
Adenoviral-Vectored Centralized Consensus Hemagglutinin Vaccine Provides Broad Protection against H2 Influenza a Virus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Influenza Viruses
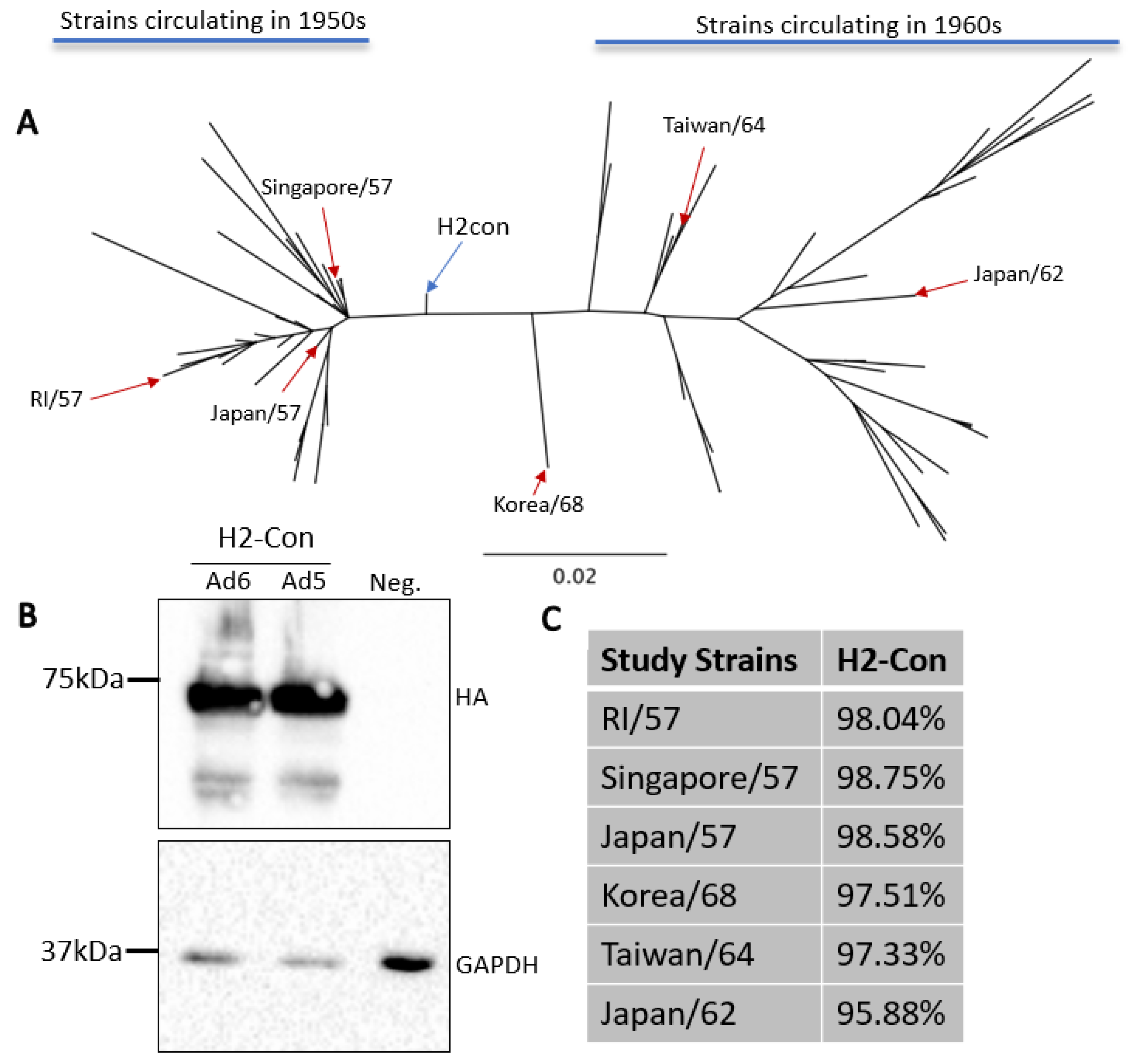
2.3. Design of Centralized Consensus Hemagglutinin and Phylogenetic Analysis
2.4. Construction of Recombinant Adenoviruses
2.5. Western Blot
2.6. Vaccination
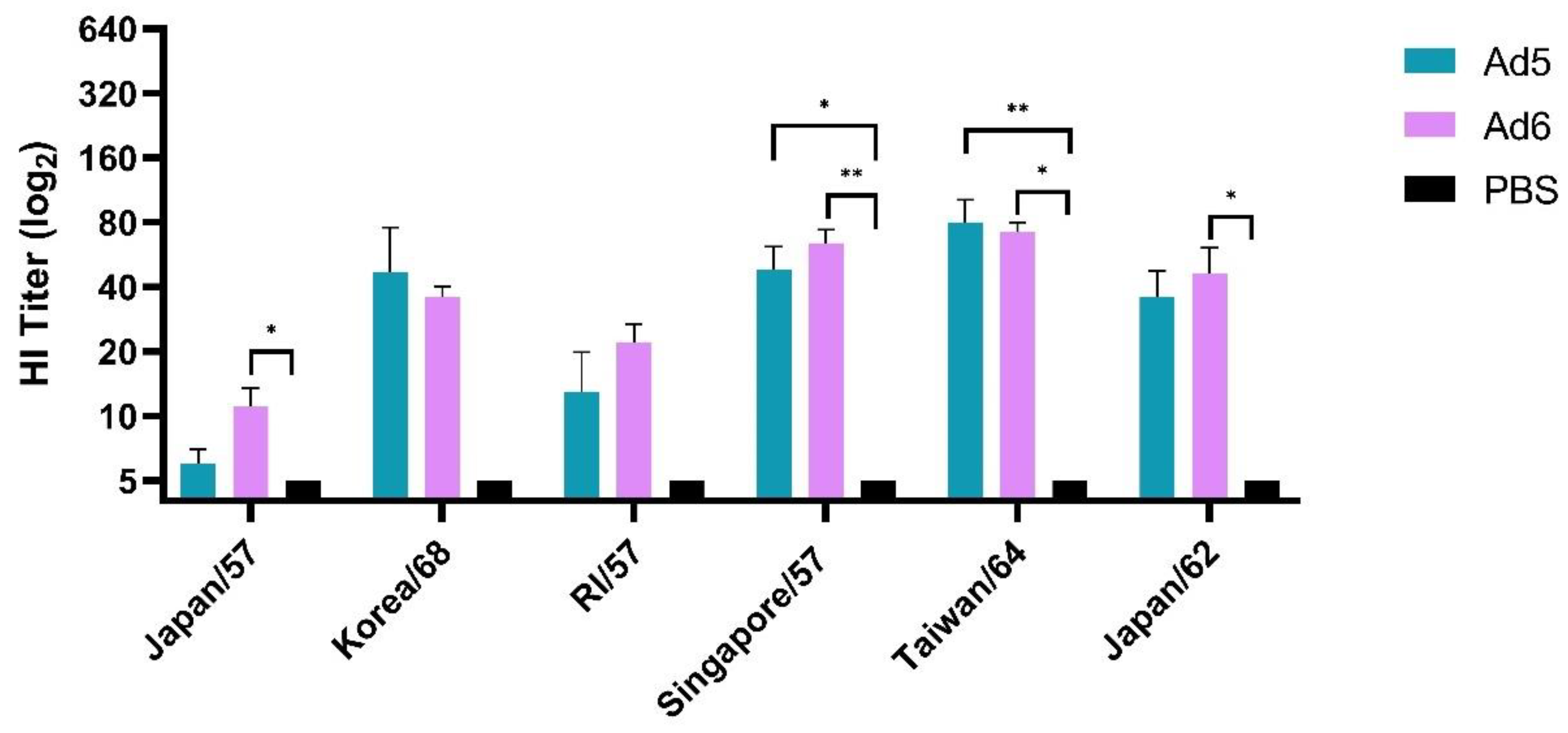
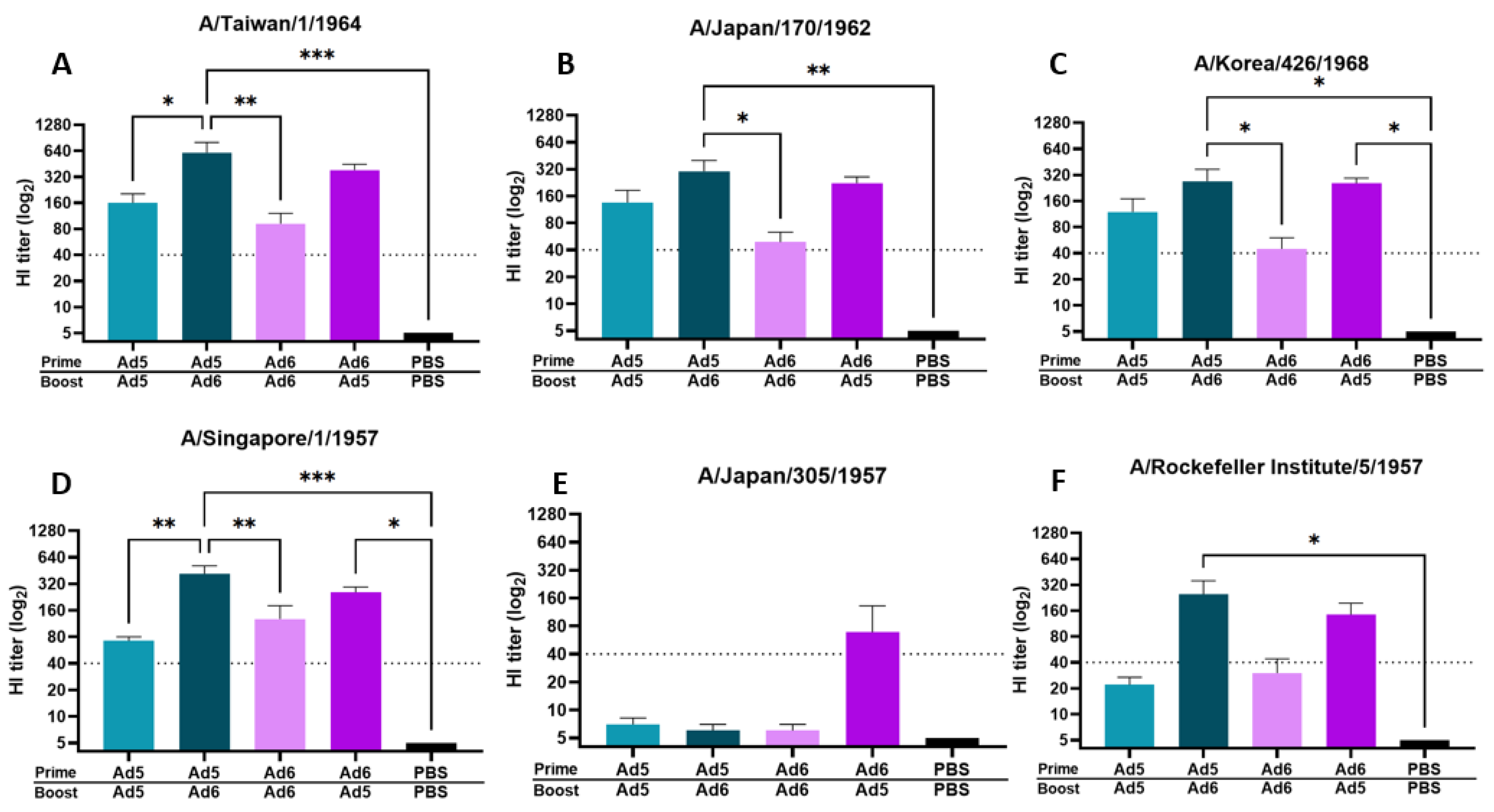
2.7. Hemagglutination Inhibition (HI) Assay
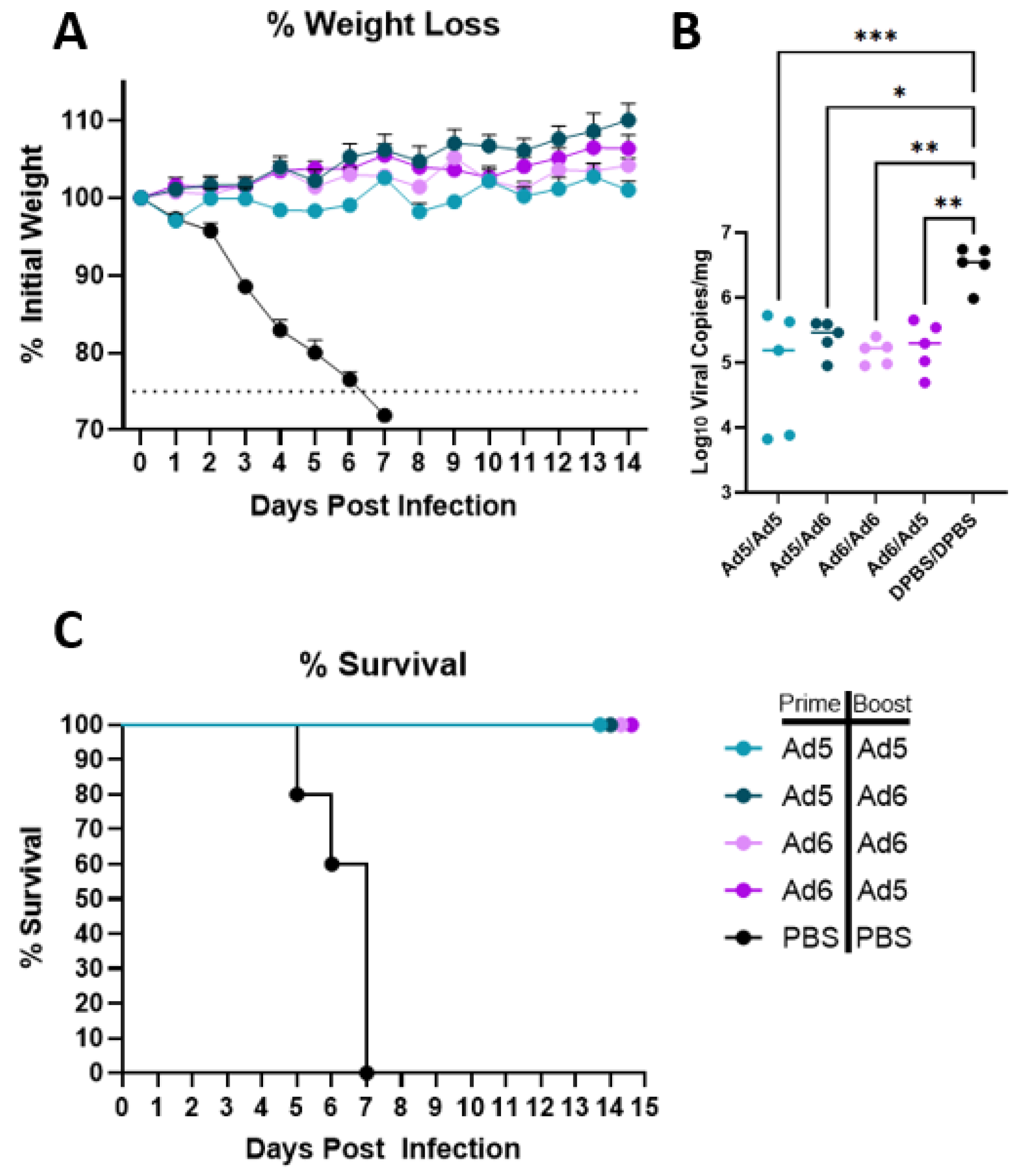
2.8. Lethal Influenza Challenge, Tissue Collection, and Lung Viral Titers
2.9. Statistical Analysis
3. Results
3.1. Construction and Characterization of Centralized Consensus H2 Vaccine Immunogen
3.2. Adenoviral-Vectored Consensus H2 Immunogen Induces Protective Humoral Immune Responses
3.3. H2 Consensus Provides Protection against Lethal A/Korea/426/1968 Challenge
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simonsen, L.; Clarke, M.J.; Schonberger, L.B.; Arden, N.H.; Cox, N.J.; Fukuda, K. Pandemic versus Epidemic Influenza Mortality: A Pattern of Changing Age Distribution. J. Infect. Dis. 1998, 178, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Lindstrom, S.E.; Cox, N.J.; Klimov, A. Genetic analysis of human H2N2 and early H3N2 influenza viruses, 1957–1972: Evidence for genetic divergence and multiple reassortment events. Virology 2004, 328, 101–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babu, T.M.; Perera, R.A.; Wu, J.T.K.; Fitzgerald, T.; Nolan, C.; Cowling, B.; Krauss, S.; Treanor, J.J.; Peiris, M. Population Serologic Immunity to Human and Avian H2N2 Viruses in the United States and Hong Kong for Pandemic Risk Assessment. J. Infect. Dis. 2018, 218, 1054–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reneer, Z.B.; Abreu, R.B.; Jamal, U.S.; Corn, M.R.; Paugh, J.L.; Ross, T.M. Seasonal influenza vaccination does not effectively expand H2 cross-reactive antibodies in humans. Vaccine 2021, 39, 4173–4183. [Google Scholar] [CrossRef]
