
The Correlation between Subolesin-Reactive Epitopes and Vaccine Efficacy

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
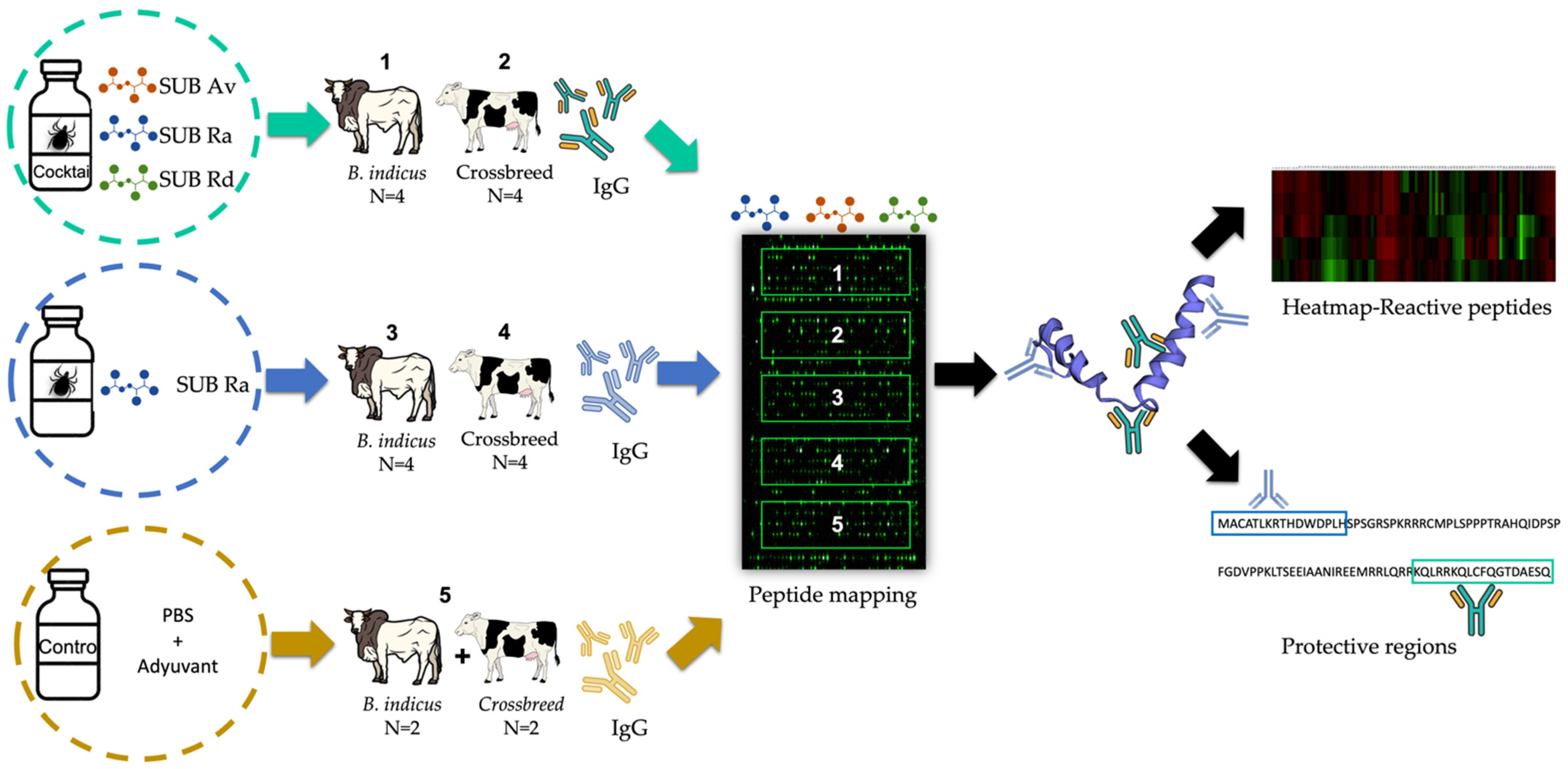
2.1. Serum Samples
2.2. Tick R. appendiculatus, R. decoloratus, and A. variegatum SUB Epitopes Microarray
2.3. Data Analysis and Peptide Characterization
3. Results
3.1. Rationale Sample Selection

3.2. Reactivity of Cattle Serum Samples against Subolesin Peptides
3.3. Characterization of the Reactive SUB Peptides by Quantum Vaccinomics Approach and Vaccine Protection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Lyme Borreliosis (Lyme Disease). Available online: https://www.euro.who.int/__data/assets/pdf_file/0008/246167/Fact-sheet-Lyme-borreliosis-Eng.pdf (accessed on 13 August 2022).
