
Activation of Anti-SARS-CoV-2 Human CTLs by Extracellular Vesicles Engineered with the N Viral Protein

 , , , and
, , , and 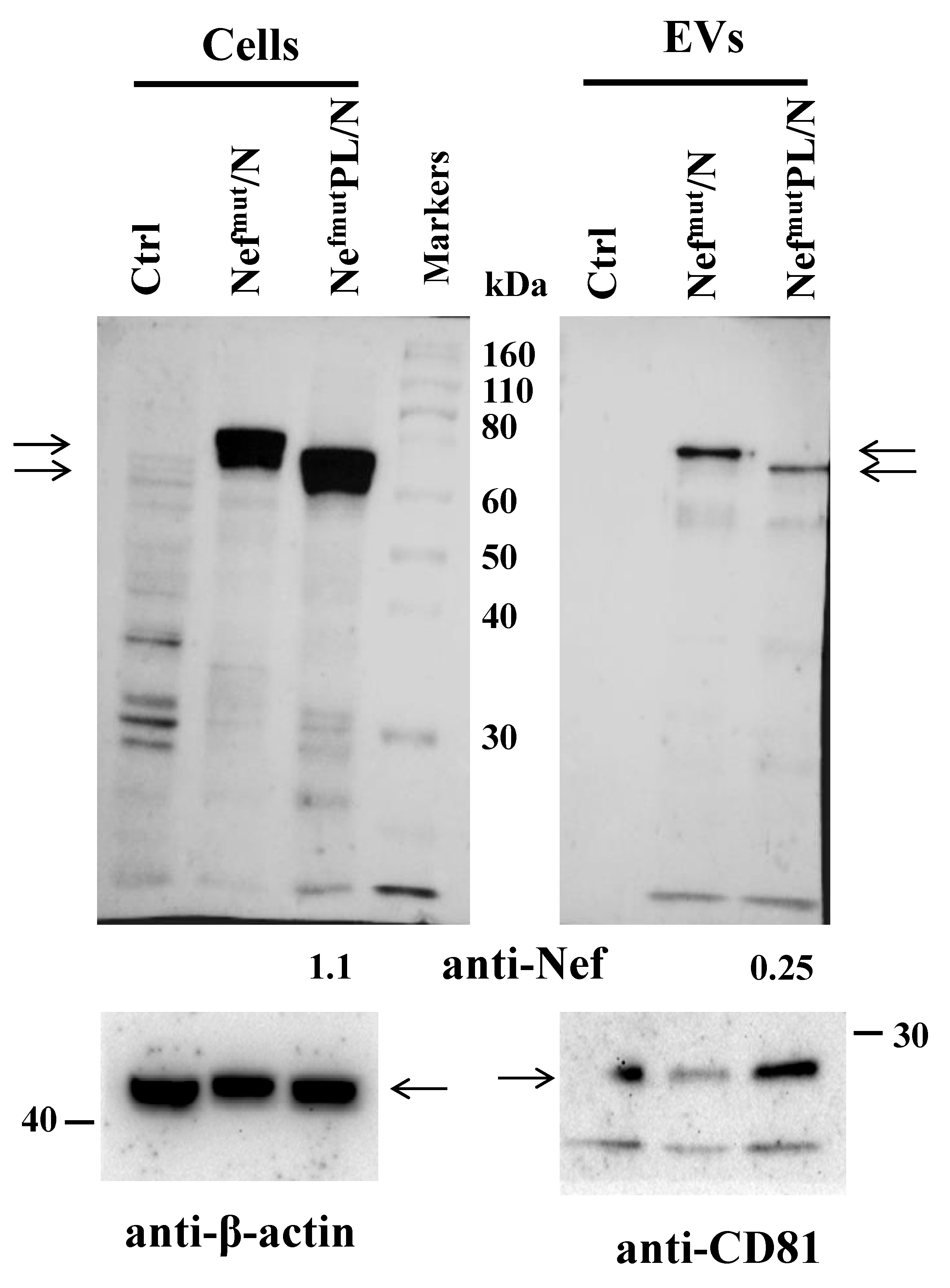{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. DNA Vector Synthesis
2.2. Cell Cultures and Transfection
2.3. EV Isolation
2.4. Western Blot Analysis
2.5. Mice Immunization
2.6. Cell Isolation from Immunized Mice
2.7. IFN-γ EliSpot Analysis
2.8. NTA Analysis of Nanovesicles
2.9. Cross-Priming Assay
2.10. Activation-Induced Degranulation and Trogocytosis Assays
2.11. Statistical Analysis
3. Results
3.1. EV Uploading of SARS-CoV-2 N upon Fusion with Nefmut/PL
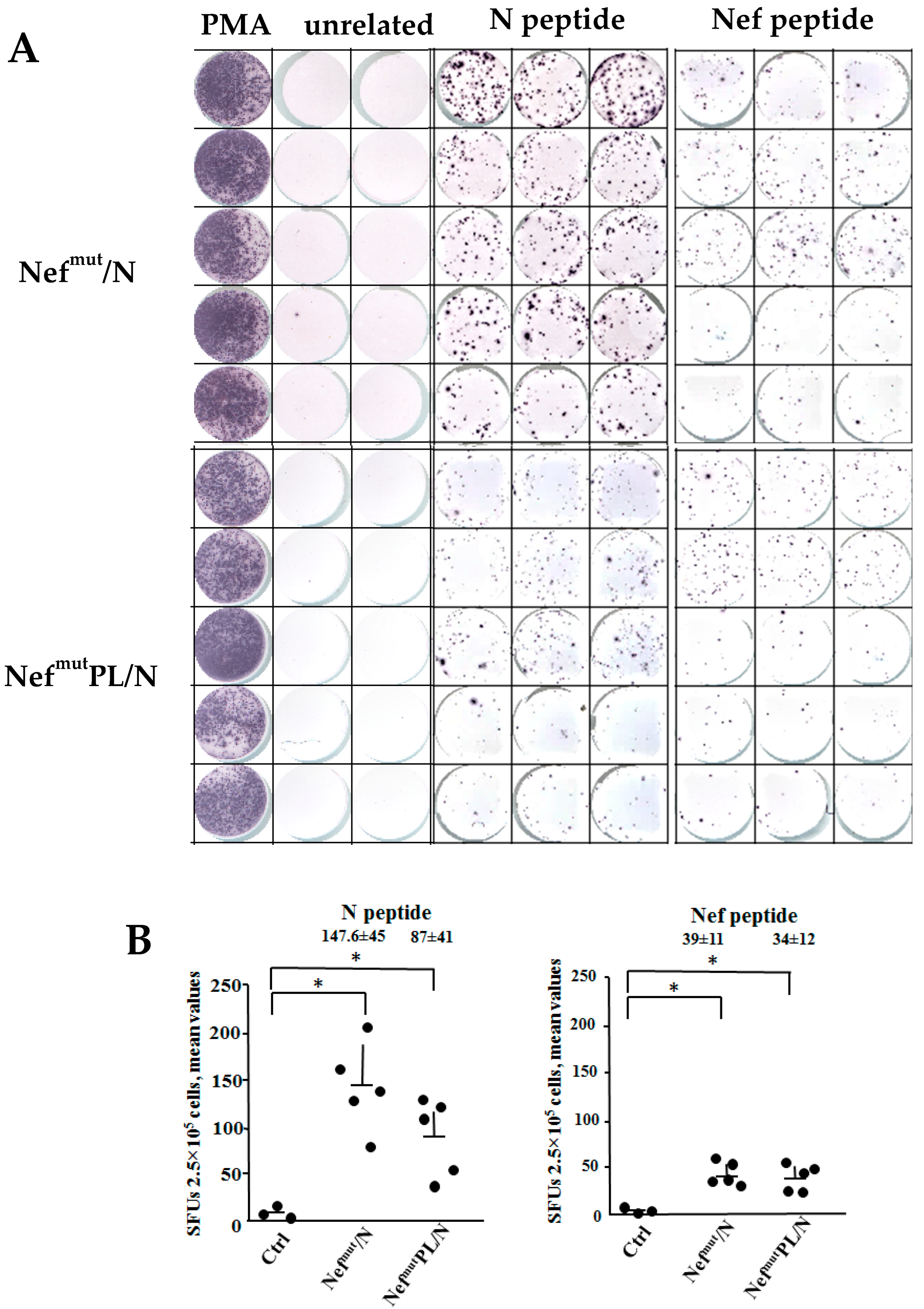
3.2. SARS-CoV-2 N-Specific Immunogenicity Induced in Mice Injected with DNA Vectors Expressing Either Nefmut/N or NefmutPL/N
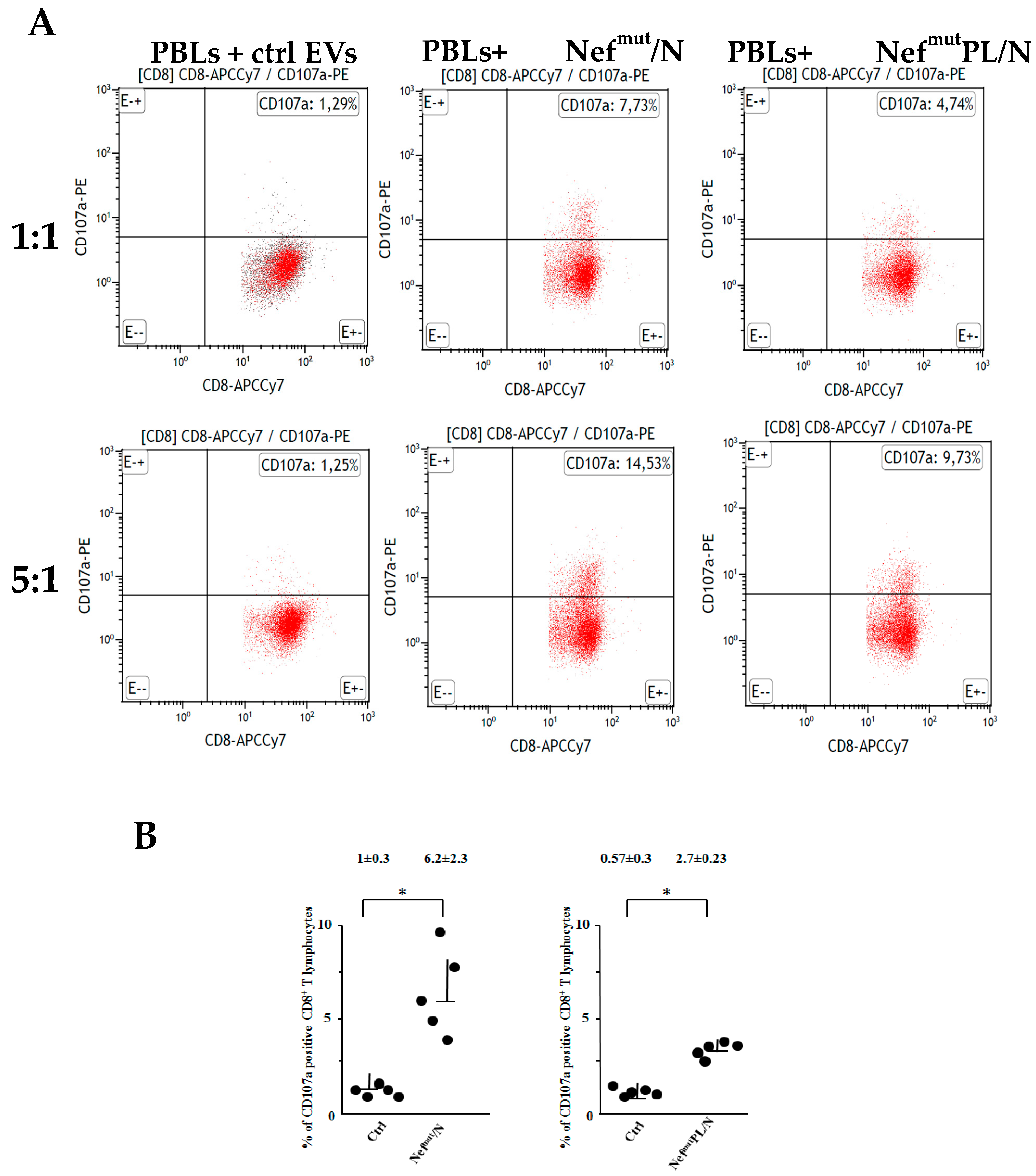
3.3. SARS-CoV-2 N Specific CTLs Are Generated upon Challenge of Human DCs with Engineered EVs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lozano-Rodríguez, R.; Valentín-Quiroga, J.; Avendaño-Ortiz, J.; Martín-Quirós, A.; Pascual-Iglesias, A.; Terrón-Arcos, V.; Montalbán-Hernández, K.; Casalvilla-Dueñas, J.C.; Bergón-Gutiérrez, M.; Alcamí, J.; et al. Cellular and Humoral Functional Responses after BNT162b2 MRNA Vaccination Differ Longitudinally between Naive and Subjects Recovered from COVID-19. Cell Rep. 2022, 38, 110235. [Google Scholar] [CrossRef] [PubMed]
- Goel, R.R.; Painter, M.M.; Apostolidis, S.A.; Mathew, D.; Meng, W.; Rosenfeld, A.M.; Lundgreen, K.A.; Reynaldi, A.; Khoury, D.S.; Pattekar, A.; et al. MRNA Vaccines Induce Durable Immune Memory to SARS-CoV-2 and Variants of Concern. Science 2021, 374, abm0829. [Google Scholar] [CrossRef] [PubMed]