- Munster, V.J.; Baas, C.; Lexmond, P.; Waldenstrom, J.; Wallensten, A.; Fransson, T.; Rimmelzwaan, G.F.; Beyer, W.E.; Schutten, M.; Olsen, B.; et al. Spatial, temporal, and species variation in prevalence of influenza A viruses in wild migratory birds. PLoS Pathog. 2007, 3, e61. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Peng, X.; Peng, X.; Cheng, L.; Wu, N. Genetic and molecular characterization of a novel reassortant H2N8 subtype avian influenza virus isolated from a domestic duck in Zhejiang Province in China. Virus Genes 2016, 52, 863–866. [Google Scholar] [CrossRef]
- Ma, M.-J.; Yang, X.-X.; Qian, Y.-H.; Zhao, S.-Y.; Hua, S.; Wang, T.-C.; Chen, S.-H.; Ma, G.-Y.; Sang, X.-Y.; Liu, L.-N.; et al. Characterization of a Novel Reassortant Influenza A Virus (H2N2) from a Domestic Duck in Eastern China. Sci. Rep. 2014, 4, 7588. [Google Scholar] [CrossRef]
- Lee, D.-H.; Park, K.-D. The preparedness plan for influenza pandemic. J. Prev. Med. Public Health 2005, 38, 386–390. [Google Scholar]
- Kieny, M.P.; Costa, A.; Hombach, J.; Carrasco, P.; Pervikov, Y.; Salisbury, D.; Greco, M.; Gust, I.; LaForce, M.; Franco-Paredes, C.; et al. A global pandemic influenza vaccine action plan. Vaccine 2006, 24, 6367–6370. [Google Scholar] [CrossRef]
- Linster, M.; Schrauwen, E.J.A.; van der Vliet, S.; Burke, D.; Lexmond, P.; Bestebroer, T.M.; Smith, D.; Herfst, S.; Koel, B.F.; Fouchier, R.A.M. The Molecular Basis for Antigenic Drift of Human A/H2N2 Influenza Viruses. J. Virol. 2019, 93, e01907-18. [Google Scholar] [CrossRef] [Green Version]
- Lenny, B.J.; Sonnberg, S.; Danner, A.F.; Friedman, K.; Webby, R.J.; Webster, R.G.; Jones, J.C. Evaluation of multivalent H2 influenza pandemic vaccines in mice. Vaccine 2017, 35, 1455–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.L.; Lamirande, E.W.; Cheng, X.; Torres-Velez, F.; Orandle, M.; Jin, H.; Kemble, G.; Subbarao, K. Evaluation of Three Live Attenuated H2 Pandemic Influenza Vaccine Candidates in Mice and Ferrets. J. Virol. 2014, 88, 2867–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isakova-Sivak, I.; Stukova, M.; Erofeeva, M.; Naykhin, A.; Donina, S.; Petukhova, G.; Kuznetsova, V.; Kiseleva, I.; Smolonogina, T.; Dubrovina, I.; et al. H2N2 live attenuated influenza vaccine is safe and immunogenic for healthy adult volunteers. Hum. Vaccines Immunother. 