
Field Efficacy of a Feed-Based Inactivated Vaccine against Vibriosis in Cage-Cultured Asian Seabass, Lates calcarifer, in Malaysia
 , ,
, ,  ,
,  , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Growth Condition
2.2. Feed-Based Vaccine Preparation
2.3. Ethics Approval
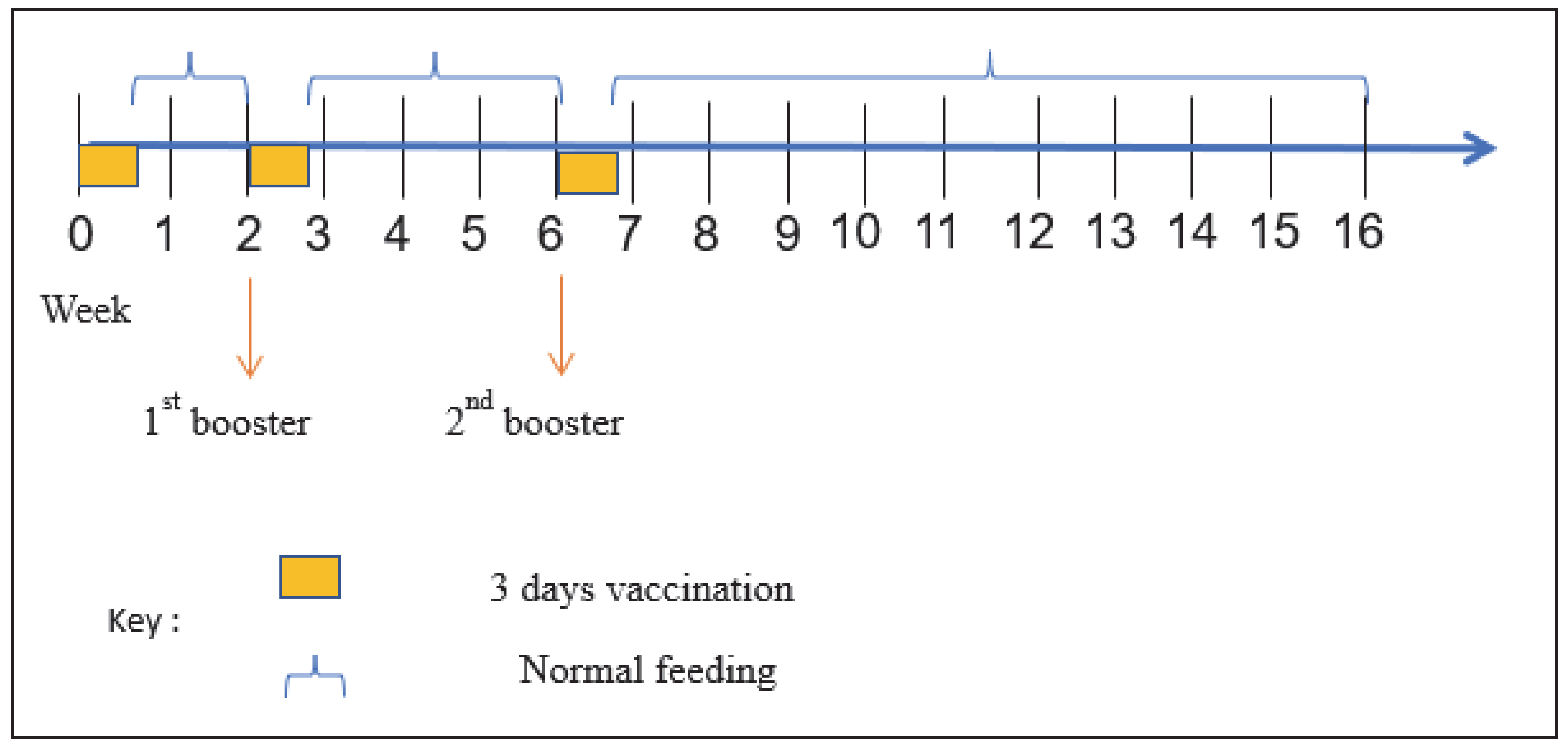
2.4. Study Location and Study Design
2.5. Growth Performance
2.6. Sample Processing
2.6.1. Isolation and Identification of Bacteria
2.6.2. Humoral Non-Specific Immune Parameters in Mucus and Serum Samples
2.6.3. Detection of V. harveyi-Specific IgM in Serum and Mucus with ELISA
2.7. Statistical Analysis
3. Results
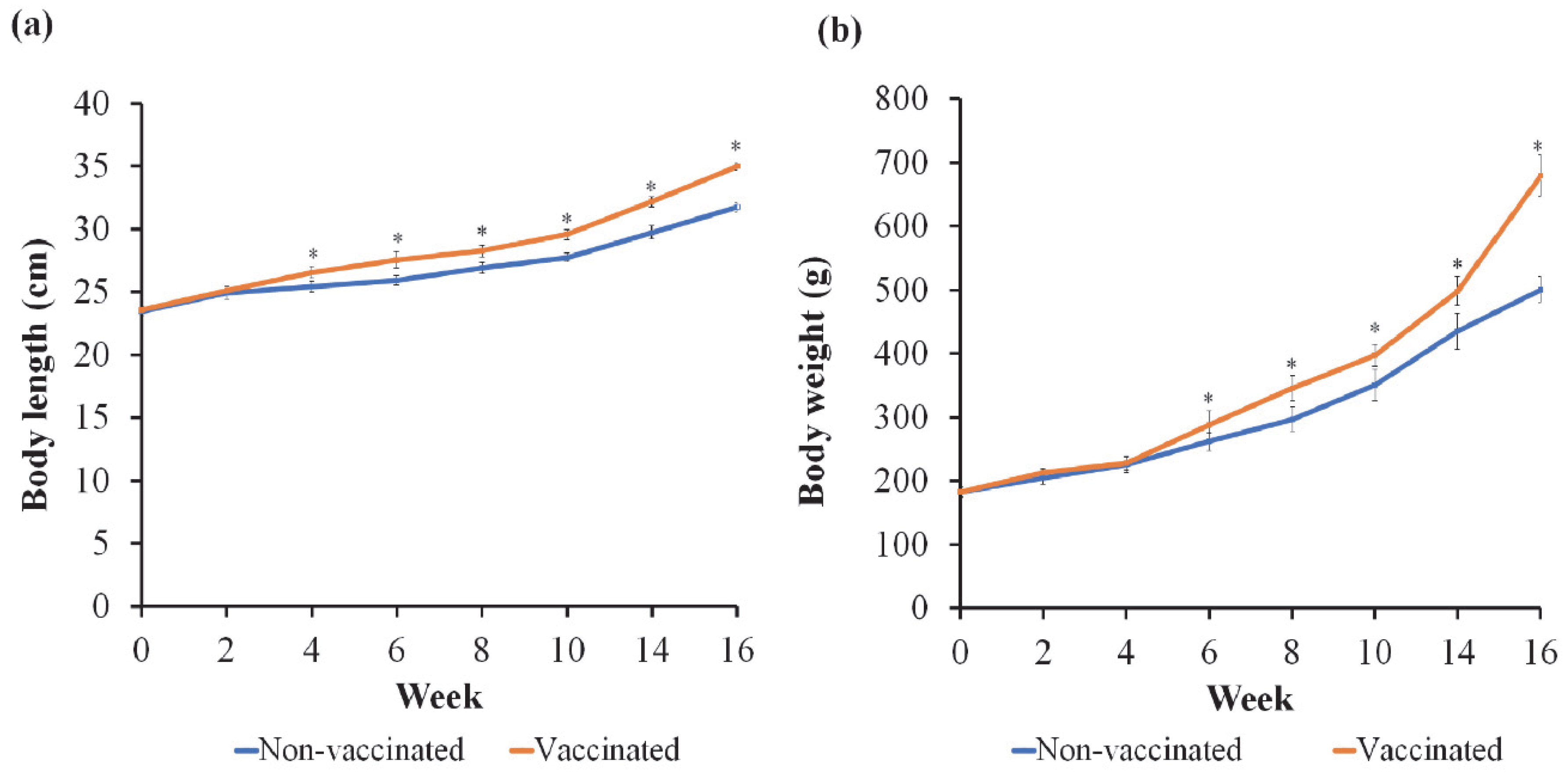
3.1. Growth Performance and Rate of Survival
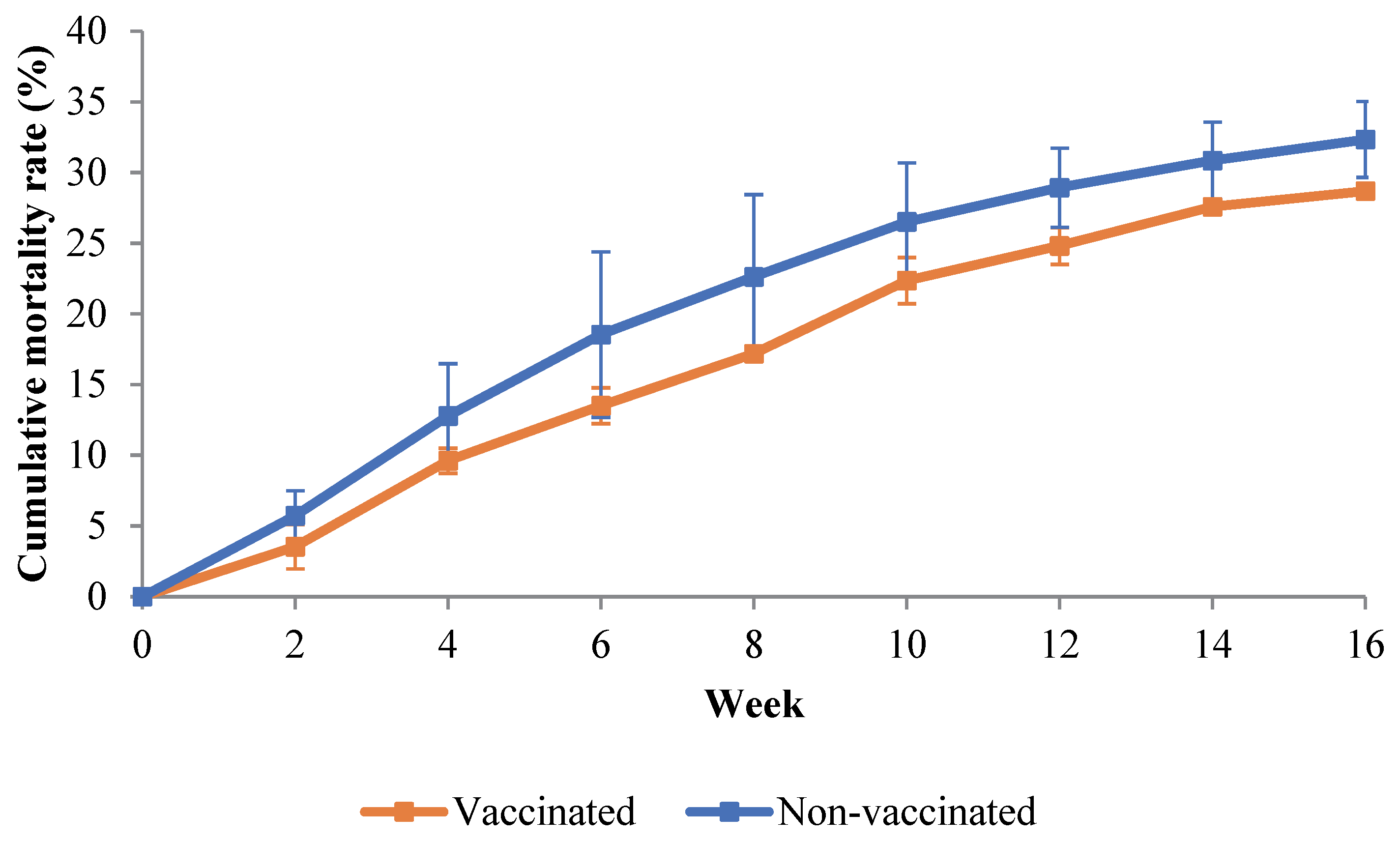
3.2. Cumulative Mortality Rate
3.3. Isolation and Identification of Bacteria
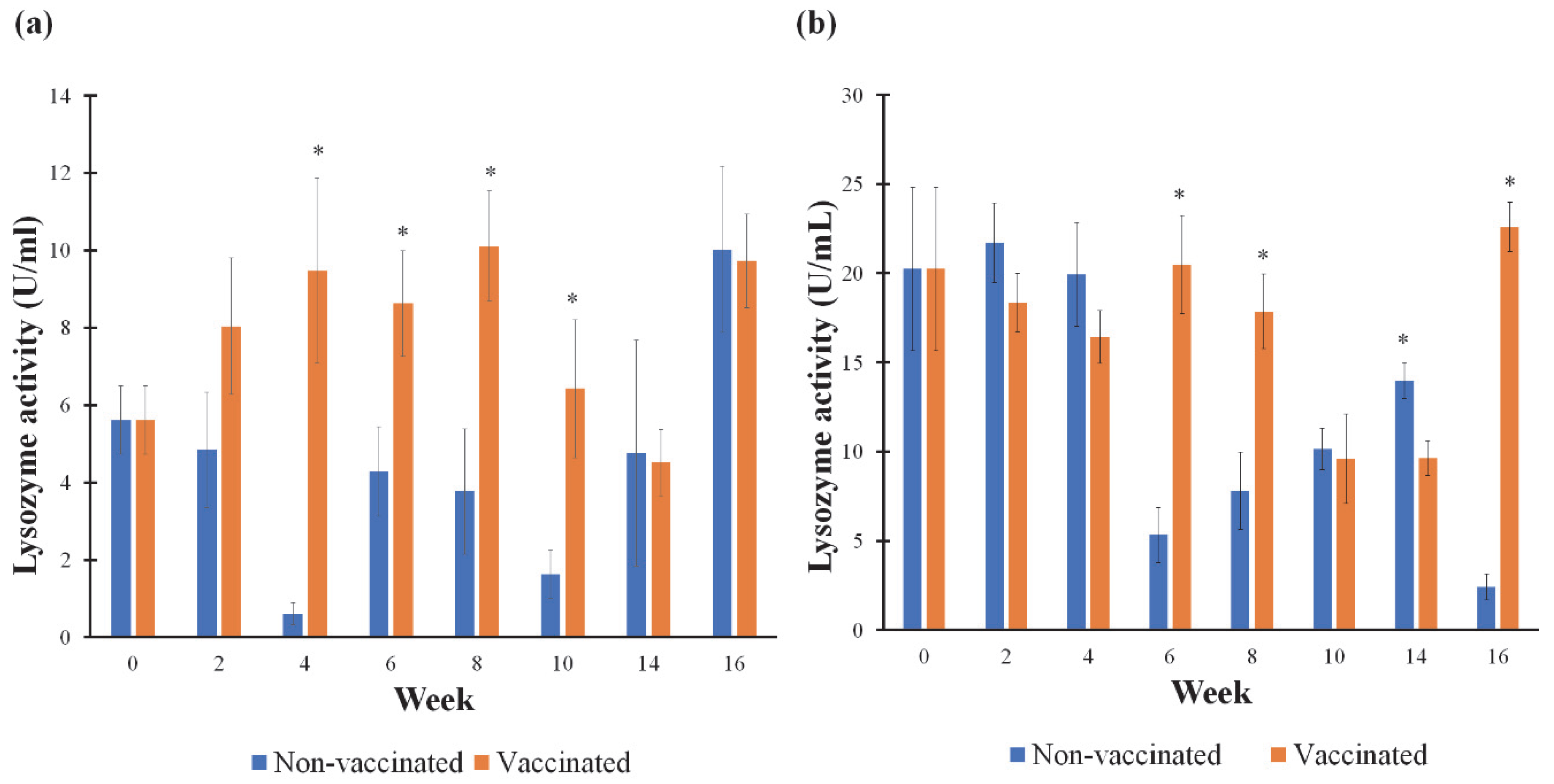
3.4. Humoral Non-Specific Immune Parameters in Mucus and Serum Samples
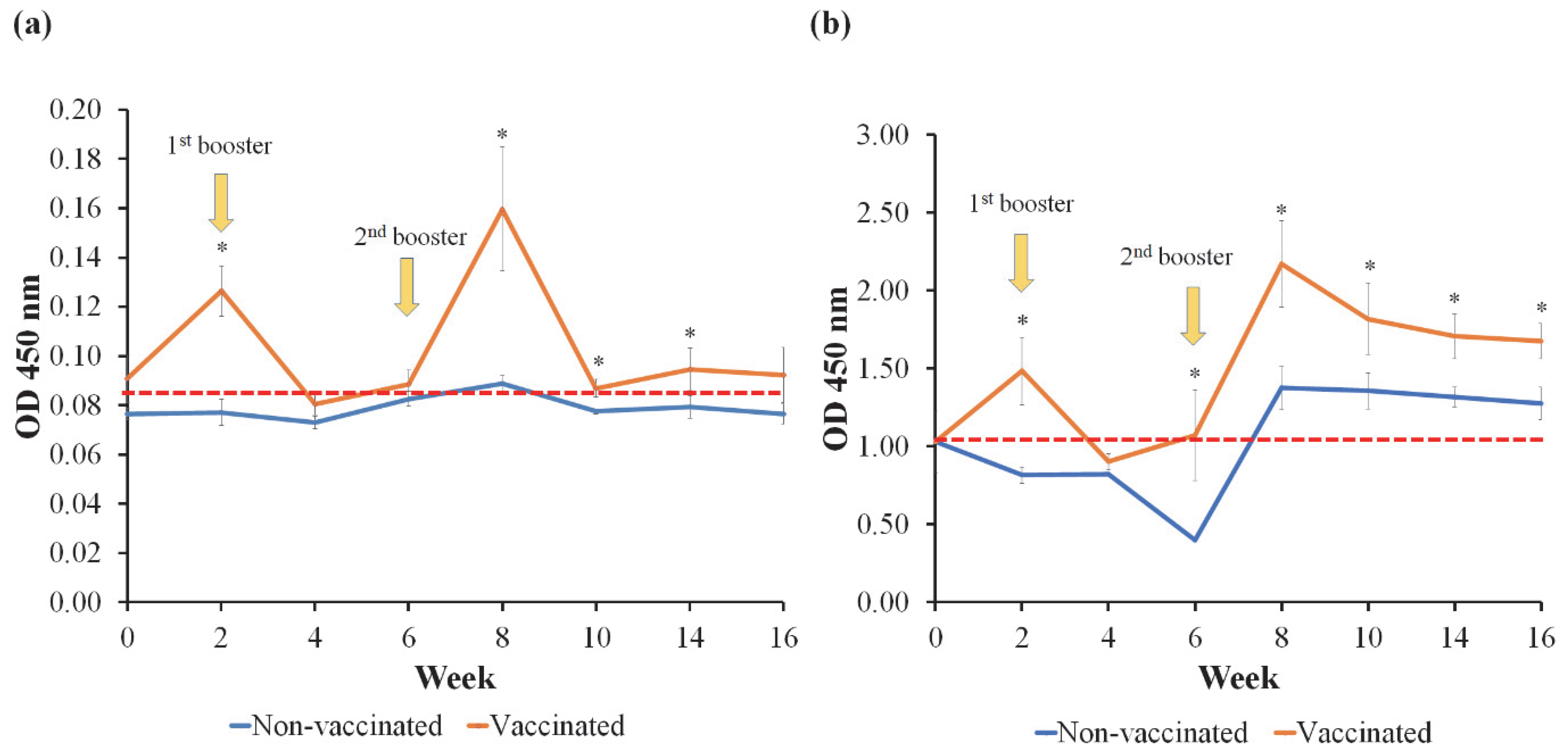
3.5. Detection of V. harveyi-Specific IgM in Serum and Mucus with ELISA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heenatigala, P.P.M.; Sun, Z.; Yang, J.; Zhao, X.; Hou, H. Expression of LamB Vaccine Antigen in Wolffia globosa (Duck Weed) Against Fish Vibriosis. Front. Immunol. 2020, 11, 1857. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Liu, D.; Han, S.; Zhou, Y.; Wang, K.Y.; Huang, X.L.; Chen, D.F.; Peng, X.; Lai, W.M. Outbreaks of vibriosis associated with Vibrio mimicus in freshwater catfish in China. Aquaculture 2014, 433, 82–84. [Google Scholar] [CrossRef]
- Atujona, D.; Huang, Y.; Wang, Z.; Jian, J.; Cai, S. Vibrio harveyi (VirB11) recombinant vaccine development against vibriosis in orange-spotted grouper (Epinephelus coioides). Aquac. Res. 2019, 50, 2628–2634. [Google Scholar] [CrossRef]
- Zhang, X.-H.; He, X.; Austin, B. Vibrio harveyi: A serious pathogen of fish and invertebrates in mariculture. Mar. Life Sci. Technol. 2020, 2, 231–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurliyana, M.; Amal, M.N.A.; Zamri-Saad, M.; Ina-Salwany, M.Y. Possible transmission routes of Vibrio spp. in tropical cage-cultured marine fishes. Lett. Appl. Microbiol. 2019, 68, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Istiqomah, I.; Sukardi; Murwantoko; Isnansetyo, A. Review Vibriosis Management in Indonesian Marine Fish Farming. E3S Web Conf. 2020, 147, 01001. [Google Scholar] [CrossRef] [Green Version]