- Piano Mortari, E.; Russo, C.; Vinci, M.R.; Terreri, S.; Fernandez Salinas, A.; Piccioni, L.; Alteri, C.; Colagrossi, L.; Coltella, L.; Ranno, S.; et al. Highly Specific Memory B Cells Generation after the 2nd Dose of BNT162b2 Vaccine Compensate for the Decline of Serum Antibodies and Absence of Mucosal IgA. Cells 2021, 10, 2541. [Google Scholar] [CrossRef]
- Haralambieva, I.H.; Monroe, J.M.; Ovsyannikova, I.G.; Grill, D.E.; Poland, G.A.; Kennedy, R.B. Homologous and Variant-Specific Memory B-Cell and Antibody Responses after SARS-CoV-2 mRNA Vaccination. medRxiv 2021. [Google Scholar] [CrossRef]
- Goel, R.R.; Apostolidis, S.A.; Painter, M.M.; Mathew, D.; Pattekar, A.; Kuthuru, O.; Gouma, S.; Hicks, P.; Meng, W.; Rosenfeld, A.M.; et al. Distinct Antibody and Memory B Cell Responses in SARS-CoV-2 Naïve and Recovered Individuals after MRNA Vaccination. Sci. Immunol. 2021, 6, eabi6950. [Google Scholar] [CrossRef] [PubMed]
- Sokal, A.; Chappert, P.; Barba-Spaeth, G.; Roeser, A.; Fourati, S.; Azzaoui, I.; Vandenberghe, A.; Fernandez, I.; Meola, A.; Bouvier-Alias, M.; et al. Maturation and Persistence of the Anti-SARS-CoV-2 Memory B Cell Response. Cell 2021, 184, 1201–1213.e14. [Google Scholar] [CrossRef]
- Mackay, L.K.; Rahimpour, A.; Ma, J.Z.; Collins, N.; Stock, A.T.; Hafon, M.-L.; Vega-Ramos, J.; Lauzurica, P.; Mueller, S.N.; Stefanovic, T.; et al. The Developmental Pathway for CD103+CD8+ Tissue-Resident Memory T Cells of Skin. Nat. Immunol. 2013, 14, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Schenkel, J.M.; Fraser, K.A.; Vezys, V.; Masopust, D. Sensing and Alarm Function of Resident Memory CD8+ T Cells. Nat. Immunol. 2013, 14, 509–513. [Google Scholar] [CrossRef] [Green Version]
