
Long-Term Longitudinal Analysis of Neutralizing Antibody Response to Three Vaccine Doses in a Real-Life Setting of Previously SARS-CoV-2 Infected Healthcare Workers: A Model for Predicting Response to Further Vaccine Doses


 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
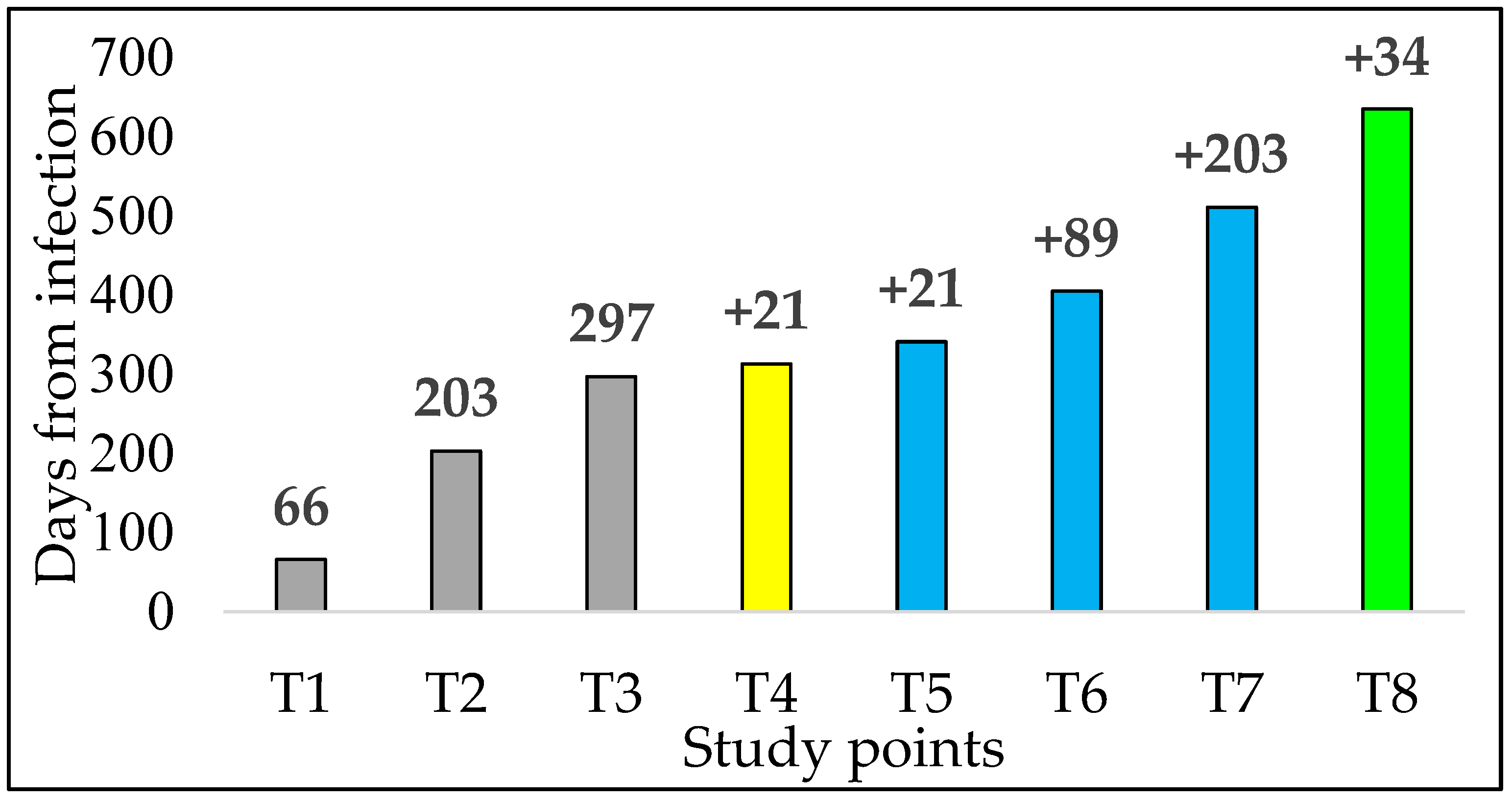
2.1. Study Design
2.2. Titration of Virus Neutralizing Antibodies
2.3. Statistical Analysis
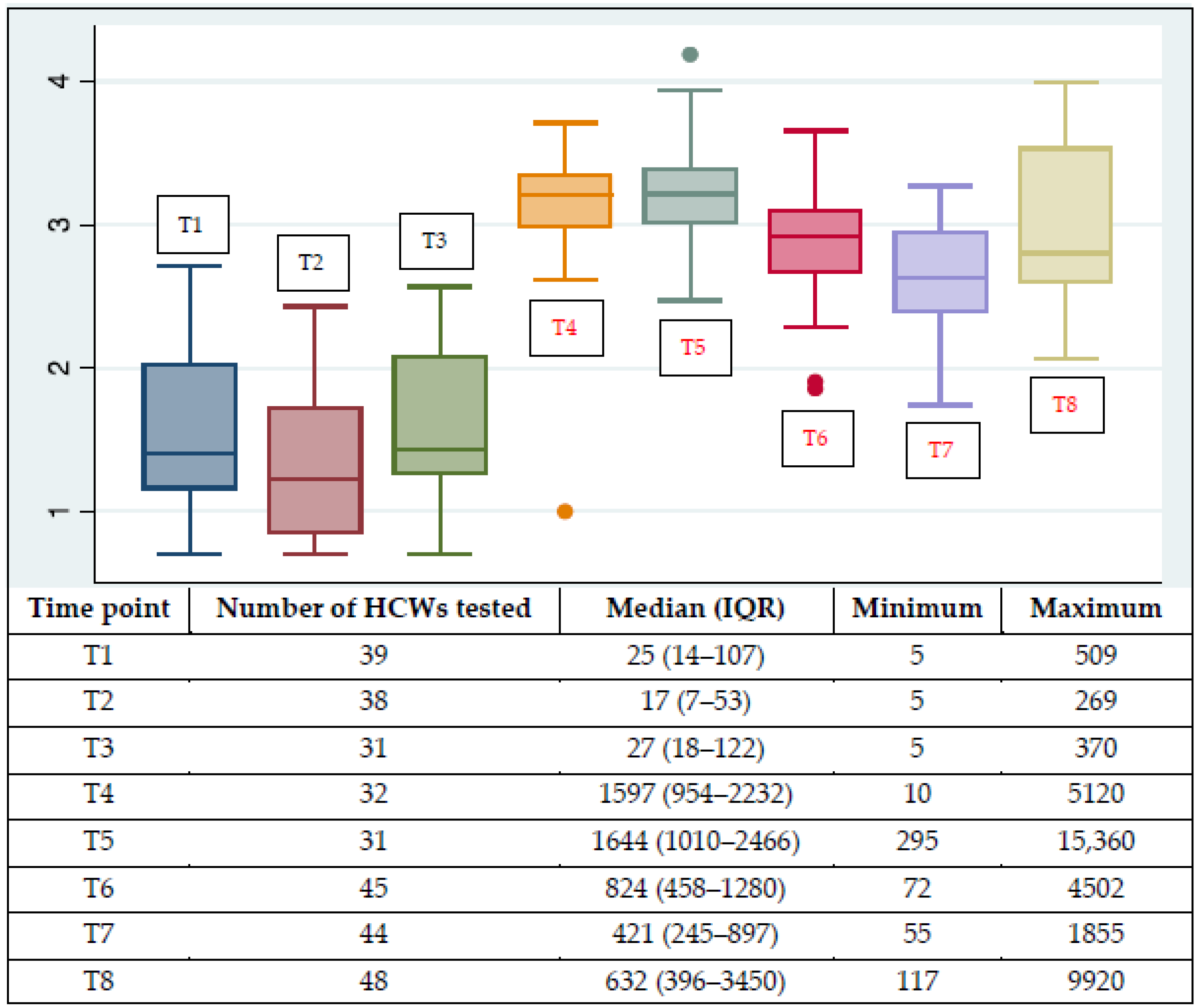
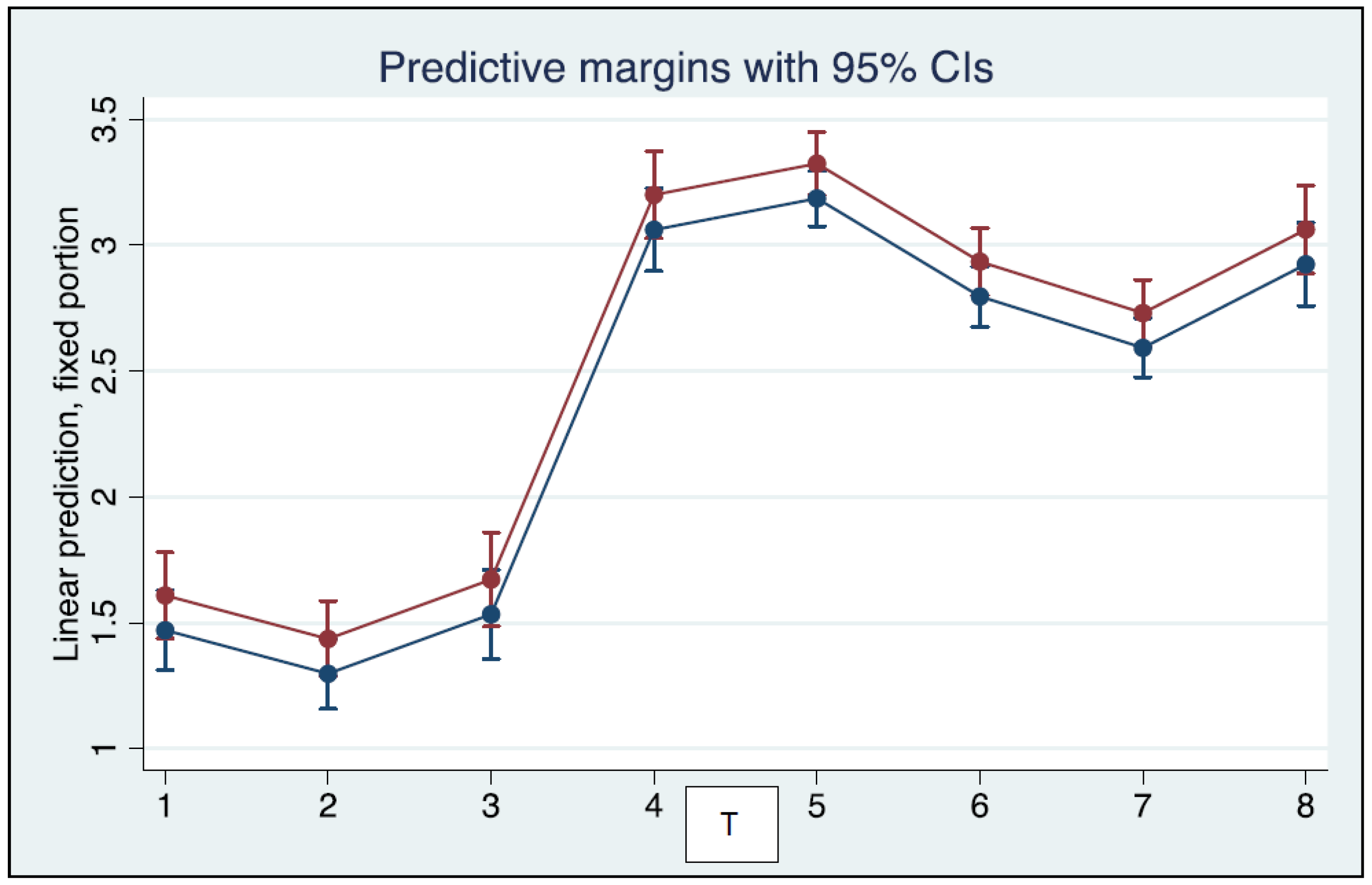
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoteit, R.; Yassine, H.M. Biological Properties of SARS-CoV-2 Variants: Epidemiological Impact and Clinical Consequences. Vaccines 2022, 10, 919. [Google Scholar] [CrossRef]
- WHO. Tracking SARS-CoV-2 Variants. Available online: www.who.int/en/activities/tracking-SARS-CoV-2-variants (accessed on 20 June 2022).