2015, 11, 970–982. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.L.; Lamirande, E.W.; Jin, H.; Kemble, G.; Subbarao, K. Safety, immunogencity, and efficacy of a cold-adapted A/Ann Arbor/6/60 (H2N2) vaccine in mice and ferrets. Virology 2010, 398, 109–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talaat, K.R.; Karron, R.A.; Liang, P.H.; McMahon, B.A.; Luke, C.J.; Thumar, B.; Chen, G.L.; Min, J.-Y.; Lamirande, E.W.; Jin, H.; et al. An open-label phase I trial of a live attenuated H2N2 influenza virus vaccine in healthy adults. Influ. Other Respir. Viruses 2012, 7, 66–73. [Google Scholar] [CrossRef]
- Weaver, E.A.; Rubrum, A.M.; Webby, R.J.; Barry, M.A. Protection against Divergent Influenza H1N1 Virus by a Centralized Influenza Hemagglutinin. PLoS ONE 2011, 6, e18314. [Google Scholar] [CrossRef]
- Lingel, A.; Bullard, B.; Weaver, E.A. Efficacy of an Adenoviral Vectored Multivalent Centralized Influenza Vaccine. Sci. Rep. 2017, 7, 14912. [Google Scholar] [CrossRef]
- Nuñez, I.A.; Huang, Y.; Ross, T.M. Next-Generation Computationally Designed Influenza Hemagglutinin Vaccines Protect against H5Nx Virus Infections. Pathogens 2021, 10, 1352. [Google Scholar] [CrossRef]
- Giles, B.M.; Bissel, S.J.; DeAlmeida, D.R.; Wiley, C.A.; Ross, T.M. Antibody Breadth and Protective Efficacy Are Increased by Vaccination with Computationally Optimized Hemagglutinin but Not with Polyvalent Hemagglutinin-Based H5N1 Virus-Like Particle Vaccines. Clin. Vaccine Immunol. 2012, 19, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Carter, D.M.; Darby, C.A.; Lefoley, B.C.; Crevar, C.J.; Alefantis, T.; Oomen, R.; Anderson, S.F.; Strugnell, T.; Cortés-Garcia, G.; Vogel, T.U.; et al. Design and Characterization of a Computationally Optimized Broadly Reactive Hemagglutinin Vaccine for H1N1 Influenza Viruses. J. Virol. 2016, 90, 4720–4734. [Google Scholar] [CrossRef] [Green Version]