- Nurhafizah, W.W.I.; Lee, K.L.; Laith A., A.R.; Nadirah, M.; Danish-Daniel, M.; Zainathan, S.C.; Najiah, M. Virulence properties and pathogenicity of multidrug-resistant Vibrio harveyi associated with luminescent vibriosis in Pacific white shrimp, Penaeus vannamei. J. Invertebr. Pathol. 2021, 186, 107594. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Yilmaz, E.; Dawood, M.A.O.; Ringø, E.; Ahmadifar, E.; Yilmaz, S. Shrimp vibriosis and possible control measures using probiotics, postbiotics, prebiotics, and synbiotics: A review. Aquaculture 2022, 551, 737951. [Google Scholar] [CrossRef]
- Chiew, I.K.M.; Salter, A.M.; Lim, Y.S. The significance of major viral and bacterial diseases in Malaysian aquaculture industry. Pertanika J. Trop. Agric. Sci. 2019, 42, 1023–1047. [Google Scholar]
- Mohamad, N.; Amal, M.N.A.; Yasin, I.S.M.; Saad, M.Z.; Nasruddin, N.S.; Al-saari, N.; Mino, S.; Sawabe, T. Vibriosis in cultured marine fishes: A review. Aquaculture 2019, 512, 734289. [Google Scholar] [CrossRef]
- Nor, N.M.; Yazid, S.H.M.; Daud, H.M.; Azmai, M.N.A.; Mohamad, N. Costs of management practices of Asian seabass (Lates calcarifer Bloch, 1790) cage culture in Malaysia using stochastic model that includes uncertainty in mortality. Aquaculture 2019, 510, 347–352. [Google Scholar] [CrossRef]
- Liu, X.; Jiao, C.; Ma, Y.; Wang, Q.; Zhang, Y. A live attenuated Vibrio anguillarum vaccine induces efficient immunoprotection in Tiger puffer (Takifugu rubripes). Vaccine 2018, 36, 1460–1466. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Beck, B.R.; Hwang, S.H.; Song, S.K. Feeding olive flounder (Paralichthys olivaceus) with Lactococcus lactis BFE920 expressing the fusion antigen of Vibrio OmpK and FlaB provides protection against multiple Vibrio pathogens: A universal vaccine effect. Fish Shellfish Immunol. 2021, 114, 253–262. [Google Scholar] [CrossRef]
- Su, C.; Chen, L. Virulence, resistance, and genetic diversity of Vibrio parahaemolyticus recovered from commonly consumed aquatic products in Shanghai, China. Mar. Pollut. Bull. 2020, 160, 111554. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.-X.; Ma, Y.-M.; Li, H.; Peng, X.-X. N-acetylglucosamine enhances survival ability of tilapias infected by Streptococcus iniae. Fish Shellfish Immunol. 2014, 40, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ye, Z.; Wen, L.; Chen, R.; Tian, L.; Zhao, F.; Pan, J. Identification of a novel vaccine candidate by immunogenic screening of Vibrio parahaemolyticus outer membrane proteins. Vaccine 2014, 32, 6115–6121. [Google Scholar] [CrossRef]