- WHO. SARS-CoV-2 Delta Variant Now Dominant in Much of European Region; Efforts Must Be Reinforced to Prevent Transmission, Warns WHO Regional Office for Europe and ECDC. 2021. Available online: www.euro.who.int/en/media-centre/sections/press-releases/2021/sars-cov-2-delta-variant-now-dominantin-much-of-european-region-efforts-must-be-reinforced-toprevent-transmission,-warns-who-regional-office-foreuropeand-ecdc (accessed on 20 June 2022).
- WHO Technical Advisory Group on SARS-CoV-2 Virus Evolution (TAG-VE). Classification of Omicron (B.1.1.529): SARS-CoV-2 Variant of Concern. 2021. Available online: www.who.int/news/item/26-11-2021-classification-of-omicron-(b.1.1.529)-sars-cov-2-variantof-concern (accessed on 20 June 2022).
- Patel, R.; Kaki, M.; Potluri, V.S.; Kahar, P.; Khanna, D. A comprehensive review of SARS-CoV-2 vaccines: Pfizer, Moderna & Johnson & Johnson. Hum. Vaccin. Immunother. 2022, 18, 2002083. [Google Scholar] [PubMed]
- Mohammed, I.; Nauman, A.; Paul, P.; Ganesan, S.; Chen, K.H.; Jalil, S.M.S.; Jaouni, S.H.; Kawas, H.; Khan, W.A.; Vattoth, A.L.; et al. The efficacy and effectiveness of the COVID-19 vaccines in reducing infection, severity, hospitalization, and mortality: A systematic review. Hum. Vaccin. Immunother. 2022, 18, 2027160. [Google Scholar] [CrossRef]
- Edara, V.V.; Manning, K.E.; Ellis, M.; Lai, L.; Moore, K.M.; Foster, S.L.; Floyd, K.; Davis-Gardner, M.E.; Mantus, G.; Nyhoff, L.E.; et al. mRNA-1273 and BNT162b2 mRNA vaccines have reduced neutralizing activity against the SARS-CoV-2 omicron variant. Cell Rep. Med. 2022, 3, 100529. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.J.; Moreira, E.D., Jr.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Polack, F.P.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine through 6 Months. N. Engl. J. Med. 2021, 385, 1761–1773. [Google Scholar] [CrossRef] [PubMed]
- Magen, O.; Waxman, J.G.; Makov-Assif, M.; Vered, R.; Dicker, D.; Hernán, M.A.; Lipsitch, M.; Reis, B.Y.; Balicer, R.D.; Dagan, N. Fourth Dose of BNT162b2 mRNA COVID-19 Vaccine in a Nationwide Setting. N. Engl. J. Med. 2022, 386, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Vicenti, I.; Gatti, F.; Scaggiante, R.; Boccuto, A.; Zago, D.; Basso, M.; Dragoni, F.; Zazzi, M.; Parisi, S.G. Single-dose BNT162b2 mRNA COVID-19 vaccine significantly boosts neutralizing antibody response in health care workers recovering from asymptomatic or mild natural SARS-CoV-2 infection. Int. J. Infect. Dis. 2021, 108, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Vicenti, I.; Gatti, F.; Scaggiante, R.; Boccuto, A.; Zago, D.; Basso, M.; Dragoni, F.; Parisi, S.G.; Zazzi, M. The second dose of the BNT162b2 mRNA vaccine does not boost SARS-CoV-2 neutralizing antibody response in previously infected subjects. Infection 2022, 50, 541–543. [Google Scholar] [CrossRef] [PubMed]
- Chenchula, S.; Karunakaran, P.; Sharma, S.; Chavan, M. Current evidence on efficacy of COVID-19 booster dose vaccination against the Omicron variant: A systematic review. J. Med. Virol. 2022, 94, 2969–2976. [Google Scholar] [CrossRef]
- Earle, K.A.; Ambrosino, D.M.; Fiore-Gartland, A.; Goldblatt, D.; Gilbert, P.B.; Siber, G.R.; Dull, P.; Plotkin, S.A. Evidence for antibody as a protective correlate for COVID-19 vaccines. Vaccine 2021, 39, 4423–4428. [Google Scholar] [CrossRef] [PubMed]
- Pieri, M.; Infantino, M.; Manfredi, M.; Nuccetelli, M.; Grossi, V.; Lari, B.; Tomassetti, F.; Sarubbi, S.; Russo, E.; Amedei, A.; et al. Performance evaluation of four surrogate Virus Neutralization Tests (sVNTs) in comparison to the in vivo gold standard test. Front. Biosci. Landmark Ed. 2022, 27, 74. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Clinical Management of Severe Acute Respiratory Infection When Novel Coronavirus (2019-nCoV) Infection Is Suspected: Interim Guidance, 28 January 2020. 2020. Available online: https://apps.who.int/iris/handle/10665/330893 (accessed on 20 June 2022).