- Allie, S.R.; Bradley, J.E.; Mudunuru, U.; Schultz, M.D.; Graf, B.A.; Lund, F.E.; Randall, T.D. The Establishment of Resident Memory B Cells in the Lung Requires Local Antigen Encounter. Nat. Immunol. 2019, 20, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Bleier, B.S.; Ramanathan, M.; Lane, A.P. COVID-19 Vaccines May Not Prevent Nasal SARS-CoV-2 Infection and Asymptomatic Transmission. Otolaryngol. Head Neck Surg. 2021, 164, 305–307. [Google Scholar] [CrossRef]
- Federico, M. Virus-Induced CD8+ T-Cell Immunity and Its Exploitation to Contain the SARS-CoV-2 Pandemic. Vaccines 2021, 9, 922. [Google Scholar] [CrossRef] [PubMed]
- Redd, A.D.; Nardin, A.; Kared, H.; Bloch, E.M.; Pekosz, A.; Laeyendecker, O.; Abel, B.; Fehlings, M.; Quinn, T.C.; Tobian, A.A.R. CD8+ T-Cell Responses in COVID-19 Convalescent Individuals Target Conserved Epitopes From Multiple Prominent SARS-CoV-2 Circulating Variants. Open Forum Infect. Dis. 2021, 8, ofab143. [Google Scholar] [CrossRef] [PubMed]
- Tarke, A.; Sidney, J.; Methot, N.; Yu, E.D.; Zhang, Y.; Dan, J.M.; Goodwin, B.; Rubiro, P.; Sutherland, A.; Wang, E.; et al. Impact of SARS-CoV-2 Variants on the Total CD4+ and CD8+ T Cell Reactivity in Infected or Vaccinated Individuals. Cell Rep. Med. 2021, 2, 100355. [Google Scholar] [CrossRef]
- Le Bert, N.; Tan, A.T.; Kunasegaran, K.; Tham, C.Y.L.; Hafezi, M.; Chia, A.; Chng, M.H.Y.; Lin, M.; Tan, N.; Linster, M.; et al. SARS-CoV-2-Specific T Cell Immunity in Cases of COVID-19 and SARS, and Uninfected Controls. Nature 2020, 584, 457–462. [Google Scholar] [CrossRef]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shdding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Di Bonito, P.; Chiozzini, C.; Arenaccio, C.; Anticoli, S.; Manfredi, F.; Olivetta, E.; Ferrantelli, F.; Falcone, E.; Ruggieri, A.; Federico, M. Antitumor HPV E7-Specific CTL Activity Elicited by in Vivo Engineered Exosomes Produced through DNA Inoculation. Int. J. Nanomed. 2017, 12, 4579–4591. [Google Scholar] [CrossRef] [Green Version]