- Donohoe, H.; Pennington-Gray, L.; Omodior, O. Lyme disease: Current issues, implications, and recommendations for tourism management. Tour. Manag. 2015, 46, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Peter, R.J.; Van den Bossche, P.; Penzhorn, B.L.; Sharp, B. Tick, fly, and mosquito control—Lessons from the past, solutions for the future. Vet. Parasitol. 2005, 132, 205–215. [Google Scholar] [CrossRef]
- Hurtado, O.J.B.; Giraldo-Ríos, C. Economic and Health Impact of the Ticks in Production Animals; IntechOpen: London, UK, 2018; ISBN 978-1-78985-766-5. [Google Scholar]
- Shyma, K.P.; Gupta, J.P.; Singh, V. Breeding strategies for tick resistance in tropical cattle: A sustainable approach for tick control. J. Parasit. Dis. Off. Organ Indian Soc. Parasitol. 2015, 39, 1–6. [Google Scholar] [CrossRef]
- Mapholi, N.O.; Marufu, M.C.; Maiwashe, A.; Banga, C.B.; Muchenje, V.; MacNeil, M.D.; Chimonyo, M.; Dzama, K. Towards a genomics approach to tick (Acari: Ixodidae) control in cattle: A review. Ticks Tick-Borne Dis. 2014, 5, 475–483. [Google Scholar] [CrossRef]
- Groot, M.J.; Van’t Hooft, K.E. The Hidden Effects of Dairy Farming on Public and Environmental Health in the Netherlands, India, Ethiopia, and Uganda, Considering the Use of Antibiotics and Other Agro-chemicals. Front. Public Health 2016, 4, 12. [Google Scholar] [CrossRef]
- Titus, R.G.; Bishop, J.V.; Mejia, J.S. The immunomodulatory factors of arthropod saliva and the potential for these factors to serve as vaccine targets to prevent pathogen transmission. Parasite Immunol. 2006, 28, 131–141. [Google Scholar] [CrossRef]
- Labarta, V.; Rodríguez, M.; Penichet, M.; Lleonart, R.; Luaces, L.L.; de la Fuente, J. Simulation of control strategies for the cattle tick Boophilus microplus employing vaccination with a recombinant Bm86 antigen preparation. Vet. Parasitol. 1996, 63, 131–160. [Google Scholar] [CrossRef]
- Ndawula, C.; Sabadin, G.A.; Parizi, L.F.; da Silva Vaz, I. Constituting a glutathione S-transferase-cocktail vaccine against tick infestation. Vaccine 2019, 37, 1918–1927. [Google Scholar] [CrossRef]
- van Oosterwijk, J.G. Anti-tick and pathogen transmission blocking vaccines. Parasite Immunol. 2021, 43, e12831. [Google Scholar] [CrossRef] [PubMed]
- Mihajlović, J.; Hovius, J.W.R.; Sprong, H.; Bogovič, P.; Postma, M.J.; Strle, F. Cost-effectiveness of a potential anti-tick vaccine with combined protection against Lyme borreliosis and tick-borne encephalitis in Slovenia. Ticks Tick-Borne Dis. 2019, 10, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Byaruhanga, C.; Collins, N.E.; Knobel, D.; Kabasa, W.; Oosthuizen, M.C. Endemic status of tick-borne infections and tick species diversity among transhumant zebu cattle in Karamoja Region, Uganda: Support for control approaches. Vet. Parasitol. Reg. Stud. Rep. 2015, 1–2, 21–30. [Google Scholar] [CrossRef]
