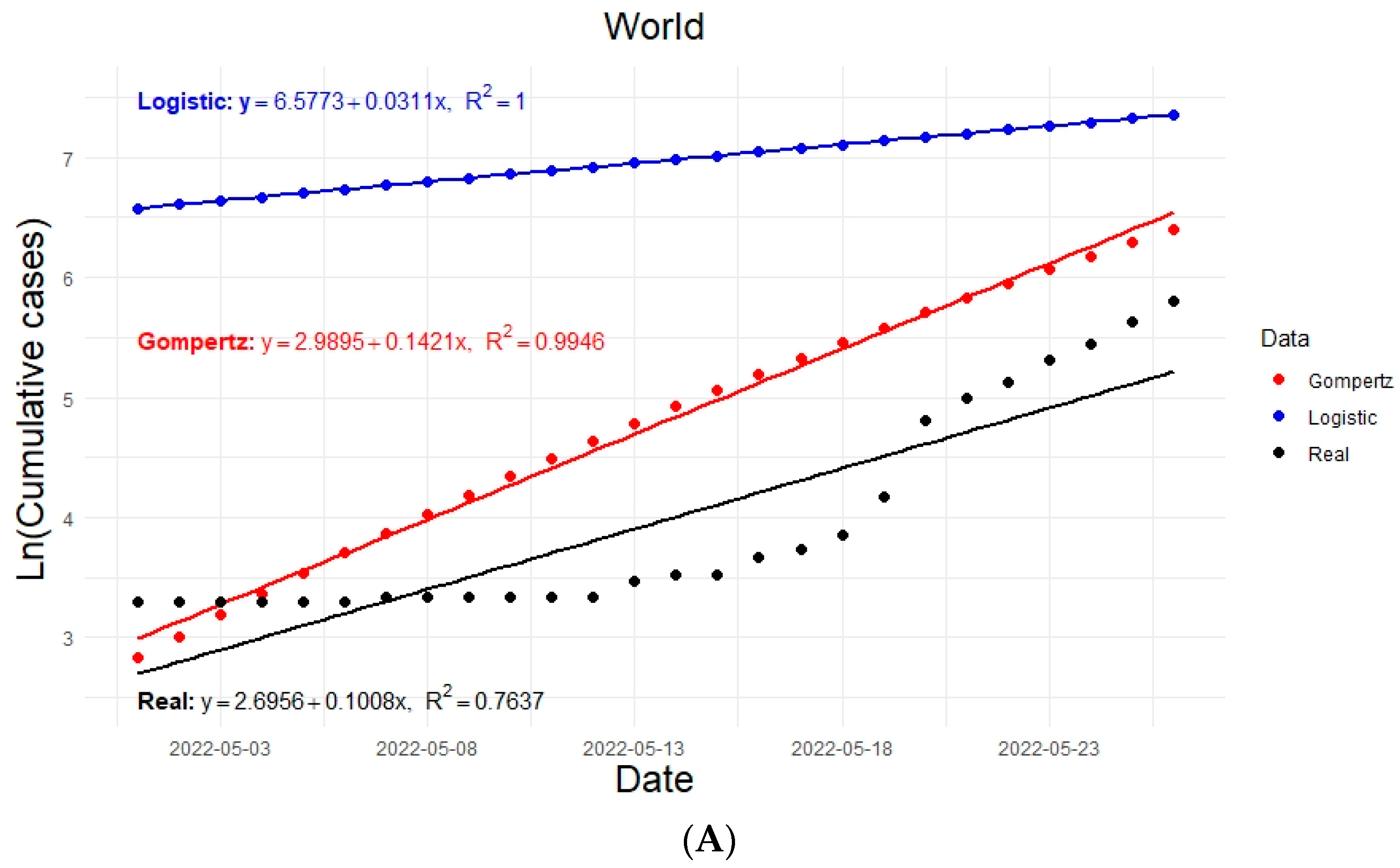
3.3.4. Multiple Linear Regression Analysis for Monkeypox Cases Velocity
3.3.5. Comparison of Modeled Variable against Real Data
3.4. Estimation of the Basic Reproduction Number
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shchelkunov, S.N.; Totmenin, A.V.; Safronov, P.F.; Mikheev, M.V.; Gutorov, V.V.; Ryazankina, O.I.; Petrov, N.A.; Babkin, I.V.; Uvarova, E.A.; Sandakhchiev, L.S.; et al. Analysis of the Monkeypox Virus Genome. Virology 2002, 297, 172–194. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.I.O.; de Oliveira, J.S.; Kroon, E.G.; Trindade, G.d.S.; Drumond, B.P. Here, There, and Everywhere: The Wide Host Range and Geographic Distribution of Zoonotic Orthopoxviruses. Viruses 2020, 13, 43. [Google Scholar] [CrossRef] [PubMed]
- Ferdous, J.; Barek, M.A.; Hossen, M.S.; Bhowmik, K.K.; Islam, M.S. A Review on Monkeypox Virus Outbreak: New Challenge for World. Health Sci. Rep. 2023, 6, e1007. [Google Scholar] [CrossRef] [PubMed]
- Marraha, F.; Al Faker, I.; Chahoub, H.; Benyamna, Y.; Rahmani, N.; Gallouj, S. Monkeypox 2022 Outbreak: How Alarming Is the Situation? Epidemiological and Clinical Review. Clin. Pract. 2023, 13, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Liang, D.; Ling, Q.; Zhang, J.; Li, Z.; Zhang, D.; Xia, P.; Zhu, Z.; Lin, J.; Shi, A.; et al. Insights into Monkeypox Pathophysiology, Global Prevalence, Clinical Manifestation and Treatments. Front. Immunol. 2023, 14, 1132250. [Google Scholar] [CrossRef] [PubMed]
- Cheema, A.Y.; Ogedegbe, O.J.; Munir, M.; Alugba, G.; Ojo, T.K. Monkeypox: A Review of Clinical Features, Diagnosis, and Treatment. Cureus 2022, 14, e26756. [Google Scholar] [CrossRef] [PubMed]
- Bethineedi, L.D.; Kutikuppala, L.V.S.; Kandi, V. Monkeypox Epidemic: A Throwback from Smallpox Eradication. Cureus 2022, 14, e26577. [Google Scholar] [CrossRef]