- Pang, H.; Chen, L.; Hoare, R.; Huang, Y.; Wu, Z.; Jian, J. Identification of DLD, by immunoproteomic analysis and evaluation as a potential vaccine antigen against three Vibrio species in Epinephelus coioides. Vaccine 2016, 34, 1225–1231. [Google Scholar] [CrossRef]
- Annas, S.; Zamri-Saad, M. Intranasal vaccination strategy to control the COVID-19 pandemic from a veterinary medicine perspective. Animals 2021, 11, 1876. [Google Scholar] [CrossRef]
- Ismail, M.S.; Siti-Zahrah, A.; Syafiq, M.R.M.; Amal, M.N.A.; Firdaus-Nawi, M.; Zamri-Saad, M. Feed-based vaccination regime against streptococcosis in red tilapia, Oreochromis niloticus × Oreochromis mossambicus. BMC Vet. Res. 2016, 12, 194. [Google Scholar] [CrossRef] [Green Version]
- Hoare, R.; Jung, S.J.; Ngo, T.P.H.; Bartie, K.; Bailey, J.; Thompson, K.D.; Adams, A. Efficacy and safety of a non-mineral oil adjuvanted injectable vaccine for the protection of Atlantic salmon (Salmo salar L.) against Flavobacterium psychrophilum. Fish Shellfish Immunol. 2019, 85, 44–51. [Google Scholar] [CrossRef]
- Mohamad, A.; Zamri-Saqaaad, M.; Amal, M.N.A.; Al-Saari, N.; Monir, M.S.; Chin, Y.K.; Md Yasin, I.S. Vaccine efficacy of a newly developed feed-based whole-cell polyvalent vaccine against vibriosis, streptococcosis and motile aeromonad septicemia in asian seabass, Lates calcarifer. Vaccines 2021, 9, 368. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, A.; Mursidi, F.-A.; Zamri-Saad, M.; Amal, M.N.A.; Annas, S.; Monir, M.S.; Loqman, M.; Hairudin, F.; Al-saari, N.; Ina-Salwany, M.Y. Laboratory and Field Assessments of Oral Vibrio Vaccine Indicate the Potential for Protection against Vibriosis in Cultured Marine Fishes. Animals 2022, 12, 133. [Google Scholar] [CrossRef] [PubMed]
- Mursidi, F. Antigenic Analysis of outer Membrane Protein of Vibrio Species and Development of Versatile Recombinant vhDnaJ Vaccine against Vibriosis. Master Thesis, Universiti Putra Malaysia, Serdang, Malaysia, 2018. [Google Scholar]
- Lin, H.L.; Shiu, Y.L.; Chiu, C.S.; Huang, S.L.; Liu, C.H. Screening probiotic candidates for a mixture of probiotics to enhance the growth performance, immunity, and disease resistance of Asian seabass, Lates calcarifer (Bloch), against Aeromonas hydrophila. Fish Shellfish Immunol. 2017, 60, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Byadgi, O.; Uyen, N.H.N.; Chou, R.L.; Guo, J.J.; Lee, Y.H.; Lee, J.W.; Cheng, T.C. Immunogenicity of inactivated formalin-killed Photobacterium damselae subsp. piscicida combined with Toll-like receptor 9 agonist in Cobia Rachycentron canadum. Aquaculture 2018, 492, 369–378. [Google Scholar] [CrossRef]