- Anticoli, S.; Manfredi, F.; Chiozzini, C.; Arenaccio, C.; Olivetta, E.; Ferrantelli, F.; Capocefalo, A.; Falcone, E.; Ruggieri, A.; Federico, M. An Exosome-Based Vaccine Platform Imparts Cytotoxic T Lymphocyte Immunity Against Viral Antigens. Biotechnol. J. 2018, 13, e1700443. [Google Scholar] [CrossRef]
- Ferrantelli, F.; Chiozzini, C.; Manfredi, F.; Giovannelli, A.; Leone, P.; Federico, M. Simultaneous CD8 + T-Cell Immune Response against SARS-CoV-2 S, M, and N Induced by Endogenously Engineered Extracellular Vesicles in Both Spleen and Lungs. Vaccines 2021, 9, 240. [Google Scholar] [CrossRef]
- Ferrantelli, F.; Chiozzini, C.; Manfredi, F.; Leone, P.; Spada, M.; Di Virgilio, A.; Giovannelli, A.; Sanchez, M.; Cara, A.; Michelini, Z.; et al. Strong SARS-CoV-2 N-Specific CD8+ T Immunity Induced by Engineered Extracellular Vesicles Associates with Protection from Lethal Infection in Mice. Viruses 2022, 14, 329. [Google Scholar] [CrossRef]
- Chiozzini, C.; Manfredi, F.; Ferrantelli, F.; Leone, P.; Giovannelli, A.; Olivetta, E.; Federico, M. The C-Terminal Domain of Nefmut Is Dispensable for the CD8+ T Cell Immunogenicity of In Vivo Engineered Extracellular Vesicles. Vaccines 2021, 9, 373. [Google Scholar] [CrossRef]
- Anticoli, S.; Aricò, E.; Arenaccio, C.; Manfredi, F.; Chiozzini, C.; Olivetta, E.; Ferrantelli, F.; Lattanzi, L.; D’Urso, M.T.; Proietti, E.; et al. Engineered Exosomes Emerging from Muscle Cells Break Immune Tolerance to HER2 in Transgenic Mice and Induce Antigen-Specific CTLs upon Challenge by Human Dendritic Cells. J. Mol. Med. 2018, 96, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and Characterization of Exosomes from Cell Culture Supernatants and Biological Fluids. Curr. Protoc. Cell Biol. 2006, 30, 3.22.1–3.22.29. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhao, J.; Perlman, S. T Cell Responses Are Required for Protection from Clinical Disease and for Virus Clearance in Severe Acute Respiratory Syndrome Coronavirus-Infected Mice. J. Virol. 2010, 84, 9318–9325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Fu, T.-M.; Xie, H.; Emini, E.A.; Shiver, J.W. Development of HIV-1 Nef Vaccine Components: Immunogenicity Study of Nef Mutants Lacking Myristoylation and Dileucine Motif in Mice. Vaccine 2002, 20, 3413–3421. [Google Scholar] [CrossRef]
- Betts, M.R.; Koup, R.A. Detection of T-Cell Degranulation: CD107a and b. Methods Cell Biol. 2004, 75, 497–512. [Google Scholar] [CrossRef]
- Ahmed, S.F.; Quadeer, A.A.; McKay, M.R. Preliminary Identification of Potential Vaccine Targets for the COVID-19 Coronavirus (SARS-CoV-2) Based on SARS-CoV Immunological Studies. Viruses 2020, 12, 254. [Google Scholar] [CrossRef] [Green Version]
- Hudrisier, D.; Riond, J.; Mazarguil, H.; Gairin, J.E.; Joly, E. Cutting Edge: CTLs Rapidly Capture Membrane Fragments from Target Cells in a TCR Signaling-Dependent Manner. J. Immunol. 2001, 166, 3645–3649. [Google Scholar] [CrossRef] [Green Version]
- Stinchcombe, J.C.; Bossi, G.; Booth, S.; Griffiths, G.M. The Immunological Synapse of CTL Contains a Secretory Domain and Membrane Bridges. Immunity 2001, 15, 751–761. [Google Scholar] [CrossRef] [Green Version]
- Joly, E.; Hudrisier, D. What Is Trogocytosis and What Is Its Purpose? Nat. Immunol. 2003, 4, 815. [Google Scholar] [CrossRef]
- Beadling, C.; Slifka, M.K. Quantifying Viable Virus-Specific T Cells without a Priori Knowledge of Fine Epitope Specificity. Nat. Med. 2006, 12, 1208–1212. [Google Scholar] [CrossRef]