- Qudus, M.S.; Cui, X.; Tian, M.; Afaq, U.; Sajid, M.; Qureshi, S.; Liu, S.; Ma, J.; Wang, G.; Faraz, M.; et al. The Prospective Outcome of the Monkeypox Outbreak in 2022 and Characterization of Monkeypox Disease Immunobiology. Front. Cell Infect. Microbiol. 2023, 13, 1196699. [Google Scholar] [CrossRef]
- Lansiaux, E.; Jain, N.; Laivacuma, S.; Reinis, A. The Virology of Human Monkeypox Virus (hMPXV): A Brief Overview. Virus Res. 2022, 322, 198932. [Google Scholar] [CrossRef]
- Adekola, H.A.; Adekunle, I.A.; Egberongbe, H.O.; Onitilo, S.A.; Abdullahi, I.N. Mathematical Modeling for Infectious Viral Disease: The COVID-19 Perspective. J. Public Aff. 2020, 20, e2306. [Google Scholar] [CrossRef]
- Josephus, B.O.; Nawir, A.H.; Wijaya, E.; Moniaga, J.V.; Ohyver, M. Predict Mortality in Patients Infected with COVID-19 Virus Based on Observed Characteristics of the Patient Using Logistic Regression. Procedia Comput. Sci. 2021, 179, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Nopour, R.; Shanbehzadeh, M.; Kazemi-Arpanahi, H. Using Logistic Regression to Develop a Diagnostic Model for COVID-19: A Single-Center Study. J. Educ. Health Promot. 2022, 11, 153. [Google Scholar] [CrossRef]
- Marín-Machuca, O.; Chacón, R.D.; Alvarez-Lovera, N.; Pesantes-Grados, P.; Pérez-Timaná, L.; Marín-Sánchez, O. Mathematical Modeling of COVID-19 Cases and Deaths and the Impact of Vaccinations during Three Years of the Pandemic in Peru. Vaccines 2023, 11, 1648. [Google Scholar] [CrossRef]
- Tjørve, K.M.C.; Tjørve, E. The Use of Gompertz Models in Growth Analyses, and New Gompertz-Model Approach: An Addition to the Unified-Richards Family. PLoS ONE 2017, 12, e0178691. [Google Scholar] [CrossRef]
- Vaidya, V.G.; Alexandro, F.J. Evaluation of Some Mathematical Models for Tumor Growth. Int. J. Biomed. Comput. 1982, 13, 19–36. [Google Scholar] [CrossRef]
- Gerlee, P. The Model Muddle: In Search of Tumor Growth Laws. Cancer Res. 2013, 73, 2407–2411. [Google Scholar] [CrossRef] [PubMed]