- Chenyambuga, S.; Waiswa, C.; Saimo, M.; Ngumi, P.; Gwakisa, P. Knowledge and perceptions of traditional livestock keepers on tick-borne diseases and sero-prevalence of Theileria parva around Lake Victoria basin. Livest. Res. Rural. Dev. 2010, 22, 135. [Google Scholar]
- Vudriko, P.; Okwee-Acai, J.; Tayebwa, D.S.; Byaruhanga, J.; Kakooza, S.; Wampande, E.; Omara, R.; Muhindo, J.B.; Tweyongyere, R.; Owiny, D.O.; et al. Emergence of multi-acaricide resistant Rhipicephalus ticks and its implication on chemical tick control in Uganda. Parasit. Vectors 2016, 9, 4. [Google Scholar] [CrossRef]
- Ocaido, M.; Muwazi, R.T.; Opuda, J.A. Economic impact of ticks and tick-borne diseases on cattle production systems around Lake Mburo National Park in South Western Uganda. Trop. Anim. Health Prod. 2009, 41, 731–739. [Google Scholar] [CrossRef]
- Mugabi, K.N.; Mugisha, A.; Ocaido, M. Socio-economic factors influencing the use of acaricides on livestock: A case study of the pastoralist communities of Nakasongola District, Central Uganda. Trop. Anim. Health Prod. 2010, 42, 131–136. [Google Scholar] [CrossRef]
- Goto, A.; Matsushita, K.; Gesellchen, V.; El Chamy, L.; Kuttenkeuler, D.; Takeuchi, O.; Hoffmann, J.A.; Akira, S.; Boutros, M.; Reichhart, J.-M. Akirins are highly conserved nuclear proteins required for NF-kappaB-dependent gene expression in drosophila and mice. Nat. Immunol. 2008, 9, 97–104. [Google Scholar] [CrossRef]
- de la Fuente, J.; Maritz-Olivier, C.; Naranjo, V.; Ayoubi, P.; Nijhof, A.M.; Almazán, C.; Canales, M.; de la Lastra, J.M.P.; Galindo, R.C.; Blouin, E.F.; et al. Evidence of the role of tick subolesin in gene expression. BMC Genom. 2008, 9, 372. [Google Scholar] [CrossRef]
- Nowak, S.J.; Aihara, H.; Gonzalez, K.; Nibu, Y.; Baylies, M.K. Akirin links twist-regulated transcription with the Brahma chromatin remodeling complex during embryogenesis. PLoS Genet. 2012, 8, e1002547. [Google Scholar] [CrossRef]
- Almazán, C.; Kocan, K.M.; Bergman, D.K.; Garcia-Garcia, J.C.; Blouin, E.F.; de la Fuente, J. Identification of protective antigens for the control of Ixodes scapularis infestations using cDNA expression library immunization. Vaccine 2003, 21, 1492–1501. [Google Scholar] [CrossRef]
- Bensaci, M.; Bhattacharya, D.; Clark, R.; Hu, L.T. Oral vaccination with vaccinia virus expressing the tick antigen subolesin inhibits tick feeding and transmission of Borrelia burgdorferi. Vaccine 2012, 30, 6040–6046. [Google Scholar] [CrossRef] [PubMed]
- Kasaija, P.D.; Contreras, M.; Kabi, F.; Mugerwa, S.; de la Fuente, J. Vaccination with Recombinant Subolesin Antigens Provides Cross-Tick Species Protection in Bos indicus and Crossbred Cattle in Uganda. Vaccines 2020, 8, 319. [Google Scholar] [CrossRef]