- Tomaru, U.; Yamano, Y.; Nagai, M.; Maric, D.; Kaumaya, P.T.P.; Biddison, W.; Jacobson, S. Detection of Virus-Specific T Cells and CD8+ T-Cell Epitopes by Acquisition of Peptide-HLA-GFP Complexes: Analysis of T-Cell Phenotype and Function in Chronic Viral Infections. Nat. Med. 2003, 9, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Machlenkin, A.; Uzana, R.; Frankenburg, S.; Eisenberg, G.; Eisenbach, L.; Pitcovski, J.; Gorodetsky, R.; Nissan, A.; Peretz, T.; Lotem, M. Capture of Tumor Cell Membranes by Trogocytosis Facilitates Detection and Isolation of Tumor-Specific Functional CTLs. Cancer Res. 2008, 68, 2006–2013. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.-H.; Plemenitas, A.; Fielding, C.J.; Peterlin, B.M. Nef Increases the Synthesis of and Transports Cholesterol to Lipid Rafts and HIV-1 Progeny Virions. Proc. Natl. Acad. Sci. USA 2003, 100, 8460–8465. [Google Scholar] [CrossRef] [Green Version]
- Cai, C.Y.; Zhang, X.; Sinko, P.J.; Burakoff, S.J.; Jin, Y.-J. Two Sorting Motifs, a Ubiquitination Motif and a Tyrosine Motif, Are Involved in HIV-1 and Simian Immunodeficiency Virus Nef-Mediated Receptor Endocytosis. J. Immunol. 2011, 186, 5807–5814. [Google Scholar] [CrossRef] [Green Version]
- McNamara, R.P.; Caro-Vegas, C.P.; Costantini, L.M.; Landis, J.T.; Griffith, J.D.; Damania, B.A.; Dittmer, D.P. Large-Scale, Cross-Flow Based Isolation of Highly Pure and Endocytosis-Competent Extracellular Vesicles. J. Extracell. Vesicles 2018, 7, 1541396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paganini, C.; Capasso Palmiero, U.; Pocsfalvi, G.; Touzet, N.; Bongiovanni, A.; Arosio, P. Scalable Production and Isolation of Extracellular Vesicles: Available Sources and Lessons from Current Industrial Bioprocesses. Biotechnol. J. 2019, 14, e1800528. [Google Scholar] [CrossRef] [PubMed]