- Vicenti, I.; Basso, M.; Dragoni, F.; Gatti, F.; Scaggiante, R.; Fiaschi, L.; Parisi, S.G.; Zazzi, M. Comparable Post-Vaccination Decay of Neutralizing Antibody Response to Wild-Type and Delta SARS-CoV-2 Variant in Healthcare Workers Recovered from Mild or Asymptomatic Infection. Vaccines 2022, 10, 580. [Google Scholar] [CrossRef]
- Yamayoshi, S.; Yasuhara, A.; Ito, M.; Akasaka, O.; Nakamura, M.; Nakachi, I.; Koga, M.; Mitamura, K.; Yagi, K.; Maeda, K.; et al. Antibody titers against SARS-CoV-2 decline, but do not disappear for several months. EClinicalMedicine 2021, 32, 100734. [Google Scholar] [CrossRef] [PubMed]
- Petrelli, F.; Luciani, A.; Borgonovo, K.; Ghilardi, M.; Parati, M.C.; Petrò, D.; Lonati, V.; Pesenti, A.; Cabiddu, M. Third dose of SARS-CoV-2 vaccine: A systematic review of 30 published studies. J. Med. Virol. 2022, 94, 2837–2844. [Google Scholar] [CrossRef] [PubMed]
- Romero-Ibarguengoitia, M.E.; Rivera-Salinas, D.; Hernández-Ruíz, Y.G.; Armendariz-Vázquez, A.G.; González-Cantú, A.; Barco-Flores, I.A.; González-Facio, R.; Montelongo-Cruz, L.P.; Del Rio-Parra, G.F.; Garza-Herrera, M.R.; et al. Effect of the third dose of BNT162b2 vaccine on quantitative SARS-CoV-2 spike 1-2 IgG antibody titers in healthcare personnel. PLoS ONE 2022, 17, e0263942. [Google Scholar] [CrossRef]
- Mannar, D.; Saville, J.W.; Zhu, X.; Srivastava, S.S.; Berezuk, A.M.; Tuttle, K.S.; Marquez, A.C.; Sekirov, I.; Subramaniam, S. SARS-CoV-2 Omicron variant: Antibody evasion and cryo-EM structure of spike protein-ACE2 complex. Science 2022, 375, 760–764. [Google Scholar] [CrossRef]
- Parums, D.V. Editorial: World Health Organization (WHO) Variants of Concern Lineages Under Monitoring (VOC-LUM) in Response to the Global Spread of Lineages and Sublineages of Omicron, or B.1.1.529, SARS-CoV-2. Med. Sci. Monit. 2022, 28, e937676. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Technical Report. Enhancing Response to Omicron SARS-CoV-2 Variant. 21 January 2022. Available online: https://www.who.int/publications/m/item/enhancing-readiness-for-omicron-(b.1.1.529)-technical-brief-and-priority-actions-for-member-states (accessed on 13 July 2022).