- Kozusko, F.; Bajzer, Z. Combining Gompertzian Growth and Cell Population Dynamics. Math. Biosci. 2003, 185, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Alharbi, Y.; Alqahtani, A.; Albalawi, O.; Bakouri, M. Epidemiological Modeling of COVID-19 in Saudi Arabia: Spread Projection, Awareness, and Impact of Treatment. Appl. Sci. 2020, 10, 5895. [Google Scholar] [CrossRef]
- Ahmadi, A.; Fadaei, Y.; Shirani, M.; Rahmani, F. Modeling and Forecasting Trend of COVID-19 Epidemic in Iran until May 13, 2020. Med. J. Islam. Repub. Iran 2020, 34, 27. [Google Scholar] [CrossRef]
- Peter, O.J.; Kumar, S.; Kumari, N.; Oguntolu, F.A.; Oshinubi, K.; Musa, R. Transmission Dynamics of Monkeypox Virus: A Mathematical Modelling Approach. Model. Earth Syst. Environ. 2022, 8, 3423–3434. [Google Scholar] [CrossRef]
- Usman, S.; Adamu, I.I. Modeling the Transmission Dynamics of the Monkeypox Virus Infection with Treatment and Vaccination Interventions. J. Appl. Math. Phys. 2017, 5, 2335–2353. [Google Scholar] [CrossRef]
- Ngungu, M.; Addai, E.; Adeniji, A.; Adam, U.M.; Oshinubi, K. Mathematical Epidemiological Modeling, and Analysis of Monkeypox Dynamism with Non-Pharmaceutical Intervention Using Real Data from United Kingdom. Front. Public Health 2023, 11, 1101436. [Google Scholar] [CrossRef]
- Okyere, S.; Ackora-Prah, J. Modeling and Analysis of Monkeypox Disease Using Fractional Derivatives. Results Eng. 2023, 17, 100786. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, A.; Namekawa, Y.; Fukui, T. Universality in COVID-19 Spread in View of the Gompertz Function. Prog. Theor. Exp. Phys. 2020, 2020, 123J01. [Google Scholar] [CrossRef]
- Pelinovsky, E.; Kokoulina, M.; Epifanova, A.; Kurkin, A.; Kurkina, O.; Tang, M.; Macau, E.; Kirillin, M. Gompertz Model in COVID-19 Spreading Simulation. Chaos Solitons Fractals 2022, 154, 111699. [Google Scholar] [CrossRef]
- World Health Organization. 2022–23 Mpox (Monkeypox) Outbreak: Global Trends. Available online: https://worldhealthorg.shinyapps.io/mpx_global/ (accessed on 11 October 2023).