- Moreno-Cid, J.A.; de la Lastra, J.M.P.; Villar, M.; Jiménez, M.; Pinal, R.; Estrada-Peña, A.; Molina, R.; Lucientes, J.; Gortázar, C.; de la Fuente, J.; et al. Control of multiple arthropod vector infestations with subolesin/akirin vaccines. Vaccine 2013, 31, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Formstecher, E.; Aresta, S.; Collura, V.; Hamburger, A.; Meil, A.; Trehin, A.; Reverdy, C.; Betin, V.; Maire, S.; Brun, C.; et al. Protein interaction mapping: A Drosophila case study. Genome Res. 2005, 15, 376–384. [Google Scholar] [CrossRef]
- Rain, J.C.; Selig, L.; De Reuse, H.; Battaglia, V.; Reverdy, C.; Simon, S.; Lenzen, G.; Petel, F.; Wojcik, J.; Schächter, V.; et al. The protein-protein interaction map of Helicobacter pylori. Nature 2001, 409, 211–215. [Google Scholar] [CrossRef]
- Artigas-Jerónimo, S.; Comín, J.J.P.; Villar, M.; Contreras, M.; Alberdi, P.; Viera, I.L.; Soto, L.; Cordero, R.; Valdés, J.J.; Cabezas-Cruz, A.; et al. A Novel Combined Scientific and Artistic Approach for the Advanced Characterization of Interactomes: The Akirin/Subolesin Model. Vaccines 2020, 8, 77. [Google Scholar] [CrossRef]
- Rawal, K.; Sinha, R.; Abbasi, B.A.; Chaudhary, A.; Nath, S.K.; Kumari, P.; Preeti, P.; Saraf, D.; Singh, S.; Mishra, K.; et al. Identification of vaccine targets in pathogens and design of a vaccine using computational approaches. Sci. Rep. 2021, 11, 17626. [Google Scholar] [CrossRef]
- Safavi, A.; Kefayat, A.; Mahdevar, E.; Abiri, A.; Ghahremani, F. Exploring the out of sight antigens of SARS-CoV-2 to design a candidate multi-epitope vaccine by utilizing immunoinformatics approaches. Vaccine 2020, 38, 7612–7628. [Google Scholar] [CrossRef]
- Poland, G.A.; Ovsyannikova, I.G.; Jacobson, R.M.; Smith, D.I. Heterogeneity in vaccine immune response: The role of immunogenetics and the emerging field of vaccinomics. Clin. Pharmacol. Ther. 2007, 82, 653–664. [Google Scholar] [CrossRef]
- Bragazzi, N.L.; Gianfredi, V.; Villarini, M.; Rosselli, R.; Nasr, A.; Hussein, A.; Martini, M.; Behzadifar, M. Vaccines Meet Big Data: State-of-the-Art and Future Prospects. From the Classical 3Is (“Isolate-Inactivate-Inject”) Vaccinology 1.0 to Vaccinology 3.0, Vaccinomics, and Beyond: A Historical Overview. Front. Public Health 2018, 6, 62. [Google Scholar] [CrossRef] [PubMed]
- Gershoni, J.M.; Roitburd-Berman, A.; Siman-Tov, D.D.; Tarnovitski Freund, N.; Weiss, Y. Epitope mapping: The first step in developing epitope-based vaccines. BioDrugs Clin. Immunother. Biopharm. Gene Ther. 2007, 21, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Irving, M.B.; Pan, O.; Scott, J.K. Random-peptide libraries and antigen-fragment libraries for epitope mapping and the development of vaccines and diagnostics. Curr. Opin. Chem. Biol. 2001, 5, 314–324. [Google Scholar] [CrossRef]