- Parums, D.V. Editorial: The 2022 World Health Organization (WHO) priority recommendations and response to the Omicron variant (B.1.1.529) of SARS-CoV-2. Med. Sci. Monit. 2022, 28, e936199. [Google Scholar] [CrossRef]
- Nemet, I.; Kliker, L.; Lustig, Y.; Zuckerman, N.; Erster, O.; Cohen, C.; Kreiss, Y.; Alroy-Preis, S.; Regev-Yochay, G.; Mendelson, E.; et al. Third BNT162b2 Vaccination Neutralization of SARS-CoV-2 Omicron Infection. N. Engl. J. Med. 2022, 386, 492–494. [Google Scholar] [CrossRef]
- Ariën, K.K.; Heyndrickx, L.; Michiels, J.; Vereecken, K.; Van Lent, K.; Coppens, S.; Willems, B.; Pannus, P.; Martens, G.A.; Van Esbroeck, M.; et al. Three doses of BNT162b2 vaccine confer neutralising antibody capacity against the SARS-CoV-2 Omicron variant. NPJ Vaccines 2022, 7, 35. [Google Scholar] [CrossRef] [PubMed]
- Carreño, J.M.; Alshammary, H.; Tcheou, J.; Singh, G.; Raskin, A.J.; Kawabata, H.; Sominsky, L.A.; Clark, J.J.; Adelsberg, D.C.; Bielak, D.A.; et al. Activity of convalescent and vaccine serum against SARS-CoV-2 Omicron. Nature 2022, 602, 682–688. [Google Scholar] [CrossRef]
- Blom, K.; Marking, U.; Havervall, S.; Norin, N.G.; Gordon, M.; García, M.; Tecleab, T.; Christ, W.; Forsell, M.; Phillipson, M.; et al. Immune responses after omicron infection in triple-vaccinated health-care workers with and without previous SARS-CoV-2 infection. Lancet Infect. Dis. 2022, 7, 943–945. [Google Scholar] [CrossRef]
- Hachmann, N.P.; Miller, J.; Collier, A.Y.; Ventura, J.D.; Yu, J.; Rowe, M.; Bondzie, E.A.; Powers, O.; Surve, N.; Hall, K.; et al. Neutralization Escape by SARS-CoV-2 Omicron Subvariants BA.2.12.1, BA.4, and BA.5. N. Engl. J. Med. 2022, 387, 86–88. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Yisimayi, A.; Jian, F.; Song, W.; Xiao, T.; Wang, L.; Du, S.; Wang, J.; Li, Q.; Chen, X.; et al. BA.2.12.1, BA.4 and BA.5 escape antibodies elicited by Omicron infection. Nature, 2022; epub ahead of print. [Google Scholar] [CrossRef]
- Goldblatt, D.; Fiore-Gartland, A.; Johnson, M.; Hunt, A.; Bengt, C.; Zavadska, D.; Snipe, H.D.; Brown, J.S.; Workman, L.; Zar, H.J.; et al. Towards a population-based threshold of protection for COVID-19 vaccines. Vaccine 2022, 40, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Singanayagam, A.; Hakki, S.; Dunning, J.; Madon, K.J.; Crone, M.A.; Koycheva, A.; Derqui-Fernandez, N.; Barnett, J.L.; Whitfield, M.G.; Varro, R.; et al. Community transmission and viral load kinetics of the SARS-CoV-2 delta (B.1.617.2) variant in vaccinated and unvaccinated individuals in the UK: A prospective, longitudinal, cohort study. Lancet Infect. Dis. 2022, 22, 183–195. [Google Scholar] [CrossRef]
- Guo, Y.; Han, J.; Zhang, Y.; He, J.; Yu, W.; Zhang, X.; Wu, J.; Zhang, S.; Kong, Y.; Guo, Y.; et al. SARS-CoV-2 Omicron Variant: Epidemiological Features, Biological Characteristics, and Clinical Significance. Front. Immunol. 2022, 13, 877101. [Google Scholar] [CrossRef]