- Mohn, K.G.-I.; Smith, I.; Sjursen, H.; Cox, R.J. Immune responses after live attenuated influenza vaccination. Hum. Vaccines Immunother. 2017, 14, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Minor, P. Vaccine-derived poliovirus (VDPV): Impact on poliomyelitis eradication. Vaccine 2009, 27, 2649–2652. [Google Scholar] [CrossRef] [PubMed]
- Luke, C.J.; Subbarao, K. Vaccines for pandemic influenza. Emerg. Infect. Dis. 2006, 12, 66–72. [Google Scholar] [CrossRef] [PubMed]
- WHO. Live attenuated vaccines (LAV). In Vaccine Safety Basics: E-learning Course; WHO: Geneva, Switzerland, 2013. [Google Scholar]
- Sharma, A.; Zeller, M.; Li, G.; Harmon, K.M.; Zhang, J.; Hoang, H.; Anderson, T.K.; Vincent, A.L.; Gauger, P.C. Detection of live attenuated influenza vaccine virus and evidence of reassortment in the U.S. swine population. J. Vet. Diagn. Investig. 2020, 32, 301–311. [Google Scholar] [CrossRef]
- Maassab, H.F. Adaptation and growth characteristics of influenza virus at 25 degrees c. Nature 1967, 213, 612–614. [Google Scholar] [CrossRef]
- Maassab, H.F. Biologic and immunologic characteristics of cold-adapted influenza virus. J. Immunol. 1969, 102, 728–732. [Google Scholar]
- Coughlan, L. Factors Which Contribute to the Immunogenicity of Non-replicating Adenoviral Vectored Vaccines. Front. Immunol. 2020, 11, 909. [Google Scholar] [CrossRef]
- Gene Therapy Clinical Trials (Wiley). Available online: https://a873679.fmphost.com/fmi/webd/GTCT (accessed on 11 April 2022).
- Vemula, S.V.; Mittal, S.K. Production of adenovirus vectors and their use as a delivery system for influenza vaccines. Expert Opin. Biol. Ther. 2010, 10, 1469–1487. [Google Scholar] [CrossRef]
- Zhu, F.-C.; Guan, X.-H.; Li, Y.-H.; Huang, J.-Y.; Jiang, T.; Hou, L.-H.; Li, J.-X.; Yang, B.-F.; Wang, L.; Wang, W.-J.; et al. Immunogenicity and safety of a recombinant adenovirus type-5-vectored COVID-19 vaccine in healthy adults aged 18 years or older: A randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2020, 396, 479–488. [Google Scholar] [CrossRef]