- Firdaus-Nawi, M.; Yusoff, S.M.; Yusof, H.; Abdullah, S.Z.; Zamri-Saad, M. Efficacy of feed-based adjuvant vaccine against Streptococcus agalactiae in Oreochromis spp. in Malaysia. Aquac. Res. 2013, 45, 87–96. [Google Scholar] [CrossRef]
- Plaza, N.; Castillo, D.; Pérez-Reytor, D.; Higuera, G.; García, K.; Bastías, R. Bacteriophages in the control of pathogenic vibrios. Electron. J. Biotechnol. 2018, 31, 24–33. [Google Scholar] [CrossRef]
- Bin Park, S.; Nho, S.W.; Bin Jang, H.; Cha, I.S.; Kim, M.S.; Lee, W.-J.; Jung, T.S. Development of three-valent vaccine against streptococcal infections in olive flounder, Paralichthys olivaceus. Aquaculture 2016, 461, 25–31. [Google Scholar] [CrossRef]
- Lun, J.; Xia, C.; Yuan, C.; Zhang, Y.; Zhong, M.; Huang, T.; Hu, Z. The outer membrane protein, LamB (maltoporin), is a versatile vaccine candidate among the Vibrio species. Vaccine 2014, 32, 809–815. [Google Scholar] [CrossRef]
- Dong, H.T.; Nguyen, V.V.; Phiwsaiya, K.; Gangnonngiw, W.; Withyachumnarnkul, B.; Rodkhum, C.; Senapin, S. Concurrent infections of Flavobacterium columnare and Edwardsiella ictaluri in striped catfish, Pangasianodon hypophthalmus in Thailand. Aquaculture 2015, 448, 142–150. [Google Scholar] [CrossRef]
- Shoemaker, C.A.; LaFrentz, B.R.; Klesius, P.H. Bivalent vaccination of sex reversed hybrid tilapia against Streptococcus iniae and Vibrio vulnificus. Aquaculture 2012, 354–355, 45–49. [Google Scholar] [CrossRef]
- Peng, B.; Lin, X.-P.; Wang, S.-N.; Yang, M.-J.; Peng, X.-X.; Li, H. Polyvalent protective immunogens identified from outer membrane proteins of Vibrio parahaemolyticus and their induced innate immune response. Fish Shellfish Immunol. 2018, 72, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Wang, S.; Ma, J.; Liu, Q. A review: Progress in the development of fish Vibrio spp. vaccines. Immunol. Lett. 2020, 226, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chu, X.; Li, D.; Zeng, Z.H.; Peng, X.X. Construction and immune protection evaluation of recombinant polyvalent OmpAs derived from genetically divergent ompA by DNA shuffling. Fish Shellfish Immunol. 2016, 49, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Mohd-Aris, A.; Saad, M.Z.; Daud, H.M.; Yusof, M.T.; Ina-Salwany, M.Y. Vibrio harveyi protease deletion mutant as a live attenuated vaccine candidate against vibriosis and transcriptome profiling following vaccination for Epinephelus fuscoguttatus. Aquac. Int. 2019, 27, 125–140. [Google Scholar] [CrossRef]