- Murugappan, S.; Frijlink, H.W.; Petrovsky, N.; Hinrichs, W.L.J. Enhanced Pulmonary Immunization with Aerosolized Inactivated Influenza Vaccine Containing Delta Inulin Adjuvant. Eur. J. Pharm. Sci. 2015, 66, 118–122. [Google Scholar] [CrossRef]
- Xu, J.; Li, S.; Wang, X.; Liu, J.; Shan, P.; Zhou, Y.; Zhao, J.; Wang, Z.; Xu, C.; Chen, M.; et al. Systemic and Mucosal Humoral Immune Responses Induced by the JY-Adjuvanted Nasal Spray H7N9 Vaccine in Mice. Emerg. Microbes Infect. 2018, 7, 140. [Google Scholar] [CrossRef] [Green Version]
- Sistigu, A.; Bracci, L.; Valentini, M.; Proietti, E.; Bona, R.; Negri, D.R.M.; Ciccaglione, A.R.; Tritarelli, E.; Nisini, R.; Equestre, M.; et al. Strong CD8+ T Cell Antigenicity and Immunogenicity of Large Foreign Proteins Incorporated in HIV-1 VLPs Able to Induce a Nef-Dependent Activation/Maturation of Dendritic Cells. Vaccine 2011, 29, 3465–3475. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manfredi, F.; Chiozzini, C.; Ferrantelli, F.; Leone, P.; Giovannelli, A.; Sanchez, M.; Federico, M. Activation of Anti-SARS-CoV-2 Human CTLs by Extracellular Vesicles Engineered with the N Viral Protein. Vaccines 2022, 10, 1060. https://doi.org/10.3390/vaccines10071060
Manfredi F, Chiozzini C, Ferrantelli F, Leone P, Giovannelli A, Sanchez M, Federico M. Activation of Anti-SARS-CoV-2 Human CTLs by Extracellular Vesicles Engineered with the N Viral Protein. Vaccines. 2022; 10(7):1060. https://doi.org/10.3390/vaccines10071060
Chicago/Turabian StyleManfredi, Francesco, Chiara Chiozzini, Flavia Ferrantelli, Patrizia Leone, Andrea Giovannelli, Massimo Sanchez, and Maurizio Federico. 2022. "Activation of Anti-SARS-CoV-2 Human CTLs by Extracellular Vesicles Engineered with the N Viral Protein" Vaccines 10, no. 7: 1060. https://doi.org/10.3390/vaccines10071060