- Falsey, A.R.; Sobieszczyk, M.E.; Hirsch, I.; Sproule, S.; Robb, M.L.; Corey, L.; Neuzil, K.M.; Hahn, W.; Hunt, J.; Mulligan, M.J.; et al. Phase 3 Safety and Efficacy of AZD1222 (ChAdOx1 nCoV-19) COVID-19 Vaccine. N. Engl. J. Med. 2021, 385, 2348–2360. [Google Scholar] [CrossRef]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Sadoff, J.; Le Gars, M.; Shukarev, G.; Heerwegh, D.; Truyers, C.; de Groot, A.M.; Stoop, J.; Tete, S.; Van Damme, W.; Leroux-Roels, I.; et al. Interim Results of a Phase 1-2a Trial of Ad26.COV2.S COVID-19 Vaccine. N. Engl. J. Med. 2021, 384, 1824–1835. [Google Scholar] [CrossRef] [PubMed]
- Logunov, D.Y.; Dolzhikova, I.V.; Shcheblyakov, D.V.; Tukhvatulin, A.I.; Zubkova, O.V.; Dzharullaeva, A.S.; Kovyrshina, A.V.; Lubenets, N.L.; Grousova, D.M.; Erokhova, A.S.; et al. Safety and efficacy of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine: An interim analysis of a randomised controlled phase 3 trial in Russia. Lancet 2021, 397, 671–681. [Google Scholar] [CrossRef]
- Tsao, Y.S.; Condon, R.; Schaefer, E.; Lio, P.; Liu, Z. Development and improvement of a serum-free suspension process for the production of recombinant ade-noviral vectors using HEK293 cells. Cytotechnology 2001, 37, 189–198. [Google Scholar] [CrossRef]
- Weaver, E.A.; Lu, Z.; Camacho, Z.T.; Moukdar, F.; Liao, H.-X.; Ma, B.-J.; Muldoon, M.; Theiler, J.; Nabel, G.J.; Letvin, N.L.; et al. Cross-Subtype T-Cell Immune Responses Induced by a Human Immunodeficiency Virus Type 1 Group M Consensus Env Immunogen. J. Virol. 2006, 80, 6745–6756. [Google Scholar] [CrossRef] [Green Version]
- Liao, H.-X.; Sutherland, L.L.; Xia, S.-M.; Brock, M.E.; Scearce, R.M.; Vanleeuwen, S.; Alam, S.M.; McAdams, M.; Weaver, E.A.; Camacho, Z.T.; et al. A group M consensus envelope glycoprotein induces antibodies that neutralize subsets of subtype B and C HIV-1 primary viruses. Virology 2006, 353, 268–282. [Google Scholar] [CrossRef] [Green Version]
- Kothe, D.L.; Li, Y.; Decker, J.M.; Bibollet-Ruche, F.; Zammit, K.P.; Salazar, M.G.; Chen, Y.; Weng, Z.; Weaver, E.A.; Gao, F.; et al. Ancestral and consensus envelope immunogens for HIV-1 subtype C. Virology 2006, 352, 438–449. [Google Scholar] [CrossRef] [Green Version]