- Mantus, G.; Nyhoff, L.E.; Edara, V.V.; Zarnitsyna, V.I.; Ciric, C.R.; Flowers, M.W.; Norwood, C.; Ellis, M.; Hussaini, L.; Manning, K.E.; et al. Pre-existing SARS-CoV-2 immunity influences potency, breadth, and durability of the humoral response to SARS-CoV-2 vaccination. Cell Rep. Med. 2022, 3, 100603. [Google Scholar] [CrossRef]
- Lin, K.; Zhou, Y.; Ai, J.; Wang, Y.A.; Zhang, S.; Qiu, C.; Lian, C.; Gao, B.; Liu, T.; Wang, H.; et al. B cell receptor signatures associated with strong and poor SARS-CoV-2 vaccine responses. Emerg. Microbes Infect. 2022, 11, 452–464. [Google Scholar] [CrossRef]
- Deb, P.; Zannat, K.E.; Talukder, S.; Bhuiyan, A.H.; Jilani, M.S.A.; Saif-Ur-Rahman, K.M. Association of HLA gene polymorphism with susceptibility, severity, and mortality of COVID-19: A systematic review. HLA 2022, 99, 281–312. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| log10 Antibody Level | ||||||
|---|---|---|---|---|---|---|
| At Time point | Coefficient | Std. Err. | z | p | 95% Conf. Interval | |
| T2 | −0.1728 | 0.0929 | −1.86 | 0.063 | −0.3549 | 0.0093 |
| T3 | 0.0632 | 0.0935 | 0.68 | 0.499 | −0.1202 | 0.2465 |
| T4 | 1.5908 | 0.0798 | 19.94 | 0.000 | 1.4345 | 1.7471 |
| T5 | 1.7150 | 0.0843 | 20.34 | 0.000 | 1.5497 | 1.8802 |
| T6 | 1.3259 | 0.0780 | 17.00 | 0.000 | 1.1730 | 1.4788 |
| T7 | 1.1215 | 0.0756 | 14.84 | 0.000 | 0.9733 | 1.2696 |
| T8 | 1.4534 | 0.0963 | 15.09 | 0.000 | 1.2647 | 1.6421 |
| other variables | ||||||
| female | −0.0388 | 0.0698 | −0.56 | 0.579 | −0.1756 | 0.0981 |
| age | −0.0076 | 0.0036 | −2.14 | 0.033 | −0.0146 | −0.0006 |
| Symptomatic infection | 0.1672 | 0.0679 | 2.46 | 0.014 | 0.0342 | 0.3002 |
| intercept | 1.8340 | 0.1739 | 10.54 | 0.000 | 1.4931 | 2.1749 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parisi, S.G.; Mengoli, C.; Basso, M.; Vicenti, I.; Gatti, F.; Scaggiante, R.; Fiaschi, L.; Giammarino, F.; Iannetta, M.; Malagnino, V.; et al. Long-Term Longitudinal Analysis of Neutralizing Antibody Response to Three Vaccine Doses in a Real-Life Setting of Previously SARS-CoV-2 Infected Healthcare Workers: A Model for Predicting Response to Further Vaccine Doses. Vaccines 2022, 10, 1237. https://doi.org/10.3390/vaccines10081237
Parisi SG, Mengoli C, Basso M, Vicenti I, Gatti F, Scaggiante R, Fiaschi L, Giammarino F, Iannetta M, Malagnino V, et al. Long-Term Longitudinal Analysis of Neutralizing Antibody Response to Three Vaccine Doses in a Real-Life Setting of Previously SARS-CoV-2 Infected Healthcare Workers: A Model for Predicting Response to Further Vaccine Doses. Vaccines. 2022; 10(8):1237. https://doi.org/10.3390/vaccines10081237
Chicago/Turabian StyleParisi, Saverio Giuseppe, Carlo Mengoli, Monica Basso, Ilaria Vicenti, Francesca Gatti, Renzo Scaggiante, Lia Fiaschi, Federica Giammarino, Marco Iannetta, Vincenzo Malagnino, and et al. 2022. "Long-Term Longitudinal Analysis of Neutralizing Antibody Response to Three Vaccine Doses in a Real-Life Setting of Previously SARS-CoV-2 Infected Healthcare Workers: A Model for Predicting Response to Further Vaccine Doses" Vaccines 10, no. 8: 1237. https://doi.org/10.3390/vaccines10081237