- Contreras, M.; Artigas-Jerónimo, S.; Pastor Comín, J.J.; de la Fuente, J. A Quantum Vaccinomics Approach Based on Protein-Protein Interactions. Methods Mol. Biol. Clifton N. J. 2022, 2411, 287–305. [Google Scholar]
- Roggen, E.L. B-cell epitope engineering: A matter of recognizing protein features and motives. Drug Discov. Today Technol. 2008, 5, e49–e55. [Google Scholar] [CrossRef]
- Lin, J.; Sampson, H.A. IgE Epitope Mapping Using Peptide Microarray Immunoassay. Methods Mol. Biol. (Clifton N. J.) 2017, 1592, 177–187. [Google Scholar]
- Cheadle, C.; Vawter, M.P.; Freed, W.J.; Becker, K.G. Analysis of microarray data using Z score transformation. J. Mol. Diagn. JMD 2003, 5, 73–81. [Google Scholar] [CrossRef]
- Dennis, J.; Walker, L.; Tyrer, J.; Michailidou, K.; Easton, D.F. Detecting rare copy number variants from Illumina genotyping arrays with the CamCNV pipeline: Segmentation of z-scores improves detection and reliability. Genet. Epidemiol. 2021, 45, 237–248. [Google Scholar] [CrossRef]
- de la Fuente, J.; Contreras, M. Vaccinomics: A future avenue for vaccine development against emerging pathogens. Expert Rev. Vaccines 2021, 20, 1561–1569. [Google Scholar] [CrossRef]
- Leitão, J.H.; Rodríguez-Ortega, M.J. Omics and Bioinformatics Approaches to Identify Novel Antigens for Vaccine Investigation and Development. Vaccines 2020, 8, 653. [Google Scholar] [CrossRef]
- Papageorgiou, A.C.; Mohsin, I. The SARS-CoV-2 Spike Glycoprotein as a Drug and Vaccine Target: Structural Insights into Its Complexes with ACE2 and Antibodies. Cells 2020, 9, 2343. [Google Scholar] [CrossRef] [PubMed]
- Bencurova, E.; Gupta, S.K.; Oskoueian, E.; Bhide, M.; Dandekar, T. Omics and bioinformatics applied to vaccine development against Borrelia. Mol. Omics 2018, 14, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Thakur, A.; Katoch, S.; Shekhar, C.; Wani, A.H.; Kumar, S.; Dohroo, S.; Singh, G.; Sharma, M. Differences in innate and adaptive immune response traits of Pahari (Indian non-descript indigenous breed) and Jersey crossbred cattle. Vet. Immunol. Immunopathol. 2017, 192, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Robbertse, L.; Richards, S.A.; Clift, S.J.; Barnard, A.-C.; Leisewitz, A.; Crafford, J.E.; Maritz-Olivier, C. Comparison of the differential regulation of T and B-lymphocyte subsets in the skin and lymph nodes amongst three cattle breeds as potential mediators of immune-resistance to Rhipicephalus microplus. Ticks Tick-Borne Dis. 2018, 9, 976–987. [Google Scholar] [CrossRef]
- Legutki, J.B.; Johnston, S.A. Immunosignatures can predict vaccine efficacy. Proc. Natl. Acad. Sci. USA 2013, 110, 18614–18619. [Google Scholar] [CrossRef]
- Sharon, J.; Rynkiewicz, M.J.; Lu, Z.; Yang, C.-Y. Discovery of protective B-cell epitopes for development of antimicrobial vaccines and antibody therapeutics. Immunology 2014, 142, 1–23. [Google Scholar] [CrossRef]