- Weaver, E.A.; Nehete, P.; Buchl, S.S.; Senac, J.S.; Palmer, D.; Ng, P.; Sastry, J.; Barry, M.A. Comparison of Replication-Competent, First Generation, and Helper-Dependent Adenoviral Vaccines. PLoS ONE 2009, 4, e5059. [Google Scholar] [CrossRef]
- Ohmit, S.E.; Petrie, J.G.; Cross, R.T.; Johnson, E.; Monto, A.S. Influenza Hemagglutination-Inhibition Antibody Titer as a Correlate of Vaccine-Induced Protection. J. Infect. Dis. 2011, 204, 1879–1885. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Stevens, D.J.; Haire, L.F.; Walker, P.A.; Coombs, P.J.; Russell, R.J.; Gamblin, S.J.; Skehel, J.J. Structures of receptor complexes formed by hemagglutinins from the Asian Influenza pandemic of 1957. Proc. Natl. Acad. Sci. USA 2009, 106, 17175–17180. [Google Scholar] [CrossRef] [Green Version]
- Reneer, Z.B.; Ross, T.M. H2 influenza viruses: Designing vaccines against future H2 pandemics. Biochem. Soc. Trans. 2019, 47, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, E.; Sugawara, K.; Hongo, S.; Matsuzaki, Y.; Muraki, Y.; Li, Z.-N.; Nakamura, K. Antigenic structure of the haemagglutinin of human influenza A/H2N2 virus. J. Gen. Virol. 2001, 82 Pt 10, 2475–2484. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.; Sealy, J.E.; Sadeyen, J.-R.; Bhat, S.; Lukosaityte, D.; Sun, Y.; Iqbal, M. Immune Escape Adaptive Mutations in the H7N9 Avian Influenza Hemagglutinin Protein Increase Virus Replication Fitness and Decrease Pandemic Potential. J. Virol. 2020, 94, e00216-20. [Google Scholar] [CrossRef] [PubMed]
- Sitaras, I.; Kalthoff, D.; Beer, M.; Peeters, B.; de Jong, M.C. Immune escape mutants of Highly Pathogenic Avian Influenza H5N1 selected using polyclonal sera: Identi-fication of key amino acids in the HA protein. PLoS ONE 2014, 9, e84628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igarashi, M.; Ito, K.; Kida, H.; Takada, A. Genetically destined potentials for N-linked glycosylation of influenza virus hemagglutinin. Virology 2008, 376, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shashkova, E.V.; May, S.M.; Barry, M.A. Characterization of human adenovirus serotypes 5, 6, 11, and 35 as anticancer agents. Virology 2009, 394, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Weaver, E.A.; Hillestad, M.L.; Khare, R.; Palmer, D.; Ng, P.; Barry, M.A. Characterization of species C human adenovirus serotype 6 (Ad6). Virology 2011, 412, 19–27. [Google Scholar] [CrossRef]