- Wickham, H.; Bryan, J. R Packages: Organize, Test, Document, and Share Your Code, 2nd ed.; O’Reilly: Sebastopol, CA, USA, 2023; ISBN 978-1-09-813494-5. [Google Scholar]
- Kot, M. Elements of Mathematical Ecology; Cambridge University Press: Cambridge, UK, 2001; ISBN 978-0-521-80213-0. [Google Scholar]
- Bacaër, N. Verhulst and the Logistic Equation (1838). In A Short History of Mathematical Population Dynamics; Bacaër, N., Ed.; Springer: London, UK, 2011; pp. 35–39. ISBN 978-0-85729-115-8. [Google Scholar]
- Bronshtein, I.; Semendiaev, K.; Rojas, I.H. Manual de Matemáticas para Ingenieros y Estudiantes; Mir: Moscow, Russia, 1977; ISBN 84-8041-022-1. [Google Scholar]
- Pereira, S.M.C.; Leslie, G. Hypothesis Testing. Aust. Crit. Care 2009, 22, 187–191. [Google Scholar] [CrossRef]
- McKight, P.E.; Najab, J. Kruskal-Wallis Test. In The Corsini Encyclopedia of Psychology; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 2010; ISBN 978-0-470-47921-6. [Google Scholar]
- Elliott, A.C.; Woodward, W.A. Statistical Analysis Quick Reference Guidebook; SAGE Publications, Inc.: Thousand Oaks, CA, USA, 2007; ISBN 978-1-4129-8594-9. [Google Scholar]
- Baguena, J.G. Regresión Lineal Simple; Universitat Oberta de Catalunya: Barcelona, Spain, 2019. [Google Scholar]
- Schober, P.; Boer, C.; Schwarte, L.A. Correlation Coefficients: Appropriate Use and Interpretation. Anesth. Analg. 2018, 126, 1763–1768. [Google Scholar] [CrossRef] [PubMed]
- Vynnycky, E.; White, R.G. An Introduction to Infectious Disease Modelling; Oxford University Press: New York, NY, USA, 2010; ISBN 978-0-19-856576-5. [Google Scholar]
- Guzzetta, G.; Mammone, A.; Ferraro, F.; Caraglia, A.; Rapiti, A.; Marziano, V.; Poletti, P.; Cereda, D.; Vairo, F.; Mattei, G.; et al. Early Estimates of Monkeypox Incubation Period, Generation Time, and Reproduction Number, Italy, May–June 2022. Emerg. Infect. Dis. 2022, 28, 2078–2081. [Google Scholar] [CrossRef] [PubMed]
- Piret, J.; Boivin, G. Pandemics Throughout History. Front. Microbiol. 2020, 11, 631736. [Google Scholar] [CrossRef] [PubMed]
- Sampath, S.; Khedr, A.; Qamar, S.; Tekin, A.; Singh, R.; Green, R.; Kashyap, R. Pandemics Throughout the History. Cureus 2021, 13, e18136. [Google Scholar] [CrossRef]
- Chacón, R.D.; Astolfi-Ferreira, C.S.; Pereira, P.C.; Assayag, M.S., Jr.; Campos-Salazar, A.B.; De la Torre, D.; Sá, L.R.M.d.; Almeida, S.R.Y.d.; Rici, R.E.G.; Ferreira, A.J.P. Outbreaks of Avipoxvirus Clade E in Vaccinated Broiler Breeders with Exacerbated Beak Injuries and Sex Differences in Severity. Viruses 2022, 14, 773. [Google Scholar] [CrossRef] [PubMed]