- Wang, P.; Xiong, X.; Jiao, J.; Yang, X.; Jiang, Y.; Wen, B.; Gong, W. Th1 epitope peptides induce protective immunity against Rickettsia rickettsii infection in C3H/HeN mice. Vaccine 2017, 35, 7204–7212. [Google Scholar] [CrossRef]
- Ortega-Tirado, D.; Arvizu-Flores, A.A.; Velazquez, C.; Garibay-Escobar, A. The role of immunoinformatics in the development of T-cell peptide-based vaccines against Mycobacterium tuberculosis. Expert Rev. Vaccines 2020, 19, 831–841. [Google Scholar] [CrossRef]
- Loeffler, F.F.; Pfeil, J.; Heiss, K. High-Density Peptide Arrays for Malaria Vaccine Development. Methods Mol. Biol. (Clifton N. J.) 2016, 1403, 569–582. [Google Scholar]
- Reineke, U.; Schutkowski, M. Epitope mapping protocols. Preface. Methods Mol. Biol. (Clifton N. J.) 2009, 524, v–vi. [Google Scholar]
- Ramaraj, T.; Angel, T.; Dratz, E.A.; Jesaitis, A.J.; Mumey, B. Antigen-antibody interface properties: Composition, residue interactions, and features of 53 non-redundant structures. Biochim. Biophys. Acta 2012, 1824, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Van Regenmortel, M.H.V. What is a B-cell epitope? Methods Mol. Biol. (Clifton N. J.) 2009, 524, 3–20. [Google Scholar]
- Prudencio, C.R.; de la Lastra, J.M.P.; Canales, M.; Villar, M.; de la Fuente, J. Mapping protective epitopes in the tick and mosquito subolesin ortholog proteins. Vaccine 2010, 28, 5398–5406. [Google Scholar] [CrossRef] [PubMed]
- Watanabe Costa, R.; Batista, M.F.; Meneghelli, I.; Vidal, R.O.; Nájera, C.A.; Mendes, A.C.; Andrade-Lima, I.A.; da Silveira, J.F.; Lopes, L.R.; Ferreira, L.R.P.; et al. Comparative Analysis of the Secretome and Interactome of Trypanosoma cruzi and Trypanosoma rangeli Reveals Species Specific Immune Response Modulating Proteins. Front. Immunol. 2020, 11, 1774. [Google Scholar] [CrossRef] [PubMed]
- Mujawar, S.; Mishra, R.; Pawar, S.; Gatherer, D.; Lahiri, C. Delineating the Plausible Molecular Vaccine Candidates and Drug Targets of Multidrug-Resistant Acinetobacter baumannii. Front. Cell. Infect. Microbiol. 2019, 9, 203. [Google Scholar] [CrossRef]
- Canales, M.; Naranjo, V.; Almazán, C.; Molina, R.; Tsuruta, S.A.; Szabó, M.P.J.; Manzano-Roman, R.; de la Lastra, J.M.P.; Kocan, K.M.; Jiménez, M.I.; et al. Conservation and immunogenicity of the mosquito ortholog of the tick-protective antigen, subolesin. Parasitol. Res. 2009, 105, 97–111. [Google Scholar] [CrossRef]
- de la Fuente, J.; Moreno-Cid, J.A.; Galindo, R.C.; Almazan, C.; Kocan, K.M.; Merino, O.; de la Lastra, J.M.P.; Estrada-Peña, A.; Blouin, E.F. Subolesin/Akirin vaccines for the control of arthropod vectors and vectorborne pathogens. Transbound. Emerg. Dis. 2013, 60 (Suppl. S2), 172–178. [Google Scholar] [CrossRef]