- Chin, Y.K.; Ina-Salwany, M.Y.; Zamri-Saad, M.; Amal, M.N.A.; Mohamad, A.; Lee, J.Y.; Annas, S.; Al-Saari, N. Effects of skin abrasion in immersion challenge with Vibrio harveyi in Asian seabass Lates calcarifer fingerlings. Dis. Aquat. Org. 2020, 137, 167–173. [Google Scholar] [CrossRef]
- Gravningen, K.; Sakai, M.; Mishiba, T.; Fujimoto, T. The efficacy and safety of an oil-based vaccine against Photobacterium damsela subsp. piscicida in yellowtail (Seriola quinqueradiata): A field study. Fish Shellfish Immunol. 2008, 24, 523–529. [Google Scholar] [CrossRef]
- Lan, N.G.T.; Salin, K.R.; Longyant, S.; Senapin, S.; Dong, H.T. Systemic and mucosal antibody response of freshwater cultured Asian seabass (Lates calcarifer) to monovalent and bivalent vaccines against Streptococcus agalactiae and Streptococcus iniae. Fish Shellfish Immunol. 2021, 108, 7–13. [Google Scholar] [CrossRef]
- Tobar, I.; Arancibia, S.; Torres, C.; Vera, V.; Soto, P.; Carrasco, C.; Alvarado, M.; Neira, E.; Arcos, S.; Tobar, J.A. Successive oral immunizations against Piscirickettsia salmonis and infectious salmon anemia virus are required to maintain a long-term protection in farmed salmonids. Front. Immunol. 2015, 6, 244. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.Y.; Chen, Y.C.; Wang, P.C.; Tsai, M.A.; Yeh, S.C.; Liang, H.J.; Chen, S.C. Efficacy of a formalin-inactivated vaccine against Streptococcus iniae infection in the farmed grouper Epinephelus coioides by intraperitoneal immunization. Vaccine 2014, 32, 7014–7020. [Google Scholar] [CrossRef]
- Hayat, M.; Yusoff, M.S.M.; Samad, M.J.; Razak, I.S.A.; Yasin, I.S.M.; Thompson, K.D.; Hasni, K. Efficacy of feed-based formalin-killed vaccine of Streptococcus iniae stimulates the gut-associated lymphoid tissues and immune response of red hybrid tilapia. Vaccines 2021, 9, 51. [Google Scholar] [CrossRef]
- Somamoto, T.; Nakanishi, T. Mucosal delivery of fish vaccines: Local and systemic immunity following mucosal immunisations. Fish Shellfish Immunol. 2020, 99, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Nur-Nazifah, M.; Sabri, M.Y.; Siti-Zahrah, A. Development and efficacy of feed-based recombinant vaccine encoding the cell wall surface anchor family protein of Streptococcus agalactiae against streptococcosis in Oreochromis sp. Fish Shellfish Immunol. 2014, 37, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Dotta, G.; de Andrade, J.I.A.; Gonçalves, E.L.T.; Brum, A.; Mattos, J.J.; Maraschin, M.; Martins, M.L. Leukocyte phagocytosis and lysozyme activity in Nile tilapia fed supplemented diet with natural extracts of propolis and Aloe barbadensis. Fish Shellfish Immunol. 2014, 39, 280–284. [Google Scholar] [CrossRef]