- Brennan, G.; Stoian, A.M.M.; Yu, H.; Rahman, M.J.; Banerjee, S.; Stroup, J.N.; Park, C.; Tazi, L.; Rothenburg, S. Molecular Mechanisms of Poxvirus Evolution. mBio 2023, 14, e0152622. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Yuan, Y.; Jiang, L.; Liu, Y.; Liu, Y.; Zhang, L. Animal Host Range of Mpox Virus. J. Med. Virol. 2023, 95, e28513. [Google Scholar] [CrossRef]
- Chacón, R.D.; Astolfi-Ferreira, C.S.; De la Torre, D.I.; de Sá, L.R.M.; Piantino Ferreira, A.J. An Atypical Clinicopathological Manifestation of Fowlpox Virus Associated with Reticuloendotheliosis Virus in Commercial Laying Hen Flocks in Brazil. Transbound. Emerg. Dis. 2020, 67, 2923–2935. [Google Scholar] [CrossRef] [PubMed]
- Khamees, A.; Awadi, S.; Al-Shami, K.; Alkhoun, H.A.; Al-Eitan, S.F.; Alsheikh, A.M.; Saeed, A.; Al-Zoubi, R.M.; Zoubi, M.S.A. Human Monkeypox Virus in the Shadow of the COVID-19 Pandemic. J. Infect. Public Health 2023, 16, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Aden, D.; Zaheer, S.; Kumar, R.; Ranga, S. Monkeypox (Mpox) Outbreak during COVID-19 Pandemic-Past and the Future. J. Med. Virol. 2023, 95, e28701. [Google Scholar] [CrossRef]
- Liao, L.-C.; Hsu, C.-Y.; Chen, H.-H.; Lai, C.-C. Estimating the Global Spread of Epidemic Human Monkeypox with Bayesian Directed Acyclic Graphic Model. Vaccines 2023, 11, 468. [Google Scholar] [CrossRef]
- Miura, F.; van Ewijk, C.E.; Backer, J.A.; Xiridou, M.; Franz, E.; Op de Coul, E.; Brandwagt, D.; van Cleef, B.; van Rijckevorsel, G.; Swaan, C.; et al. Estimated Incubation Period for Monkeypox Cases Confirmed in the Netherlands, May 2022. Euro Surveill. 2022, 27, 2200448. [Google Scholar] [CrossRef]
- Du, Z.; Shao, Z.; Bai, Y.; Wang, L.; Herrera-Diestra, J.L.; Fox, S.J.; Ertem, Z.; Lau, E.H.Y.; Cowling, B.J. Reproduction Number of Monkeypox in the Early Stage of the 2022 Multi-Country Outbreak. J. Travel Med. 2022, 29, taac099. [Google Scholar] [CrossRef]
- Zingaropoli, M.A.; Parente, A.; Kertusha, B.; Campagna, R.; Tieghi, T.; Garattini, S.; Marocco, R.; Carraro, A.; Tortellini, E.; Guardiani, M.; et al. Longitudinal Virological and Immunological Profile in a Case of Human Monkeypox Infection. Open Forum Infect. Dis. 2022, 9, ofac569. [Google Scholar] [CrossRef] [PubMed]
- Ophinni, Y.; Frediansyah, A.; Sirinam, S.; Megawati, D.; Stoian, A.M.; Enitan, S.S.; Akele, R.Y.; Sah, R.; Pongpirul, K.; Abdeen, Z.; et al. Monkeypox: Immune Response, Vaccination and Preventive Efforts. Narra J. 2022, 2. [Google Scholar] [CrossRef]
- World Health Organization. History of Smallpox Vaccination. Available online: https://www.who.int/news-room/spotlight/history-of-vaccination/history-of-smallpox-vaccination (accessed on 12 October 2023).