- Adhikary, A.K.; Banik, U.; Numaga, J.; Suzuki, E.; Inada, T.; Okabe, N. Heterogeneity of the fibre sequence in subgenus C adenoviruses. J. Clin. Pathol. 2004, 57, 612–617. [Google Scholar] [CrossRef] [Green Version]
- Wu, E.; Pache, L.; Von Seggern, D.J.; Mullen, T.-M.; Mikyas, Y.; Stewart, P.L.; Nemerow, G.R. Flexibility of the Adenovirus Fiber Is Required for Efficient Receptor Interaction. J. Virol. 2003, 77, 7225–7235. [Google Scholar] [CrossRef] [Green Version]
- Mennechet, F.J.D.; Paris, O.; Ouoba, A.R.; Arenas, S.S.; Sirima, S.B.; Dzomo, G.R.T.; Diarra, A.; Traore, I.T.; Kania, D.; Eichholz, K.; et al. A review of 65 years of human adenovirus seroprevalence. Expert Rev. Vaccines 2019, 18, 597–613. [Google Scholar] [CrossRef]
- Capone, S.; Meola, A.; Ercole, B.B.; Vitelli, A.; Pezzanera, M.; Ruggeri, L.; Davies, M.E.; Tafi, R.; Santini, C.; Luzzago, A.; et al. A Novel Adenovirus Type 6 (Ad6)-Based Hepatitis C Virus Vector That Overcomes Preexisting Anti-Ad5 Immunity and Induces Potent and Broad Cellular Immune Responses in Rhesus Macaques. J. Virol. 2006, 80, 1688–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tollefson, A.E.; Ying, B.; Spencer, J.F.; Sagartz, J.E.; Wold, W.S.; Toth, K. Pathology in Permissive Syrian Hamsters after Infection with Species C Human Adenovirus (HAdV-C) Is the Result of Virus Replication: HAdV-C6 Replicates More and Causes More Pathology than HAdV-C5. J. Virol. 2017, 91, e00284-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khare, R.; May, S.M.; Vetrini, F.; Weaver, E.A.; Palmer, D.; Rosewell, A.; Grove, N.; Ng, P.; Barry, M.A. Generation of a Kupffer Cell-evading Adenovirus for Systemic and Liver-directed Gene Transfer. Mol. Ther. 2011, 19, 1254–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, J.; Ward, S.; Mendy, J.; Manayani, D.J.; Farness, P.; Avanzini, J.B.; Guenther, B.; Garduno, F.; Jow, L.; Snarsky, V.; et al. Pre-Clinical Evaluation of a Replication-Competent Recombinant Adenovirus Serotype 4 Vaccine Expressing Influenza H5 Hemagglutinin. PLoS ONE 2012, 7, e31177. [Google Scholar] [CrossRef]