- Nor, N.A.; Zamri-Saad, M.; Yasin, I.S.M.; Salleh, A.; Mustaffa-Kamal, F.; Matori, M.F.; Azmai, M.N.A. Efficacy of whole cell inactivated Vibrio harveyi vaccine against vibriosis in a marine red hybrid tilapia (Oreochromis niloticus × O. mossambicus) model. Vaccines 2020, 8, 734. [Google Scholar] [CrossRef] [PubMed]
- Sutthi, N.; Panase, A.; Chitmanat, C.; Sookying, S.; Ratworawong, K.; Panase, P. Effects of dietary leaf ethanolic extract of Apium graveolens L. on growth performance, serum biochemical indices, bacterial resistance and lysozyme activity in Labeo chrysophekadion (Bleeker, 1849). Aquac. Rep. 2020, 18, 100551. [Google Scholar] [CrossRef]
- Tian, J.; Yu, J. Corrigendum to “poly (lactic-co-glycolic acid) nanoparticles as candidate DNA vaccine carrier for oral immunization of Japanese flounder (Paralichthys olivaceus) against lymphocystis disease virus” Fish Shellfish Immun. 30 (2011) 109-117. Fish Shellfish Immunol. 2011, 31, 364. [Google Scholar] [CrossRef]
- Kole, S.; Kumari, R.; Anand, D.; Kumar, S.; Sharma, R.; Tripathi, G.; Makesh, M.; Rajendran, K.V.; Bedekar, M.K. Nanoconjugation of bicistronic DNA vaccine against Edwardsiella tarda using chitosan nanoparticles: Evaluation of its protective efficacy and immune modulatory effects in Labeo rohita vaccinated by different delivery routes. Vaccine 2018, 36, 2155–2165. [Google Scholar] [CrossRef]
- Monir, M.S.; Yusoff, S.B.M.; Zulperi, Z.B.M.; Hassim, H.B.A.; Mohamad, A.; Ngoo, M.S.B.M.H.; Ina-Salwany, M.Y. Haemato-immunological responses and effectiveness of feed-based bivalent vaccine against Streptococcus iniae and Aeromonas hydrophila infections in hybrid red tilapia (Oreochromis mossambicus × O. niloticus). BMC Vet. Res. 2020, 16, 226. [Google Scholar] [CrossRef]
- Lazado, C.C.; Caipang, C.M.A. Mucosal immunity and probiotics in fish. Fish Shellfish Immunol. 2014, 39, 78–89. [Google Scholar] [CrossRef]
- Zhang, J.; Dong, C.; Feng, J.; Li, J.; Li, S.; Feng, J.; Duan, X.; Sun, G.; Xu, P.; Li, X. Effects of dietary supplementation of three strains of Lactococcus lactis on HIFs genes family expression of the common carp following Aeromonas hydrophila infection. Fish Shellfish Immunol. 2019, 92, 590–599. [Google Scholar] [CrossRef]