- Contreras, M.; de la Fuente, J. Control of Ixodes ricinus and Dermacentor reticulatus tick infestations in rabbits vaccinated with the Q38 Subolesin/Akirin chimera. Vaccine 2016, 34, 3010–3013. [Google Scholar] [CrossRef]
- Artigas-Jerónimo, S.; Villar, M.; Cabezas-Cruz, A.; Valdés, J.J.; Estrada-Peña, A.; Alberdi, P.; de la Fuente, J. Functional Evolution of Subolesin/Akirin. Front. Physiol. 2018, 9, 1612. [Google Scholar] [CrossRef]
- Morgan, W.D.; Frenkiel, T.A.; Lock, M.J.; Grainger, M.; Holder, A.A. Precise epitope mapping of malaria parasite inhibitory antibodies by TROSY NMR cross-saturation. Biochemistry 2005, 44, 518–523. [Google Scholar] [CrossRef]
- Zobayer, N.; Hossain, A.A.; Rahman, M.A. A combined view of B-cell epitope features in antigens. Bioinformation 2019, 15, 530–534. [Google Scholar] [CrossRef] [PubMed]
- De Groot, A.S.; Moise, L.; Terry, F.; Gutierrez, A.H.; Hindocha, P.; Richard, G.; Hoft, D.F.; Ross, T.M.; Noe, A.R.; Takahashi, Y.; et al. Better Epitope Discovery, Precision Immune Engineering, and Accelerated Vaccine Design Using Immunoinformatics Tools. Front. Immunol. 2020, 11, 442. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serum Groups | |||||||
|---|---|---|---|---|---|---|---|
| Peptide n° | Tick Species | Peptide Sequence | Control | Cocktail Crossbred | Cocktail B. indicus | SUBra Crossbred | SUBra B. indicus |
| 20 | A.v | 13DPLHSPNGRSPKRRR27 | x | ||||
| 21 | A.v | 14PLHSPNGRSPKRRRC28 | x | ||||
| 55 | A.v | 28SPFGDVPPKLTSEEI42 | † | ||||
| 79 | A.v | 72RLQRRKQLCFQGAEC86 | x † | † | |||
| 86 | A.v | 79LCFQGAECSSPPEGC93 | x † | x † | |||
| 88 | A.v | 81FQGAECSSPPEGCSP95 | † | ||||
| 110 | A.v | 103TSGLLSPVRRDQPLF117 | x † | x † | |||
| 112 | A.v | 105GLLSPVRRDQPLFTF119 | x | ||||
| 119 | A.v | 112RDQPLFTFRQVGLIC126 | x | ||||
| 123 | A.v | 116LFTFRQVGLICERMM130 | x | x | |||
| 124 | A.v | 117FTFRQVGLICERMMK131 | x | x | x | ||
| 125 | A.v | 118TFRQVGLICERMMKE132 | x | x | |||
| 126 | A.v | 119FRQVGLICERMMKER133 | x | x | x | ||
| 127 | A.v | 120RQVGLICERMMKERE134 | x | x | x | ||
| 128 | A.v | 121QVGLICERMMKERES135 | x | x | x | ||
| 129 | A.v | 122VGLICERMMKERESQ136 | x | ||||
| 130 | A.v | 123GLICERMMKERESQI137 | x | x | |||
| 132 | A.v | 125ICERMMKERESQIRE139 | x | ||||
| 142 | A.v | 135SQIREEYDHVLSTKL149 | x † | ||||
| 144 | A.v | 137IREEYDHVLSTKLAE151 | x † | x † | |||
| 146 | A.v | 139EEYDHVLSTKLAEQY153 | † | x † | † | ||
| 147 | A.v | 140EYDHVLSTKLAEQYD154 | † | † | x † | x † | |
| 149 | A.v | 142DHVLSTKLAEQYDTF156 | † | † | x † | x † | |
| 150 | A.v | 143HVLSTKLAEQYDTFV157 | † | x † | x † | ||
| Peptide n° | Tick species | Peptide sequence | Control | Cocktail Crossbred | Cocktail B. indicus | SUBra Crossbred | SUBra B. indicus |
| 152 | A.v | 145LSTKLAEQYDTFVKF159 | x † | x † | |||
| 19 | R.a | 12WDPLHSPSGRSPKRR26 | x | ||||
| 21 | R.a | 14PLHSPSGRSPKRRRC28 | x | x | |||