- Matsuda, K.; Migueles, S.A.; Huang, J.; Bolkhovitinov, L.; Stuccio, S.; Griesman, T.; Pullano, A.A.; Kang, B.H.; Ishida, E.; Zimmerman, M.; et al. A replication-competent adenovirus-vectored influenza vaccine induces durable systemic and mucosal immunity. J. Clin. Investig. 2021, 131, e140794. [Google Scholar] [CrossRef]
- Ma, W.; Lager, K.M.; Vincent, A.L.; Janke, B.H.; Gramer, M.R.; Richt, J.A. The role of swine in the generation of novel influenza viruses. Zoonoses Public Health 2009, 56, 326–337. [Google Scholar] [CrossRef]
- Ma, W.; E Kahn, R.; Richt, J.A. The pig as a mixing vessel for influenza viruses: Human and veterinary implications. J. Mol. Genet. Med. 2009, 3, 158–166. [Google Scholar] [CrossRef]
- Ma, W.; Vincent, A.L.; Gramer, M.R.; Brockwell, C.B.; Lager, K.M.; Janke, B.H.; Gauger, P.C.; Patnayak, D.P.; Webby, R.J.; Richt, J.A. Identification of H2N3 influenza A viruses from swine in the United States. Proc. Natl. Acad. Sci. USA 2007, 104, 20949–20954. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petro-Turnquist, E.M.; Bullard, B.L.; Pekarek, M.J.; Weaver, E.A. Adenoviral-Vectored Centralized Consensus Hemagglutinin Vaccine Provides Broad Protection against H2 Influenza a Virus. Vaccines 2022, 10, 926. https://doi.org/10.3390/vaccines10060926
Petro-Turnquist EM, Bullard BL, Pekarek MJ, Weaver EA. Adenoviral-Vectored Centralized Consensus Hemagglutinin Vaccine Provides Broad Protection against H2 Influenza a Virus. Vaccines. 2022; 10(6):926. https://doi.org/10.3390/vaccines10060926
Chicago/Turabian StylePetro-Turnquist, Erika M., Brianna L. Bullard, Matthew J. Pekarek, and Eric A. Weaver. 2022. "Adenoviral-Vectored Centralized Consensus Hemagglutinin Vaccine Provides Broad Protection against H2 Influenza a Virus" Vaccines 10, no. 6: 926. https://doi.org/10.3390/vaccines10060926