- Ma, Y.; Hao, L.; Liang, Z.; Ma, J.; Ke, H.; Kang, H.; Yang, H.; Wu, J.; Feng, G.; Liu, Z. Characterization of novel antigenic vaccine candidates for nile tilapia (Oreochromis niloticus) against Streptococcus agalactiae infection. Fish Shellfish Immunol. 2020, 105, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, F.; Guo, H.; Zhu, Y.; Yuan, J.; Yang, G.; An, L. Molecular characterization of hepcidin gene in common carp (Cyprinus carpio L.) and its expression pattern responding to bacterial challenge. Fish Shellfish Immunol. 2013, 35, 1030–1038. [Google Scholar] [CrossRef] [PubMed]
- Jun, L.; Woo, N.Y.S. Pathogenicity of vibrios in fish: An overview. J. Ocean Univ. China 2003, 2, 117–128. [Google Scholar] [CrossRef]
- Frans, I.; Michiels, C.W.; Bossier, P.; Willems, K.A.; Lievens, B.; Rediers, H. Vibrio anguillarum as a fish pathogen: Virulence factors, diagnosis and prevention. J. Fish Dis. 2011, 34, 643–661. [Google Scholar] [CrossRef] [PubMed]
- Amar, E.C.; Faisan, J.P.; Gapasin, R.S.J. Field efficacy evaluation of a formalin-inactivated white spot syndrome virus (WSSV) vaccine for the preventive management of WSSV infection in shrimp grow-out ponds. Aquaculture 2021, 531, 735907. [Google Scholar] [CrossRef]
- Kim, D.; Beck, B.R.; Heo, S.B.; Kim, J.; Kim, H.D.; Lee, S.M.; Kim, Y.; Oh, S.Y.; Lee, K.; Do, H.K.; et al. Lactococcus lactis BFE920 activates the innate immune system of olive flounder (Paralichthys olivaceus), resulting in protection against Streptococcus iniae infection and enhancing feed efficiency and weight gain in large-scale field studies. Fish Shellfish Immunol. 2013, 35, 1585–1590. [Google Scholar] [CrossRef] [PubMed]
- Beck, B.R.; Lee, S.H.; Kim, D.; Park, J.H.; Lee, H.K.; Kwon, S.S.; Lee, K.H.; Lee, J., II; Song, S.K. A Lactococcus lactis BFE920 feed vaccine expressing a fusion protein composed of the OmpA and FlgD antigens from Edwardsiella tarda was significantly better at protecting olive flounder (Paralichthys olivaceus) from edwardsiellosis than single antigen vac. Fish Shellfish Immunol. 2017, 68, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M.S.; Syafiq, M.R.; Siti-Zahrah, A.; Fahmi, S.; Shahidan, H.; Hanan, Y.; Amal, M.N.A.; Zamri Saad, M. The effect of feed-based vaccination on tilapia farm endemic for streptococcosis. Fish Shellfish Immunol. 2017, 60, 21–24. [Google Scholar] [CrossRef]
- Chatakondi, N.; Peterson, B.C.; Greenway, T.E.; Byars, T.S.; Wise, D.J. Efficacy of a Live-attenuated Edwardsiella ictaluri Oral Vaccine in Channel and Hybrid Catfish. J. World Aquac. Soc. 2018, 49, 686–691. [Google Scholar] [CrossRef]