- Mukherjee, A.G.; Wanjari, U.R.; Kannampuzha, S.; Das, S.; Murali, R.; Namachivayam, A.; Renu, K.; Ramanathan, G.; Doss, C.G.P.; Vellingiri, B.; et al. The Pathophysiological and Immunological Background of the Monkeypox Virus Infection: An Update. J. Med. Virol. 2023, 95, e28206. [Google Scholar] [CrossRef] [PubMed]
- Zandi, M.; Shafaati, M.; Hosseini, F. Mechanisms of Immune Evasion of Monkeypox Virus. Front. Microbiol. 2023, 14, 1106247. [Google Scholar] [CrossRef] [PubMed]
- Shafaati, M.; Zandi, M. Human Monkeypox (hMPXV) Re-Emergence: Host Immunity Status and Current Vaccines Landscape. J. Med. Virol. 2023, 95, e28251. [Google Scholar] [CrossRef] [PubMed]
- Cohn, H.; Bloom, N.; Cai, G.Y.; Clark, J.J.; Tarke, A.; Bermúdez-González, M.C.; Altman, D.R.; Lugo, L.A.; Lobo, F.P.; Marquez, S.; et al. Mpox Vaccine and Infection-Driven Human Immune Signatures: An Immunological Analysis of an Observational Study. Lancet Infect. Dis. 2023, 23, 1302–1312. [Google Scholar] [CrossRef]
- Faniran, T.S.; Ali, A.; Al-Hazmi, N.E.; Asamoah, J.K.K.; Nofal, T.A.; Adewole, M.O. New Variant of SARS-CoV-2 Dynamics with Imperfect Vaccine. Complexity 2022, 2022, e1062180. [Google Scholar] [CrossRef]
- Ali, A.; Ullah, S.; Khan, M.A. The Impact of Vaccination on the Modeling of COVID-19 Dynamics: A Fractional Order Model. Nonlinear Dyn. 2022, 110, 3921–3940. [Google Scholar] [CrossRef]
- Xu, C.; Wang, Y.; Cheng, K.; Yang, X.; Wang, X.; Guo, S.; Liu, M.; Liu, X. A Mathematical Model to Study the Potential Hepatitis B Virus Infections and Effects of Vaccination Strategies in China. Vaccines 2023, 11, 1530. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Parameters | World | Europe | North America | South America |
|---|---|---|---|---|---|
| Logistic | Coefficient of Determination R2 | 0.8885 | 0.8720 | 0.9023 | 0.8774 |
| Logistic | Critical time tc (days) | 152 | 108 | 117 | 144 |
| Logistic | Date on tc | 30 September 2022 | 23 August 2022 | 28 September 2022 | 25 October 2022 |
| Logistic | Nmax on tc | 43,648 | 13,926 | 18,515 | 11,160 |
| Logistic | Highest contagion rate (people/day) | 690 | 230 | 278 | 206 |
| Gompertz | Coefficient of Determination R2 | 0.9952 | 0.9881 | 0.9900 | 0.9988 |
| Gompertz | Critical time tc (days) | 100 | 62 | 75 | 94 |
| Gompertz | Date on tc | 9 August 2022 | 8 July 2022 | 17 August 2022 | 5 September 2022 |
| Gompertz | Nmax on tc | 32,656 | 9463 | 13,699 | 8264 |
| Gompertz | Highest contagion rate (people/day) | 696 | 268 | 308 | 202 |
| Model | Geographic Region | p-Value | Hypothesis Testing | Interpretation |
|---|---|---|---|---|
| Logistic | World | 6.39 × 10−34 | Reject H0 | Non-Normal Distribution |
| Logistic | Europe | 4.11 × 10−60 | Reject H0 | Non-Normal Distribution |
| Logistic | North America | 1.08 × 10−32 | Reject H0 | Non-Normal Distribution |
| Logistic | South America | 1.68 × 10−30 | Reject H0 | Non-Normal Distribution |
| Gompertz | World | 1.94 × 10−39 | Reject H0 | Non-Normal Distribution |
| Gompertz | Europe | 3.77 × 10−98 | Reject H0 | Non-Normal Distribution |
| Gompertz | North America | 5.37 × 10−39 | Reject H0 | Non-Normal Distribution |
| Gompertz | South America | 2.87 × 10−36 | Reject H0 | Non-Normal Distribution |