| 31 | R.a | 24KRRRCMPLSPPPTRA38 | x | ||||
| 35 | R.a | 28CMPLSPPPTRAHQID42 | † | ||||
| 69 | R.a | 62IREEMRRLQRRKQLC76 | x † | ||||
| 94 | R.a | 87TSGLSSPVRRDQPLF101 | x † | ||||
| 113 | R.a | 106VGLICERMMKERESK120 | x | ||||
| 114 | R.a | 107GLICERMMKERESKI121 | x | ||||
| 115 | R.a | 108LICERMMKERESKIR122 | x | x | |||
| 116 | R.a | 109ICERMMKERESKIRE123 | x | x | x | ||
| 117 | R.a | 110CERMMKERESKIREE124 | x | x | |||
| 118 | R.a | 111ERMMKERESKIREEY125 | x † | x | |||
| 119 | R.a | 112RMMKERESKIREEYD126 | x | ||||
| 128 | R.a | 121IREEYDHVLSTKLAE135 | † | x † | x † | ||
| 130 | R.a | 123EEYDHVLSTKLAEQY137 | † | † | x † | ||
| 131 | R.a | 124EYDHVLSTKLAEQYD138 | † | † | x † | x † | |
| 132 | R.a | 125YDHVLSTKLAEQYDT139 | † | ||||
| 133 | R.a | 126DHVLSTKLAEQYDTF140 | † | † | x † | x † | |
| 134 | R.a | 127HVLSTKLAEQYDTFV141 | † | x † | x † | ||
| 136 | R.a | 129LSTKLAEQYDTFVKF143 | x † | x † | |||
| 139 | R.a | 132KLAEQYDTFVKFTYD146 | † | ||||
| 141 | R.a | 134AEQYDTFVKFTYDQI148 | † | x † | † | ||
| 42 | R.d | 35PTRAHQIDPSPFGDV49 | † | ||||
| 43 | R.d | 36TRAHQIDPSPFGDVP50 | † | ||||
| 94 | R.d | 87TSGLSSPVHRDQPLF101 | x † | x † | |||
| 111 | R.d | 104RQVGLICERMMKERE118 | x | x | |||
| Peptide n° | Tick species | Peptide sequence | Control | Cocktail Crossbred | Cocktail B. indicus | SUBra Crossbred | SUBra B. indicus |
| 114 | R.d | 107GLICERMMKERESKI121 | x | ||||
| 115 | R.d | 108LICERMMKERESKIR122 | x | x | |||
| 116 | R.d | 109ICERMMKERESKIRE123 | x | x | |||
| 117 | R.d | 110CERMMKERESKIREE124 | x | x | x | ||
| 118 | R.d | 111ERMMKERESKIREEY125 | x | x | |||
| 119 | R.d | 112RMMKERESKIREEYD126 | x | ||||
| 120 | R.d | 113MMKERESKIREEYDH127 | x † | ||||
| 128 | R.d | 121IREEYDHVLSTKLAE135 | x † | x † | |||
| 130 | R.d | 123EEYDHVLSTKLAEQY137 | † | † | x † | ||
| 131 | R.d | 124EYDHVLSTKLAEQYD138 | † | † | † | † | |
| 132 | R.d | 125YDHVLSTKLAEQYDT139 | † | ||||
| 133 | R.d | 126DHVLSTKLAEQYDTF140 | † | † | x † | x † | |
| 134 | R.d | 127HVLSTKLAEQYDTFV141 | † | † | † | † | |
| 136 | R.d | 129LSTKLAEQYDTFVKF143 | x † | x † | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contreras, M.; Kasaija, P.D.; Kabi, F.; Mugerwa, S.; De la Fuente, J. The Correlation between Subolesin-Reactive Epitopes and Vaccine Efficacy. Vaccines 2022, 10, 1327. https://doi.org/10.3390/vaccines10081327
Contreras M, Kasaija PD, Kabi F, Mugerwa S, De la Fuente J. The Correlation between Subolesin-Reactive Epitopes and Vaccine Efficacy. Vaccines. 2022; 10(8):1327. https://doi.org/10.3390/vaccines10081327
Chicago/Turabian StyleContreras, Marinela, Paul D. Kasaija, Fredrick Kabi, Swidiq Mugerwa, and José De la Fuente. 2022. "The Correlation between Subolesin-Reactive Epitopes and Vaccine Efficacy" Vaccines 10, no. 8: 1327. https://doi.org/10.3390/vaccines10081327