- Hazreen-Nita, M.; Azila, A.; Mukai, Y.; Firdaus-Nawi, M.; Nur-Nazifah, M. A review of betanodavirus vaccination as preventive strategy to viral nervous necrosis (VNN) disease in grouper. Aquac. Int. 2019, 27, 1565–1577. [Google Scholar] [CrossRef]
- Brudeseth, B.E.; Wiulsrød, R.; Fredriksen, B.N.; Lindmo, K.; Løkling, K.E.; Bordevik, M.; Steine, N.; Klevan, A.; Gravningen, K. Status and future perspectives of vaccines for industrialised fin-fish farming. Fish Shellfish Immunol. 2013, 35, 1759–1768. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Treatment Group | |
|---|---|---|
| Non-Vaccinated | Vaccinated | |
| Initial body weight (g) | 182.00 ± 32.09 | 182.40 ± 30.15 |
| Final body weight (g) | 500.67 ± 79.12 | 680.00 ± 126.54 * |
| Survival rate (%) | 67.7 ± 3.80 | 71.3 ± 0.05 |
| Weight gain (g) | 315.76 ± 34.79 | 498.94 ± 32.24 * |
| Specific growth rate (% day) | 0.90 ± 0.02 | 1.18 ± 0.05 * |
| Feed conversion ratio | 4.02 ± 0.44 | 2.78 ± 0.18 * |
| Feed efficiency | 0.25 ± 0.03 | 0.36 ± 0.02 * |
| Week | Gut | |
|---|---|---|
| Non-Vaccinated (%) | Vaccinated (%) | |
| 0 | 0.00 | 0.00 |
| 2 | 46.67 | 20.00 |
| 4 | 53.33 | 26.67 |
| 6 | 46.67 | 26.67 |
| 8 | 46.67 | 26.67 |
| 10 | 40.00 | 26.67 |
| 14 | 60.00 | 26.67 |
| 16 | 46.67 | 26.67 |
| Mean ± SD | 46.67 ± 7.91 * | 22.50 ± 9.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amir-Danial, Z.; Zamri-Saad, M.; Amal, M.N.A.; Annas, S.; Mohamad, A.; Jumria, S.; Manchanayake, T.; Arbania, A.; Ina-Salwany, M.Y. Field Efficacy of a Feed-Based Inactivated Vaccine against Vibriosis in Cage-Cultured Asian Seabass, Lates calcarifer, in Malaysia. Vaccines 2023, 11, 9. https://doi.org/10.3390/vaccines11010009
Amir-Danial Z, Zamri-Saad M, Amal MNA, Annas S, Mohamad A, Jumria S, Manchanayake T, Arbania A, Ina-Salwany MY. Field Efficacy of a Feed-Based Inactivated Vaccine against Vibriosis in Cage-Cultured Asian Seabass, Lates calcarifer, in Malaysia. Vaccines. 2023; 11(1):9. https://doi.org/10.3390/vaccines11010009
Chicago/Turabian StyleAmir-Danial, Zahaludin, Mohd Zamri-Saad, Mohammad Noor Azmai Amal, Salleh Annas, Aslah Mohamad, Sutra Jumria, Tilusha Manchanayake, Ali Arbania, and Md Yasin Ina-Salwany. 2023. "Field Efficacy of a Feed-Based Inactivated Vaccine against Vibriosis in Cage-Cultured Asian Seabass, Lates calcarifer, in Malaysia" Vaccines 11, no. 1: 9. https://doi.org/10.3390/vaccines11010009