| Model | Geographic Region | World | Europe | North America |
|---|---|---|---|---|
| Logistic | Europe | 3.71 × 10−30 | - | - |
| Logistic | North America | 3.92 × 10−15 | 8.65 × 10−6 | - |
| Logistic | South America | 6.23 × 10−32 | 0.894 | 1.73 × 10−6 |
| Gompertz | Europe | 3.48 × 10−32 | - | - |
| Gompertz | North America | 1.71 × 10−15 | 2.93 × 10−6 | - |
| Gompertz | South America | 5.17 × 10−31 | 0.546 | 5.15 × 10−6 |
| Model | Geographic Region | p-Value | Hypothesis Testing | Interpretation |
|---|---|---|---|---|
| Logistic | World | <2.2 × 10−16 | Reject H0 | Significant effect |
| Logistic | Europe | 0.001295 | Reject H0 | Significant effect |
| Logistic | North America | 0.000108 | Reject H0 | Significant effect |
| Logistic | South America | 9.74 × 10−7 | Reject H0 | Significant effect |
| Gompertz | World | <2.2 × 10−16 | Reject H0 | Significant effect |
| Gompertz | Europe | 0.000314 | Reject H0 | Significant effect |
| Gompertz | North America | <2.2 × 10−16 | Reject H0 | Significant effect |
| Gompertz | South America | <2.2 × 10−16 | Reject H0 | Significant effect |
| Geographic Region | –Logistic | –Gompertz | A.I.C.–Logistic | A.I.C.–Gompertz |
|---|---|---|---|---|
| World | 0.8885 | 0.9952 | 6836.8260 | 5669.1384 |
| Europe | 0.8720 | 0.9881 | 5783.6794 | 5066.7892 |
| North America | 0.9023 | 0.9900 | 6087.4334 | 5336.7998 |
| South America | 0.8774 | 0.9988 | 5869.6060 | 4229.3746 |
| Geographic Region | Graphical | Graphical | Analytical | Analytical |
|---|---|---|---|---|
| World | 0.0311 | 0.1421 | 0.0315 | 0.2458 |
| Europe | 0.0349 | 0.1179 | 0.0362 | 0.2893 |
| North America | 0.0287 | 0.0931 | 0.0300 | 0.2377 |
| South America | 0.0364 | 0.1820 | 0.0367 | 0.2454 |
| Geographic Region | Graphical Logistic | Graphical Gompertz | Analytical Logistic | Analytical Gompertz |
|---|---|---|---|---|
| World | 1.78178 | 6.36625 | 1.79327 | 13.18179 |
| Europe | 1.89239 | 5.12781 | 1.93098 | 16.76913 |
| North America | 1.71361 | 3.99690 | 1.75038 | 12.56134 |
| South America | 1.93695 | 8.69906 | 1.94593 | 13.15080 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marín-Sánchez, O.; Pesantes-Grados, P.; Pérez-Timaná, L.; Marín-Machuca, O.; Sánchez-Llatas, C.J.; Chacón, R.D. Comparative Epidemiological Assessment of Monkeypox Infections on a Global and Continental Scale Using Logistic and Gompertz Mathematical Models. Vaccines 2023, 11, 1765. https://doi.org/10.3390/vaccines11121765
Marín-Sánchez O, Pesantes-Grados P, Pérez-Timaná L, Marín-Machuca O, Sánchez-Llatas CJ, Chacón RD. Comparative Epidemiological Assessment of Monkeypox Infections on a Global and Continental Scale Using Logistic and Gompertz Mathematical Models. Vaccines. 2023; 11(12):1765. https://doi.org/10.3390/vaccines11121765
Chicago/Turabian StyleMarín-Sánchez, Obert, Pedro Pesantes-Grados, Luis Pérez-Timaná, Olegario Marín-Machuca, Christian J. Sánchez-Llatas, and Ruy D. Chacón. 2023. "Comparative Epidemiological Assessment of Monkeypox Infections on a Global and Continental Scale Using Logistic and Gompertz Mathematical Models" Vaccines 11, no. 12: 1765. https://doi.org/10.3390/vaccines11121765